

системные механизмы
.pdfотносительно простые аналитические описания основных51 свойств системы. Напротив, исследования в ненейтральной динамике, связывающие конкурентоспособные стратегии и существенные свойства экосистемы. в недавнем прошлом были ограничены, вследствие необходимости проведения исследования гетерогенных демографических норм поведения и относительным недостатком детальных наблюдений пространственного распределения разнообразия разновидностей (beta-разнообразие).
Предложена ненейтральная (Marani M, Zillio T, Belluco E, Silvestri S, Maritan A. 2006) модель динамики растительности, основанная на клонировании и числах дисперсии, которая формирует характеристики beta-разнообразия. Предсказания ненейтральной модели подтверждены, на материале наблюдений растительности солончака в Венецианской Лагуне (Италия) с высокой степенью точности. Модельные выражения beta-разнообразия показывают существенное совпадение с наблюдаемыми распределениями. Рассчитывается также нейтральная версия модели. Авторы заключают, что ненейтральные характеристики beta-разнообразия способны описывать динамику экосистемы в присутствии зависимых разновидностями свойств и успешно связывать наблюдаемые паттерны с процессами формирующими растительность.
Проблема анализа сложных систем, требующая, зачастую, применения методов нелинейного оценивания, делает необходимым разработку более простых алгоритмов.
Предпринята попытка описания сложных форм поведении я внутриклеточных сетей обычными дифференциальными уравнениями с известными кинетическими параметрами. При анализе предлагается расчленять полную систему на множество менее сложно организованных подсистем, а затем рассматривать возможность взаимодействий между ними. При данном алгоритма сеть анализируется как совокупность непостоянных устойчивых состояний, формирующих сложное нелинейное поведение. Также анализируются механизмы, дестабилизации этих устойчивых состояний. Обсуждаются проблемы пределов данного подхода. Предложенный метод был применен при моделировании митотического управления в Xenopus яйцах лягушки, и к модели циркадных колебаний в Дрозофиле (Schmidt H., Jacobsen E.W. 2004).
В настоящее время сформировались направления системного анализа называемые «биоинформатика» и «нейроинформатика».
Биоинформатика использует системную идеологию и52 компьютерные методы для решения задач молекулярной биологии, в основном анализа разных последовательностей (аминокислотных, нуклеотидных). Эта наука возникла в 1976-1978 годах, окончательно оформилась в 1980 году в связи со специальным выпуском журнала
«Nucleic Acid Research» (NAR). Биоинформатика включает в себя:
базы данных, в которых хранится биологическая информация
набор инструментов для анализа тех данных, которые лежат в таких базах
правильное применение компьютерных методов для правильного решения биологических задач
Биолог в биоинформатике обычно имеет дело с базами данных и инструментами их анализа.
Ние приведены примеры некоторых достижений в биоинформатике на основе «Отчет о деятельности Российской академии наук в 2002 году. Основные результаты в области естественных, технических, гуманитарных и общественных наук. Москва 2003».
Разработана модель оптической ассоциативной памяти на основе известного в нелинейной оптике процесса параметрического четырехволнового смешения. Элементная база сети - параметрические нейроны с q дискретными состояниями; с ростом q
увеличивается помехозащищенность сети и объем нейросетевой памяти (в q2 раз большая, чем у стандартных нейросетевых архитектур). (ИОНТ РАН).
Разработана по методу autonomous adaptive control (AAC)
бионическая нейроноподобная система управления, построенная на базе специальных нейроноподобных элементов, исследовано ее применение для адаптивного управления мобильным роботом. Она может рассматриваться как модель простейшей нервной системы. (ИОНТ РАН)
Получены принципиально новые алгоритмы поиска регуляторных сигналов в геномах бактерий, а также для поиска горизонтальных переносов генов в ходе их исторического развития, реализованные для последовательной и параллельной вычислительных архитектур. На этой основе с использованием алгоритма поиска альтернативных и консервативных вторичных структур РНК показано, что биосинтез рибофлавина и тиамина регулируется за счет образования альтернативных вторичных
структур РНК, причем один и тот же структурный механизм53 действует на разных функциональных уровнях – аттенюации (преждевременной терминации) транскрипции и ингибирования инициации трансляции. При анализе биотинового регулона впервые обнаружен сигнал регуляции транскрипции, общей для бактерий и архей. (ИППИ РАН)
Развита трехмерная математическая модель речевого тракта, найдены адекватные критерии оптимальности в решении динамической обратной задачи, обеспечившие существенное продвижение в восстановлении формы речевого тракта по измеренным акустическим параметрам речевого сигнала. Это решение необходимо для создания принципиально новых систем синтеза речи по тексту, автоматического распознавания речи и экономной передачи речевого сигнала в каналах связи. (ИППИ РАН)
Разработан и апробирован метод неинвазивной локации ишемических очагов в сердце на основе биофизического и информационного анализа электрокардиографических сигналов, регистрируемых при помощи предложенной модификации электрокардиографической измерительной системы Франка. Метод может быть использован для кардиологической диагностики, наблюдения за состоянием сердечно-сосудистой системы и как составная часть системы телемедицины. (ИППИ РАН)
Разработана информационная технология для исследования электрической активности мозга, устойчивая к возмущениям и позволяющая классифицировать функциональное состояние пациента и оценивать функциональную зрелость головного мозга. (СПИИРАН).
Внастоящее время начата работа на стыке нейроинформатики
ипсихологии - моделирование психики как специфически организованной системы информационных элементов. (Мирошников С.А., 2000).
Для описания взаимоотношений психики и соматики сформирована концепцию взаимозависимости, согласно которой два несовместимых описания должны использоваться, чтобы описать что-либо полностью. Автор рассматривает психическую деятельность и соматическую как два дополнительных понятия
(Walach H. 2005).
Врамках теории функциональной организации, развиваемой Chauvet G.A., предложен подход к пониманию старения основанный на понятии необратимости времени. Этот подход базируется на
эффекте физически маленьких, но многочисленных54 пертурбаций, известных как колебания, структурных единиц в процессе динамики биологической системы в течение ее взрослой жизни. Процессом старения управляют определенные гены, но необратимость времени реализуется вследствие деградации физиологических функций. В понимании автора для биологической системы, природа времени является определенной и высокоинтегрированной функцией. Внутренний физиологический возраст отражает необратимый курс живущего организма к смерти из-за необратимого курса физиологических функций к дисфункции, из-за необратимых изменений в регулирующих процессах. Данная проблема рассматривается в терминах отношений между макроскопической необратимостью функциональной организации и основных механизмов регулирования на самом низком "микроскопическом" уровне, то есть на молекулярном уровне - наиболее низком уровене организации. Предложен метод определения элементарного функционального взаимодействия с целью определения старения, основанный на исследовании нейроннейронального взаимодействия (Chauvet GA. 2006).
1.2.1. Системный анализ эволюции судорожных очагов и их отношений на биологической модели многоочаговой корковой фокальной эпилепсии
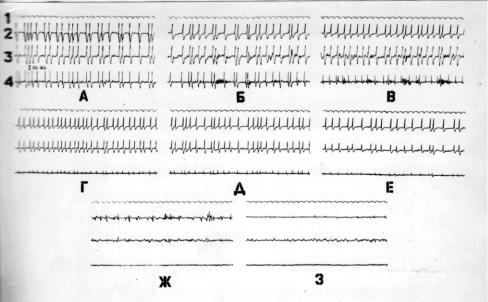
Многочаговую корковую фокальную эпилепсию моделировали аппликацией конвульсантов (стрихнина и пенициллина) в сенсомоторной коре мозга кошки (Лобасюк Б.А., 1988) (рис. 1.1.).
Зависимые судорожные очаги располагали в передней и задней сигмовидных извилинах, а детерминантный в средней сигмовидной извилине мозга кошки.
Функциональными показателями эпилептического многоочагового коркового комплекса являются: 1. Частота генерации судорожных потенциалов эпилептическими очагами; 2. Амплитуда судорожных потенциалов эпилептических очагов. Использованная в работе биологическая модель многоочагового эпилептического комплекса состояла из трех очагов: детерминантного, располагавшегося в средней сигмовидной извилине; первого зависимого, располагавшегося в задней сигмовидной извилине; второго зависимого, располагавшегося в
55
Рис. 1.1. Электрическая активность стрихнинового эпилептического комплекса при ОТСУТСТВИИ воздействий (контроль)
А–апликацией 0,1% стрихнина на зоны 3,4 и З% стрихнина на зону 2 создан эпилептический комплекс. Б, В, Г, Д, Е, Ж, 3 - через 10, 18, 46, 57, 68 и 72 минуты после А,
Обозначения: I - отметка времени, 2 - средняя сигмовидная извилина, 3 - задняясигмовиднаяизвилина,4–передняясигмовиднаяизвилина.Отметкавремени– 1с.,калибровка–500мкв.
передней сигмовидной извилине. Экспериментально определенные значения амплитуды судорожныых потенциалов обозначались: для детерминантного судорожного очага Ад; для первого зависимого – А1; для второго зависимого судорожного очага А2. Расчетные значения обозначались соответственно: Âд, Â1, Â2. Эпилептический комплекс состоявший из трех судорожных очагов, из которых один превосходит по мощности два других, можно рассматривать как упорядоченное множество (Стариков А.И., 1997), отдельные элементы которого (судорожные очаги генерирующие эпилептические потенциалы) единовременно являются функцией времени (t) и остальных элементов множества.
Сформированная прагматическая концепция эпилептического комплекса предопределила формализм использованных регрессионных моделей. Функциональные показатели эпилептических комплексов аппроксимировали во времени. В качестве апроксимирующих, были избраны линейная и квадратная функции.
Уравнение линейной регрессии:
56
Y’ = а0 + b1t+ e, где:
а0 – среднее значение Y, если независимая переменная t равна 0; b1 – среднее изменение Y на единицу измерения t.
е – погрешность, означающая любое колебание Y1, не вызванное изменением независимой переменной в модели.
Уравнение квадратной регрессии: Y’ = а0 + b1t + b2t2 + e,
При формировании математических моделей, исходя из допущения, что эпилептический многоочаговый корковый комплекс представляет собой упорядоченное множество эпилептических очагов, методом множественной линейной регрессии определяли взаимные ориентированные влияния судорожных очагов друг на друга, как элементов множества. С этой целью расчетные значения амплитуды судорожных потенциалов одного из эпилептических очагов, рассматривали, как функцию амплитуды судорожных потенциалов двух остальных судорожных очагов. Этот прием назван методом псевдоротации (Кузьмин В.Е., 1998; Мавродий, Коноваленко, Лобасюк, 1994; Лобасюк и др., 1995; Бабов, Лобасюк, Алексеенко, 1995; Золотарева, Лобасюк, Ручкина, 1995; Нагорная, Артеменко, Золотарева., Лобасюк, 1996; Лобасюк, Бабов, Алексеенко, 1996; Родионов Д.Б., Лобасюк Б.А., 2001; Олейник А.А., Гнатковский В.В., Лобасюк Б.А., Шандра А.А., 2001).
Таким образом, определяли: Âд= φ (А1 ; А2)
Â1= φ (Ад ; А2) Â2= φ (Ад ; А1)
Общая формула множественной регрессии:
Y’ = а0 + b1X1 + b2X2… +…bnXn + e.
Совокупность связей между тремя судорожными очагами эпилептичсекого комплекса описывалась тремя уравнениями множественной линейной регрессии вида:
Âд= а0 + b1А1 + b2А2 + e. Â1= а0 + b1Ад + b2А2 + e. Â2= а0 + b1Ад + b2А1 + e.
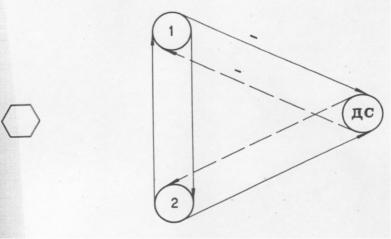
Использование трёх уравнений множественной линейной регрессии, описывающих связи между судорожными очагами эпилептического комплекса и трех уравнений простой линейной регрессии, описывающих изменения амплитуды судорожных потенциалов эпилептических очагов во времени, обеспечило возможность геометрической интерпретации эпилептического комплекса как многомерной системы в виде смешанного полициклического мультиграфа (Лобасюк Б.А., 1988).
Анализ математических моделей - уравнений линейной регрессии (рис. 1.2), аппроксимирующих изменения амплитуды судорожных

потенциалов эпилептических очагов во времени позволил57 сформировать математическое понятие «резистентность судорожного очага» и установить его меру. Оказалось, что в качестве меры резистентности можно использовать величину b1 – среднее изменение Y на единицу измерения t, (тангенс угла альфа), определяющего интенсивность уменьшения величины амплитуды судорожных потенциалов и теоретическое время существования судорожной ак тивности в эпилептических очагах (коэффициент С - отношение а0/b1).
Рис. 1.2. Диаграммы изменений средних величин частоты А амплитуды судорожных потенциалов в очагах эпилептического комплекса и теоретические линии регрессии, алпроксимирующие эти изменения во времени (контрольные опыты)
А - изменения частоты генерации судорожных потенциалов в эпилептических очагах а.=(0,206+0,13); b= (39,59±6,29);
N=13; r +0,63; t = 2,69; Р<0,0.Б - изменения амплитуды судорожных
судорожных потенциалов в детерминантном очаге
а.=(0,0151±0,0043); b= (2,64±0,17); с=(175±57)
N=13; r -0,91; t = 7,28; Р<0,001 В - изменения амплитудысудорожных потенциалов в первом зависимом
очаге
а.=(0,0243±0,002); b= (1,94±0,071); с=(80±7,23)
N=12; r -0,99; t = 22,19; Р<0,001. Г - изменения амплитуд судорожных потенциалов во втором зависимом очаге
58
а.=( 0,022±0,0038); b=(1,52±0,13); с=(69±13); n =12; r -0,97; t = 12,62; Р<0,00.
Следует подчеркнуть, что установление времени существования судорожной активности в эпилептических очагах экспериментально связано с трудностями.
Показана возможность экспериментального различения детерминантного и зависимых судорожных очагов эпилептического комплекса применением эфира или фторотана (Крыжановский Г.Н, Макулькин Р.Ф., Шандра А.А., 1978; Шандра А.А., 1977). У животных у которых был сформирован эпилептический комплекс аппликацией конвульсантов на кору мозга при общем угнетении возбудимости мозга применением эфира или фторотана внача
ле подавлялись зависимые судорожные очаги, причем вначале те из них, которые испытывали наименьшее влияние со стороны детерминантного очага, а детерминантный очаг подавлялся в последнюю очередь (Крыжановский Г.Н., 1980). Полученные результаты рассматриваются как доказательство превосходящей резистентности детерминантного судорожного очага над зависимыми.
Анализ регрессионных моделей аппроксимирующих изменения амплитуды судорожных потенциалов эпилептических очагов во времени позволил различить детерминантный и зависимые судорожные очаги комплекса и без применения эфира или фторотана. Интенсивность уменьшения амплитуды т.е. величина b1 в зависимых очагах была большей, чем в детерминантном, а теоретическое время существования судорожной активности (коэффициент С) – меньше.
Таким образом, иерархические отношения в эпилептическом комплексе в котором организующую роль играет детерминантный очаг могут быть выявлены как в эксперименте, так и при анализе математических моделей. Иерархические отношения являются одним из признаков системности (Уотермен Т., 1971, Месарович М.Д,. 1971). Следует отметить, что вопреки утверждению К.В. Судакова (1987) о том, что «иерархическое взаимодействие… всегда строится на основе открытого А.А. Ухтомским принципа доминанты», в эпилептическом комплексе иерархические отношения формируются на основе принципа детерминанты.
Как подчеркивает Г.Н. Крыжановский (2002) «внутренние свойства доминанты (повышенная возбудимость, суммация
возбуждений, подкрепление и др.) не являются её59 специфическими признаками, они присущи возбужденным структурам в ЦНС, которые, однако, могут не быть доминантными».
Сравнивая эволюцию отдельного судорожного очага, и такого судорожного очага, но являющегося элементом эпилептического множества, можно отметить его большую резистентность, судя по коэффициентам регрессии и теоретическому времени существования судорожной активности в таком очаге. Полученные результаты свидетельствуют о том, что одним из важных показателей системности любого уровня является её устойчивость
(Судаков К.В., 1984).
Использованная в работе биологическая модель многоочаговой корковой эпилепсии состояла из трех судорожных очагов, которые могут рассматриваться как элементы множества. Применением множественной линейной регрессии в стрихниновом эпилептическом комплексе при отсутствии воздействий (контрольные опыты) (рис. 1.3) определялись взаимные ориентированные влияния между судорожными очагами, т.е элементами множества на раннем этапе формирования эпилептического комплекса – первые тридцать минут эволюции и за весь период эволюции 60 минут. На первом этапе эволюции комплекса выявлялись статистически значимые отрицательное влияние первого зависимого очага, созданного применением 0,1% раствора стрихнина на судорожный очаг созданный применением 3% раствора стрихнина и положительное – второго зависимого на судорожный очаг созданный стрихнином высокой концентрации. Между судорожными очагами созданными 0,1% раствором стрихнина выявлялись двусторонние положительные влияния. Обратные влияния от мощного судорожного очага на судорожные очаги созданные слабой концентрацией стрихнина были подобны таковым от слабых судорожных очагов на мощный судорожный очаг, но статистически значимы при Р>0,1.
Учитывая, выявленные разнонаправленные влияния судо рожных очагов созданных применением слабых растворов стрихнина на судорожный очаг созданный применением -стрихнина высокой концентрации, можно предположить, что на первом этапе эволюции эпилептического комплекса судо-
60
Рис. 1.3. Математическая модель эпилептического комплекса за 30-ти минутный период эволюции Уравнения множественной линейной регрессии, отражающие ссвязи между очагами:
Aд= (2,197±0.181)+(-0,51±0,51)А1+(0,841±0,493)А2
N = 7; r = 0 , 74 ; t = 2 , 18; P < 0, 1> 0, 05 ; S y =0, 131 8 A1= (0,854±0,11)+(-0,196±0,665) Aд +(1,01±0,307)А2
n=7; r=- 0,97; t = 8,53; P< 0,002; S y = 0,03138. А2=(0,947±0,107)+(0,292±0,553)Ад + (0,906±0,301 )А1 n=7 r=0,98; t= 9,36; P< 0,001; S y = 0,07709,
рожный очаг, созданный применением стрихнина более высокой концентрации выступает в роли управляемой структуры (Уотерман Т., 1971).
На втором этапе эволюции эпилептического комплекса (рис. 1.4) выявлялось наличие двусторонних положительных связей между наиболее мощным судорожным очагом (детерминантным) и вторым зависимым очагом, а также между зависимыми очагами. Двусторонние положительные влияния могут быть рассмотрены как положительные петли обратной связи, являющиеся основой процесса детемирнации.
На первом этапе эволюции эпилептического комплекса между одним из судорожных очагов созданных 0,1% раствором стрихнина и мощным судорожным очагом созданным 3% рас твором стрихнина формируются доминантные отношения, а между другим судорожным очагом созданных 0,1% раствором стрихнина и мощным судорожным очагом созданным 3%