

Красильникова - Анатомия растений
.pdfа |
б |
в |
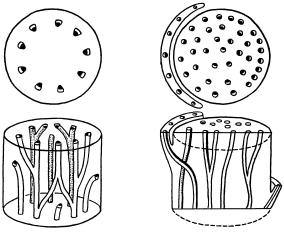
Рис. 107. Типы стел:
а — протостела; б — сифоностела; в — диктиостела
пcилофиты) и некоторых современных папоротников. Растения, имеющие протостелу, могут быть только небольших размеров, поскольку диаметр стелы очень мал (до 3 мм) и ее ксилема является слабой опорой стеблю (рис. 107).
Следующим этапом развития стелы явилась сифоностела, в центре которой образовалась сердцевина. Это более прогрессивная структура центрального цилиндра, так как, во-первых, за счет сердцевины увеличилась поверхность соприкосновения проводящих тканей с живыми паренхимными клетками, во-вторых, в результате перемещения прочной ксилемы на периферию стела превратилась в трубчатую конструкцию, что сделало ее более прочной и дало толчок развитию более высоких растений. Сифоностела встречается у папоротников.
Затем в процессе эволюции появилась диктиостела. Она сформировалась в результате прорывов сплошного кольца проводящих тканей паренхимными тяжами, идущими от сердцевины к первичной коре, иными словами, диктиостела распалась на отдельные проводящие пучки. Этому способствовало развитие листьев и боковых ветвей, проводящие системы которых при вхождении в стебель образуют листовые и веточные лакуны. В диктиостеле много паренхимы, с которой соприкасаются проводящие элементы: сердцевина, сердцевинные лучи, листовые и веточные лакуны, перицикл (рис. 108). Диктиостела характерна для папоротников.
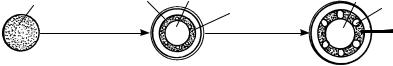
Дальнейшее эволюционное развитие сифоностелы и диктиостелы привело к образованию современной структуры стелы голосеменных и покрытосеменных растений — эустелы и атактостелы. Эустела характерна для голосеменных и многих двудольных. Она состоит из расположенных по окружности открытых коллатеральных проводящих
161
1
2
а |
б |
Рис. 108. Эустела (а) и атактостела (б):
1 — схема поперечного среза; 2 — объемное изображение
пучков и межпучковых тяжей паренхимы (сердцевинные лучи). У атактостелы однодольных закрытые коллатеральные пучки характеризуются сложным прохождением и на поперечном срезе стебля рассеяны без видимого порядка (рис. 108). Наиболее высокоорганизованными считаются стелы цветковых растений, содержащие большое количество паренхимы (многие травянистые растения).
Вторичное строение стеблей двудольных
Как уже отмечалось, в стеблях двудольных при первичном строении присутствует камбий, в результате работы которого образуются вторичные ткани и формируется вторичное строение стебля. Камбий возникает из прокамбия конуса нарастания стебля.
Образование прокамбия
В конусе нарастания ниже листовых бугорков начинается дифференцировка первичной коры и сердцевины, на что указывает вакуолизация клеток в зонах формирования этих частей стебля. Между дифференцированными слоями остается кольцо меристематических клеток, которые называют остаточной меристемой. В ней путем продольного деления и роста в длину образуются первые вытянутые прокамбиальные клетки.
162
1 2 3 3
4 |
5 |
|
|
|
6 |
а б в
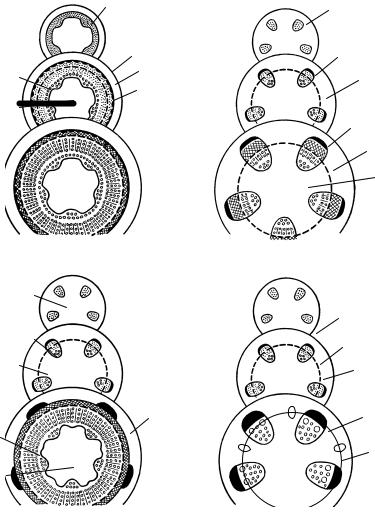
Рис. 109. Cхема образования прокамбия в конусе нарастания (поперечный срез):
а — меристема в точке роста; б — дифференцировка первичной коры и сердцевины; в — возникновение пучков прокамбия; 1 — образовательная ткань; 2 — первичная кора; 3 — сердцевина; 4 — остаточная меристема; 5 — пучки прокамбия; 6 — межпучковая паренхима
Далее в остаточной меристеме формируются прокамбиальные тяжи. Первые из них являются листовыми следами ближайших листовых зачатков (см. рис. 104). После формирования всех прокамбиальных пучков на данном уровне конуса нарастания участки остаточной меристемы между ними превращаются в межпучковую паренхиму. В узлах часть остаточной меристемы образует паренхиму листовых лакун. В кольце остаточной меристемы различных растений может образовываться разноеколичествопрокамбиальныхпучков:многомелких,небольшоечисло крупных или пучки могут соединяться в сплошное кольцо (рис. 109).
Типы заложения прокамбия и особенности строения стеблей
Русский ботаник С.П. Костычев в 1917—1920 гг. показал, что строение стеблей двудольных, в первую очередь их стелы, зависит от особенностей заложения прокамбия.
1.Из сплошного кольца прокамбия, сформированного в точке роста, образуется кольцо проводящих тканей: к периферии откладывается флоэма, к центру — ксилема. Между ними остается прослойка прокамбия, который дифференцируется в камбий и откладывает непрерывные слои вторичной флоэмы (луба) и вторичной ксилемы (древесины). В результате формируется стебель с непучковым (сплошным) строением стелы (рис. 110).
2.В случае пучкового заложения прокамбия каждый пучок образует первичные флоэму и ксилему. Образуется пучковый тип первичного строения стебля. Между пучками находится паренхима сердцевинных лучей. После тангентального деления клетки прокамбия
впучках дифференцируются в камбий, который называют пучковым. В то же время паренхима сердцевинных лучей под влиянием пучкового камбия приобретает способность делиться. Клетки ее вытягиваются, заостряются на концах и образуют камбий. В отличие от пучкового его
163
|
1 |
7 |
|
2 |
8 |
5 |
3 |
9 |
|
||
|
4 |
|
6 |
|
|
|
|
8 |
9
6
а |
в |
7
7
8
8
9
9
2 |
11 |
10
12
6
б |
г |
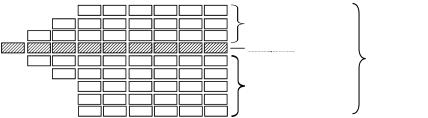
Рис. 110. Типы строения стеблей двудольных в зависимости от заложения прокамбия:
а — непучковый тип строения стебля, образовавшийся при непучковом заложении прокамбия; б — непучковый тип строения стебля, образовавшийся при пучковом заложении прокамбия; в — пучковый тип строения стебля, возникший при пучковом заложении прокамбия; г — промежуточный тип строения стебля; 1 — сплошное кольцо прокамбия; 2 — первичная кора; 3 — первичная флоэма; 4 — камбий; 5 — первичная ксилема; 6 — сердцевина; 7 — пучки прокамбия; 8 — проводящие пучки с пучковым камбием; 9 — межпучковый камбий; 10 — сплошное кольцо вторичного луба и вторичной древесины с камбиальной зоной между ними; 11 — крупные проводящие пучки; 12 — мелкие добавочные пучки
164
называли межпучковым. Таким образом, возникает сплошное камбиальное кольцо, состоящее из пучковых и непучковых участков. Затем пучковый камбий всегда формирует вторичные проводящие ткани,
амежпучковый может работать по-разному, отчего зависят типы вторичного строения стеблей:
а) межпучковый камбий функционирует так же, как и пучковый, образуя вторичные луб и древесину. В результате получается непучковый тип строения стебля, как и в случае работы сплошного прокамбиального кольца. Непучковый тип стебля имеют большинство древесных и некоторые травянистые растения (тысячелистник, вьюнок, гвоздика, барвинок, лебеда и др.) (рис. 110);
б) межпучковый камбий формирует паренхиму сердцевинных лучей, поэтому стебель при вторичном строении сохраняет пучковый тип (рис. 110). Такое строение центрального цилиндра встречается у трав (кирказон, тыква, клевер, чистотел и др.);
в) возможен переходной тип строения стебля. Он возникает в том случае, когда межпучковый камбий в широких сердцевинных лучах, кроме паренхимы, образует мелкие добавочные проводящие пучки. Эти пучки имеют ряд особенностей. Во-первых, они не связаны с листьями, т. е. являются специальными, во-вторых, в отличие от крупных они не имеют первичных тканей. Добавочные пучки отделены от крупных узкими сердцевинными лучами. По мере роста стебля крупные и мелкие пучки сливаются в сплошное кольцо. Такой тип строения можно видеть в стебле подсолнечника, топинамбура (рис. 110).
3.У некоторых растений межпучковый камбий не закладывается,
апучковый работает слабо. В таком случае проводящие пучки погружены в однородную паренхиму (лютиковые).
Работа камбия
После образования первичных флоэмы и ксилемы у двудольных между ними остается прослойка прокамбия, который превращается в камбий после тангентального (параллельно поверхности органа) деления прокамбиальных клеток. Камбий состоит из одного слоя тонкостенных вытянутых вдоль оси стебля клеток с заостренными концами. Этот слой, возникший из прокамбия, называют инициальным камбием. Каждая камбиальная клетка делится тангентально. Одна из двух дочерних клеток остается инициальной, способной к неограниченному числу делений, а другая после нескольких делений дифференцируется в элемент вторичного луба, если это происходит с клеткой, находящейся кнаружи от слоя камбия, или вторичной древесины, если превращению подвергается клетка, расположенная вовнутрь от камбиального кольца. При этом в сторону вторичной древесины обычно отчленяется большее количество (в 3—5 раз) производных кам-
165
Производные камбия, дифференцирующиеся в элементы флоэмы
Инициали |
Камбиальная |
|
|
|
зона |
Производные камбия, дифференцирующиеся в элементы ксилемы
Рис. 111. Схема камбиальной зоны
бия, поэтому слой вторичной древесины особенно у древесных растений гораздо более мощный, чем слой вторичного луба. Камбий вместе с примыкающими к нему с обеих сторон ближайшими производными клетками, сходными с ним по внешним признакам, называют камбиальной зоной. Собственно инициальный камбий в этой зоне представлен одним слоем клеток (рис. 111).
Удревесных растений инициальный слой камбия состоит из клеток двух типов: прозенхимных с заостренными концами и округлых паренхимных. Первые называют веретеновидными инициалями, вторые — лучевыми инициалями. Лучевые инициали образуют паренхиму сердцевинных лучей, которые у древесных растений называют лубо-
древесинными.
По мере роста стебля в толщину камбий испытывает натяжение по окружности, поэтому время от времени его клетки делятся радиально, что приводит к увеличению общей поверхности камбиального слоя. Кроме того веретеновидные инициали делятся поперечными перегородками, увеличивая количество радиальных рядов камбиальных клеток, а значит, и рядов элементов вторичных луба и древесины.
Лучевые инициали образуются из веретеновидных клеток путем их неравномерного поперечного деления. При делении поперечные перегородки смещаются к одному из концов клетки. Меньшая из образовавшихся дочерних клеток становится лучевой инициалью и далее, делясь поперечными и продольными перегородками, формирует многослойный многорядный луч (рис. 112).
В связи с тем, что камбий древесных растений состоит из инициалей двухтипов,егопроизводные—элементывторичногостроениястебля— различаются по структуре и ориентации по отношению к продольной оси органа. Из веретеновидных инициалей формируются продольные элементы, вытянутые вдоль органа (трахеи, трахеиды, ситовидные трубки, волокна), а из лучевых — поперечные, или радиальные (клетки лубодревесинных лучей).
Удревесных различают два типа камбия: ярусный и неярусный.
Вярусном концы камбиальных клеток располагаются приблизитель-
166

Рис. 112. Образование лучевых инициалей
но на одном уровне, а в неярусном — на разных. Первый тип камбия характерен для большинства лиственных деревьев (акация, ясень), а второй — для хвойных и некоторых лиственных (яблоня, груша). Неярусный камбий считается более давним и примитивным (рис. 113).
В связи с умеренным климатом камбий у древесных растений работает периодически — по сезонам. Активное деление камбиальных клеток начинается весной, летом замедляется, а осенью совсем прекращается. Значит отложение новых слоев вторичного луба и вторичной древесины также происходит посезонно.
1
2
а |
б |
Рис. 113. Типы камбия на тангентальном срезе стебля:
а — неярусный камбий; б — ярусный камбий; 1 — веретеновидные инициали; 2 — лучевые инициали
167
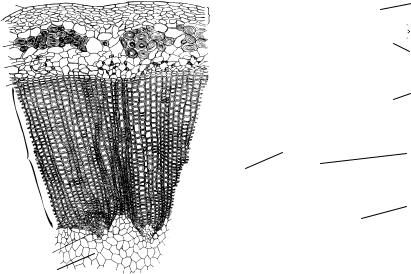
Вторичное строение стеблей травянистых растений
Анатомическое строение стеблей травянистых растений после вторичных изменений имеет свои особенности. Это, во-первых, наличие большого количества паренхимы (первичная кора, сердцевина, сердцевинные лучи); во-вторых, диаметр центрального цилиндра, значительно превышающий диаметр первичной коры; в-третьих, размещение механической ткани (чаще колленхимы) на периферии стебля.
Стебли трав, в том числе и многолетних, обычно живут один сезон, а затем отмирают, сохраняя при этом подземные органы (корневища, клубни). Поэтому в качестве покровной ткани у них функционирует эпидерма даже при вторичном строении.
Под эпидермой располагается паренхимная первичная кора. У многих растений в ней находится колленхима уголковая, пластинчатая или смешанная. Она придает травянистым стеблям прочность и упругость, что позволяет им выдерживать сильные удары ветра, даже те, которые ломают стволы деревьев.
Эндодерма превращается в крахмалоносное влагалище, а перицикл у травянистых растений обнаруживается далеко не всегда.
1 |
1 |
|
2 |
||
|
||
|
2 |
|
3 |
10 |
|
|
||
4 |
|
5
7
8
4
5 6 9
6 9 
7
 8
8
а |
б |
Рис. 114. Вторичное строение стеблей травянистых растений на поперечном срезе: а — лен; б — кирказон; 1 — эпидерма; 2 — первичная кора; 3 — пучки склеренхимных волокон; 4 — вторичная флоэма (луб); 5 — камбиальная зона; 6 — вторичная ксилема (древесина); 7 — первичная ксилема; 8 — сердцевина; 9 — сердцевинные лучи; 10 — кольцо механической ткани
168
8 |
12 |
3 |
10 |
6 |
9 |
|
2 |
|
|
|
|
|
1 |
|
|
|
|
|
|
|
4
5
 7
7
12 |
11 |
|
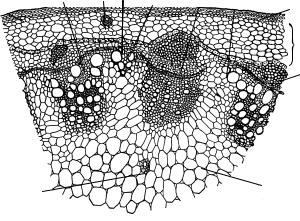
Рис. 115. Часть поперечного среза стебля подсолнечника:
1 — эпидерма; 2 — колленхима; 3 — эндодерма; 4 — первичная кора; 5 — пучки склеренхимы; 6 — флоэма; 7 — ксилема; 8 — пучковый камбий; 9 — межпучковый камбий; 10 — добавочные проводящие пучки, образованные межпучковым камбием; 11 — сердцевина; 12 — смоляные ходы
Как говорилось выше, в результате вторичных изменений в стеблях травянистых растений может формироваться пучковый или непучковый тип строения стелы. Пучковое строение имеет, например, стебель кирказона (рис. 114, б). Его отличает сплошное кольцо механической ткани на границе первичной коры и центрального цилиндра, а также широкие сердцевинные лучи. Непучковое строение характерно для стебля льна. Его особенностью являются пучки толстостенных лубяных волокон, которые служат сырьем для текстильной промышленности.
Всердцевине льна заметны выступы первичной ксилемы (рис. 114, а). Промежуточный тип строения стебля можно наблюдать у подсол-
нечника. Его особенностью является наличие мелких дополнительных проводящих пучков, которые размещаются между крупными основными и состоят только из вторичных флоэмы и ксилемы. В более старой части стебля пучки сливаются, образуя сплошное кольцо проводящих тканей (рис. 115).
В центре стебля находится паренхимная сердцевина, оставшаяся от первичного строения стебля.
Строение стебля древесных растений
В стеблях древесных растений камбий работает в течение всей их жизни, отчленяя все новые и новые слои элементов вторичного стро-
169

ения. Для древесных стеблей характерно присутствие большого количества мертвых клеток с одревесневшими стенками, что повышает их прочность и позволяет удерживать груз кроны. При этом большая часть стебля (до 90 %) занята вторичной древесиной.
Снаружи древесного стебля находится покровная ткань, под ней — первичная кора, далее — вторичный луб, за ним идет камбиальная зона. Внутрь от камбиальной зоны располагается мощная вторичная древесина, а в центре стебля — сердцевина. Через вторичную древесину и вторичный луб тянутся сердцевинные (лубодревесинные) лучи. Строение древесных стеблей удобно наблюдать на срезах (поперечном, радиальном, тангентальном) липы, яблони, бузины, груши, вяза, дуба, березы и др. (рис. 116).
9
8
1
 2
2
7
3
4
 10
10
5
8 
6
Рис. 116. Поперечный срез трехлетней ветки липы:
1 — перидерма; 2 — первичная кора; 3 — вторичный луб; 4 — камбиальная зона; 5 — вторичная древесина; 6 — сердцевина; 7 — лубодревесинные лучи; 8 — мягкий луб (ситовидные трубки); 9 — твердый луб (лубяные волокна); 10 — годичные кольца
170