

Гиббереллины - гормоны листа
История открытия.
В Японии распространено заболевание риса, которое местные жители называют "баканоэ"-"бешеный рис", "дурные проростки". Рассада пораженных болезнью растений опережает в росте здоровый рис, но колосья вырастают уродливыми и зерна не бывает. В 1926 году японский ботаник Куросава выделил и описал возбудителя болезни - гриб Gibberella fujikuroi (сейчас этот гриб перенесли в род Fusarium). Вскоре выяснилось, что многие симптомы "бешеного риса" можно вызвать культуральной жидкостью, в которой рос гриб. Это значит, что гриб выделяет некоторое растворимое в воде вещество, усиливающее рост риса. По родовому названию гриба вещество было названо гиббереллином.
В 1930 году японские химики выделили гиббереллин в кристаллическом виде и предложили структурную формулу. Но эту работу прервала война и до 1950 года о гиббереллине почти никто не знал. Исследования возобновили несколько групп исследователей в Англии и США, и к 1955 году структура первого из гиббереллинов была окончательно установлена.
Обнаружилось, что растения и сами способны вырабатывать похожие вещества, за которыми сохранилось название гиббереллинов. Это - самый обширный класс растительных гормонов, на сегодня их известно более 100. Поскольку большинство из гиббереллинов - кислоты, их принято обозначать как ГК (гибберелловая кислота) с соответствующим индексом. Например: ГК24, ГК53 и т.д. Наиболее часто в экспериментах используют ГК3.
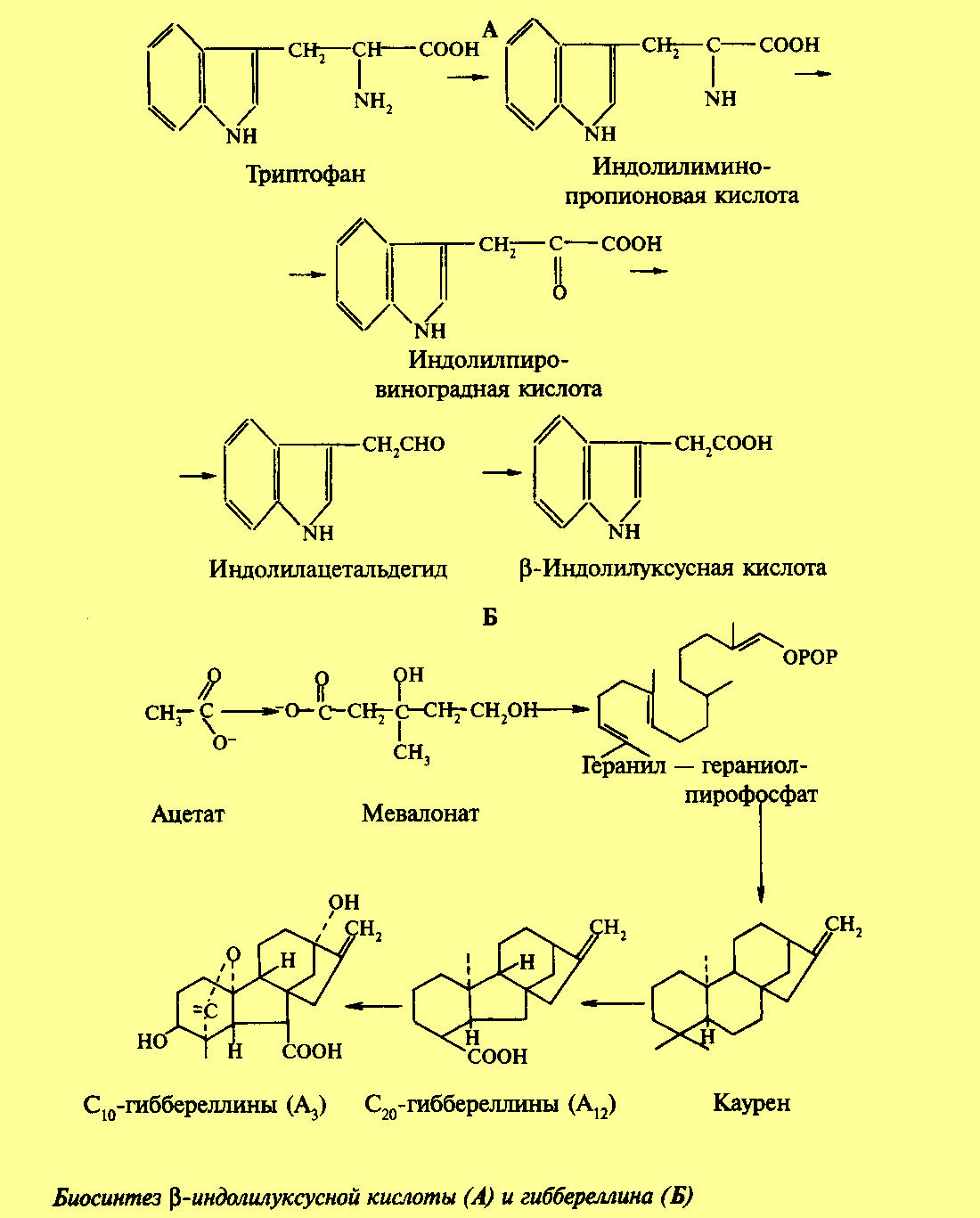
Биосинтез гиббереллинов.
В растениях биосинтез гиббереллинов начинается в пластидном компартменте. Из дезоксиксилозо-5-фосфата образуется изопентенилпирофосфат и далее (путем конденсации) геранилпирофосфат и геранилгераниолпирофосфат. Ключевым моментом биосинтеза гиббереллинов является циклизация последнего продукта с образованием энт-копалилдифосфата и энт-каурена, который считается предшесственником всех гиббереллинов.
Энт-каурен покидает пластиду и дальнейший синтез идет в цитоплазме. Последовательными реакциями окисления образуются энт-кауреновая кислота, энт-гидроксикауреновая кислота ГК12-альдегид и ГК12-кислота. После этого биосинтез гиббереллинов разветвляется на параллельные ветви, которые путем модификации радикалов и замыканием дополнительных циклов внутри молеккул приводят ко всему разнообразию гиббереллинов. Физиологически активными являются далеко не все гиббереллины. Физиологической активностью обладают в частности ГК1, ГК3, ГК4 , ГК7 и др., причем физиологическая активность зависит от видовой принадлежности растения. Одно и то же соединение может быть активно в одном виде растений, но не проявлять физиологической активност в другом.
Биосинтез гиббереллинов контролируется многими факторами. Например, начальные стадии биосинтеза находятся под контролем развития (т.е. включаются на определенных стадиях развития и дифференцировки). Переход от ГК12 или ГК53 к ГК9 или ГК20 зависит от длины дня и от уровня ауксинов. В этой точке метаболизма регулируется переход к цветению под действием гиббереллинов.
На переход от ГК9/20 к активным ГК4/1 влияют как ауксины, так и красный свет. Эта стадия биосинтеза находится под контролем при прорастании семян.
Рецепция гиббереллиновго сигнала приводит к замедлению указанных выше переходов, и одновременно усиливается превращение активных ГК4/1 в неактивные ГК34/8.
Как и большинство растительных гормонов, гиббереллины могут конъюгировать с сахарами, образуя неактивные гликозиды (запасные формы гиббереллинов). Кроме того, в растениях существуют специфические оксидазы, которые необратимо переводят гиббереллины в неактивные соединения.
О путях передачи гиббереллинового сигнала известно достаточно мало. Рецептор(ы), связывающиеся с гиббереллинами, пока еще не охарактеризованы. Показано, что в трансдукции сигнала участвует цГМФ.
Был выделен мутант арабидопсис spy (spindly) с конститутивным ответом на гиббереллин: растения сильно вытягивались в длину, семена преждевременно прорастали и т.п. Последовательность гена SPY позволяет сделать вывод, что белок работает как N-ацетилглюкозаминтрансфераза. По-видимому, гликозилирование является важным событием в репрессии ответа на гиббереллин. При повреждении белка репрессия снимается, и можно наблюдать постоянный ответ на гиббереллин, который не зависит от добавления гормона.
Еще один белок гиббереллинового ответа был выделен благодаря мутации gai (GA-insensitive). Он оказался транскрипционным регулятором. При повреждении генного локуса GAI растения становились нечувствительными к гиббереллинам.
Гиббереллины способны вызывать синтез специфических транскрипционных факторов, которые обозначены как GAMyb. Эти регуляторы узнают последовательности в промоторных участках многих генов (например, у гена a-амилазы ячменя - о роли этого фермента см. ниже).