

Лабораторна робота 6 матриця рам
Мета роботи – ознайомитись з поняттям матриці РАМ та навчитися програмувати розрахунок вагових матриць для різних еволюційних відстаней, оволодіти навичками використання програми для порівняння білкових послідовностей.
Теоретичні відомості
При порівнянні білкових послідовностей проста схема оцінювання ваг, така як +1 за збіг, -1 за розбіжність і -2 за пробіл, вже не ефективна. В процесі еволюції амінокислотні заміни відбувалися не рівномірно, а в залежності від біохімічних та інших властивостей самих амінокислот. Так, наприклад, вірогідність замін для амінокислот однакового розміру повинна бути більше, подібна гідрофобність/гідрофільність також повинна впливати на вірогідність заміни. Оскільки порівняння білкових послідовностей звичайно використовуються для виявлення еволюційних зв’язків, при побудові ваг необхідно враховувати ці міркування.
Насправді через складність та множинність різних факторів, що впливають на вірогідність заміни однієї амінокислоти іншою, кращий спосіб визначити її – це просто порахувати частоту замін для пари амінокислот. Стандартна процедура такого розрахунку заснована на важливому сімействі вагових матриць – так званих РАМ-матрицях (Point Accepted Mutation або Percent Accepted Mutation: відсоток реалізованих мутацій). За умови, що мутація не зачіпає один і той же сайт двічі, 1 РАМ – це час, еволюційний період, на протязі якого очікується одна заміна в поліпептидному ланцюзі зі 100 амінокислот.
Опис алгоритму
В програмах для розрахунку РАМ-матриць потрібно вибрати еволюційну відстань, на якій порівнюються дві послідовності. Матриці, в свою чергу, є функціями від відстані. Наприклад, для порівняння послідовностей, що знаходяться на відстані 250 еволюційних одиниць, потрібна матриця 250РАМ.
Для побудови матриці в 1 РАМ потрібно розрахувати матрицю вірогідності мутацій М та вагову матрицю S. Матриця М будується за наборами дозволених мутацій і вірогідностей появи кожної амінокислоти а, ра . Для одержання дозволених мутацій можна, наприклад, вирівняти два гомологічних білка у різних видів, тоді дозволені мутації будуть міститися в позиціях розбіжностей. Ми розглядаємо ці мутації як неспрямовані події, тобто, для пари амінокислот а, b, що вирівняли, невідомо – чи відбулася заміна a→b, або ж b→a. Можливо також, що початкова послідовність містила в цій позиції якусь третю амінокислоту, яка потім була замінена в одному випадку на а, в другому на b, але вірогідність такої події мінімізується вибором високогомологічних послідовностей. Для матриці 1 РАМ необхідно використовувати саме безпосередні мутації a→b, а не опосередковані, такі як а→с→b.
Вірогідності
появи різних амінокислот оцінюються
за частотою зустрічаємості на достатньо
великому диференційованому наборі
білкових послідовностей. Вони задовольняють
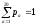 .
.
Далі
можна розрахувати частоти дозволених
мутацій
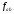 ,
при цьому
,
при цьому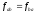 .
Сумарна
кількість мутацій по амінокислоті а:
.
Сумарна
кількість мутацій по амінокислоті а: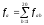 ,
,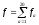 —сумарна
кількість амінокислот, що мутували, або
подвоєна кількість мутацій.
—сумарна
кількість амінокислот, що мутували, або
подвоєна кількість мутацій.
Тепер за частотами fab і вирогідностями ра можна побудувати 1 РАМ. Це матриця розміром 20х20 з елементами Мab, що рівні вірогідності амінокислотної заміни а на b. Відмітимо, що у випадку а=b, Маа відповідає вірогідності відсутності заміни у певний проміжок часу для амінокислоти а. Для РАМ матриць Маа розраховується за допомогою відносної мутабільності а, що визначається як ma=fa/(100fpa). Мутабільність – це вірогідність того, що за певний проміжок часу дана амінокислота буде замінена, так що вірогідність для а залишитись без змін розраховується за формулою.
Маа=1-ma.
З іншого боку, вірогідність заміни а на b можна розрахувати через добуток умовної вірогідності заміни а на b, за умови заміни а, помноженої на вірогідність заміни а. Умовну вірогідність можна оцінити як відношення кількості замін а↔b до загальної кількості мутацій за амінокислотою а, тобто: Маb-Pr(а↔b)=Pr(a→b|заміна а)=( fabma)/ fa.
Відмітимо, що всі розрахунки проводяться для спрощеної моделі еволюції білка. Припускається, що амінокислота а мутує незалежно як від еволюційної історії білку, так і від сусідніх амінокислот, які в принципі могли б впливати на мутабільність а.
Легко
показати, що
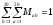 i
i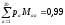 .
Перша рівність прямо вказує на те, що
для а сума вірогідності заміни на іншу
амінокислоту і вірогідності залишитись
без змін дорівнює 1. Нагадаємо, що ці
вірогідності відносяться до певного
еволюційного періоду. Одиницю еволюції
можна уявити собі як еволюційний період,
за який в середньому на 100 амінокислот
відбувається одна заміна. Це і є еволюційна
відстань в 1 РАМ. Матриця вірогідностей
переходів нормована таким чином, щоб
враховувати цей факт. Якби при розрахунку
відносної мутабільності а (ma)
в знаменнику було би використано,
наприклад 50 замість 100, то властивості
матриці не змінилися б, але одиниця
еволюції тепер вказувала на період, за
який в середньому вже на 50 амінокислот
відбувається 1 заміна.
.
Перша рівність прямо вказує на те, що
для а сума вірогідності заміни на іншу
амінокислоту і вірогідності залишитись
без змін дорівнює 1. Нагадаємо, що ці
вірогідності відносяться до певного
еволюційного періоду. Одиницю еволюції
можна уявити собі як еволюційний період,
за який в середньому на 100 амінокислот
відбувається одна заміна. Це і є еволюційна
відстань в 1 РАМ. Матриця вірогідностей
переходів нормована таким чином, щоб
враховувати цей факт. Якби при розрахунку
відносної мутабільності а (ma)
в знаменнику було би використано,
наприклад 50 замість 100, то властивості
матриці не змінилися б, але одиниця
еволюції тепер вказувала на період, за
який в середньому вже на 50 амінокислот
відбувається 1 заміна.
За матрицею М можна розрахувати вірогідності переходів для різних еволюційних відстаней. Наприклад, яка вірогідність того, що b замінить а через два еволюційних періоди? За перший період а може перейти в с з вірогідністю Mac, за другий с може перейти в bз вірогідністю Mcb. Складаючи, одержимо, що ця вірогідність дорівнює M2ab. В загальному випадку Мk це матриця перехідних вірогідностей для k еволюційних переходів. Слід відмітити, що при зростанні k (порядку тисячі) матриця М зходиться до матриці з однаковими стовпчиками, кожен з яких містить відносні вірогідності появи b, pb. Тобто, незалежно від «початкового наближення», після досить довгого еволюційного проміжку з вірогідністю pb, будь-яка амінокислота перейде в b.
Вагова матриця визначається наступним чином. Її значеннями є відношення вірогідності мутації a→b до вірогідності випадкової появи b, інакше кажучи, відношення правдоподібності Мab/pb. На практиці використовується десятинний логарифм відношення. Ми розглядали вагову матрицю для відстані в 1 РАМ, що легко може бути узагальнено на випадок k. Вагова матриця для відстані в k РАМ визначається як
scorek(a, b)=10∙log10(Mkab/pb).
Матриця симетрична внаслідок того, що розглядалися fab=fba.
Незважаючи на те, що ці матриці розраховуються тільки на наборах дозволених мутацій і частот зустрічаємості амінокислот, вони відображають загальні властивості амінокислот, наприклад, такі як гідрофобність/гідрофільність, заряд, розмір та ін. Звичайно амінокислоти з подібними властивостями мають високу вірогідність взаємозамін, що не дивно хоча б тому, що схожі амінокислоти частіше беруть участь в дозволених мутаціях. Заміни за принципом подібності дозволяють зберегтися функції білку.
Найчастіше для двох послідовностей нічого не відомо про еволюційну відстань. В таких випадках рекомендується порівнювати послідовності з використанням різних матриць відстаней, наприклад 40РАМ, 120РАМ і 250РАМ. В загальному випадку менші РАМ використовуються для пошуку коротких ділянок повної гомології, а більші – для виявлення довгих частково гомологічних фрагментів.