

БИОФИЗИКА КУРС ЛЕКЦИЙ
.pdfется и протаскивает актиновую нить относительно миозиновой приблизительно на 10 нм (рис. 7.8, в).
Образовавшийся актин-миозиновый комплекс препятствует дальнейшему скольжению нитей относительно друг друга, поэтому необходимо его разъединение. Это возможно только за счет энергии АТФ. Миозин обладает АТФ-азной активностью, то есть, способен вызывать гидролиз АТФ. Выделяющаяся при этом энергия разрывает связь между актином и миозином (рис. 7.8, г), и миозиновая головка способна взаимодействовать с новым участком молекулы актина. Работа мостиков синхронизирована таким образом, что связывание, наклон и разрыв всех мостиков одной нити происходит одновременно.
При расслаблении мышцы активизируется работа кальциевого насоса, что понижает концентрацию Са2+ в цитоплазме; следовательно, связи между тонкими и толстыми нитями уже не могут образовываться. В этих условиях при растяжении мышцы нити беспрепятственно скользят относительно друг друга. Однако такая растяжимость возможна только в присутствии АТФ. Если в клетке отсутствует АТФ, то актин-миозиновый комплекс не может разорваться. Нити остаются жестко сцепленными между собой. Это явление наблюдается при трупном окоченении.
Существует два режима сокращения мышцы: изотоническое (изменяется длина волокна, а напряжение остается неизменным) и изометрическое (концы мышцы неподвижно закреплены, вследствие чего изменяется не длина, а напряжение).
7.3. Мощность и скорость одиночного сокращения.
Важными характеристиками работы мышцы являются сила и скорость сокращения. Уравнения, выражающие эти характеристики, были эмпирически получены А. Хиллом и впоследствии подтверждены кинетической теорией мышечного сокращения (модель Дещеревского).
Уравнение Хилла, связывающее между собой силу и скорость сокращения мышцы, имеет следующий вид
(Р + a)(v + b) = (P0 + a)b = a(vmах + b), |
(7.2) |
где v — скорость укорочения мышцы; Р — мышечная сила или приложенная к ней нагрузка (внешняя сила, действующая на мышцу); vmах – максимальная скорость укорочения мышцы (v = vmах при Р = 0); Р0 — сила, развиваемая мышцей в изометрическом режиме сокращения, то есть при такой нагрузке, при которой не происходит изменение длины мыш-
123

цы (P = Р0 при v = 0); а и b — константы. Приведенное
уравнение имеет вид гиперболы (рис. 7.9).
При сокращении за время t мышца совершает работу
А:
A = Pvt. |
(7.3) |
Выразив v из уравнения |
|
Хилла (7..2.), получаем: |
|
P0 – P |
|
A = bP |
t |
P + a
(7.4)
Рис. 7.9. Зависимость относительной скорости укорочения от относительной силы В процессе сокращения
мышцы выделяется некоторое количество теплоты Q. Эта величина называется теплопродукцией. Как показал Хилл, теплопродукция зависит только от изменения длины х мышцы и не зависит от нагрузки Р: Q = ax.
Общая мощность No6щ, развиваемая мышцей, определяется скоростями выполнения мышцей работы и выделения теплоты:
Рис. 7.10. Зависимость суммарной мощности портняжной мышцы лягушки от нагрузки при тетаническом изотоническом сокращении
|
dА |
|
dQ |
|
dQ |
|
Noбщ = |
|
+ |
|
= Pv + |
|
|
dt |
dt |
dt |
||||
|
|
|
(7.5)
Как следует из уравнения Хилла,
Noбщ = (Р + a)v = b(P – P0), |
(7.6) |
то есть зависимость мощности Noбщ от нагрузки Р является линейной
(рис. 7.10).
124

Коэффициент полезного действия мышцы η записывается в виде:
А |
Pv |
|
η = A + Q |
Nобщ |
(7.7) |
где Pv = Nпол — полезная мощность. КПД мышцы ε сохраняет постоянное значение (около 40 %). В диапазоне значений силы от 0,2Р0 до 0,8Р0.
Константы а и b имеют постоянные значения для данной мышцы. Константа а имеет размерность силы, а b — скорости. Константа b в значительной степени зависит от температуры, например, при нагревании на 10°С вблизи 0°С b удваивается. Константа а находится в диапазоне значений от 0,25Р0 до 0,4Р0. По этим данным легко оценить максимальную скорость сокращения для данной мышцы. Так как (следует из последнего равенства уравнения Хилла)
vmax = b |
P0 |
(7.8) |
P |
|
|
|
|
то, следовательно, vmax превышает константу b в 2,5—4 раза.
7.4.Электромеханическое сопряжение в мышцах.
Электромеханическое сопряжение - это цикл последовательных процессов, начинающийся с возникновения потенциала действия ПД на сарколемме (клеточной мембране) и заканчивающийся сократительным ответом мышцы. Нарушение последовательности процессов сопряжения может приводить к патологиям и даже к летальному исходу. Основные этапы этого процесса можно проследить по схеме (рис. 7.11.) Процесс сокращения кардиомиоцита происходит следующим образом (номера пунктов в тексте соответствуют номерам процессов в схеме электромеханического сопряжения на рис. 7.11):
1 - при подаче на клетку стимулирующего импульса открываются быстрые (время активации 2 мс) натриевые каналы, ионы Na+ входят в клетку, вызывая деполяризацию мембраны;
2 - в результате деполяризация плазматической мембраны в ней и в Т-трубочках открываются потенциал-зависимые медленные кальциевые каналы (время жизни 200 мс), и ионы Са2+ поступают из внеклеточной среды, где их концентрация = 2 • 10 –3 моль/л, внутрь клетки (внутриклеточная концентрация Са2+ ~ 10 –7 моль/л);
3 - кальций, поступающий в клетку, активирует мембрану СР, являющегося внутриклеточным депо ионов Са2+ (в СР их концентрация достигает = 10–3 моль/л), и высвобождает кальций из пузырьков СР, в ре-
125

зультате чего возникает так называемый "кальциевый залп". Ионы Са2+ из СР поступают на актин-миозиновый комплекс МФ, открывают активные центры актиновых цепей, вызывая замыкание мостиков и дальнейшее развитие силы и укорочения саркомера;
Рис. 7.11. Схема электромеханического сопряжения в кардиомиоците (М - клеточная мембрана-сарколемма, СР - саркоплазматический ретикулум, МФ - миофибрилла, Z - z-диски, Т - Т-система поперечных трубочек); 1 - поступления Na+ и 2 - поступления Са2+ в клетку при возбуждении мембраны, 3 - "кальциевый залп", 4 - активный транспорт Са2+ в СР, 5 - выход из клетки К+, вызывающий реполяризацию мембраны, 6 - активный транспорт Са2+ из клетки
4 - по окончании процесса сокращения миофибрилл ионы Са2+ с помощью кальциевых насосов, находящихся в мембране СР, активно закачиваются внутрь саркоплазматического ретикулума;
5 - процесс электромеханического сопряжения заканчивается тем, что К+ пассивно выходит из клетки, вызывая реполяризацию мембраны;
6 - ионы Са2+ активно выводятся во внеклеточную среду с помощью кальциевых насосов сарколеммы.
Таким образом, в кардиомиоците электромеханическое сопряжение идет в две ступени: вначале небольшой входящий поток кальция активирует мембраны СР, способствуя большему выбросу кальция из внутриклеточного депо, а затем в результате этого выброса происходит сокращение саркомера. Описанный выше двухступенчатый процесс сопряжения доказан экспериментально. Опыты показали, что: а) отсутствие потока кальция извне клетки jCa прекращает сокращение саркомеров, б) в условиях постоянства количества кальция, высвобождаемого из СР, из-
126

менение амплитуды потока кальция приводит к хорошо коррелирующему изменению силы сокращения. Поток ионов Са2+ внутрь клетки выполняет таким образом две функции: формирует длительное (200 мс) плато потенциала действия кардиомиоцита и участвует в процессе электромеханического сопряжения.
Следует отметить, что не во всех мышечных клетках организма процесс сопряжения происходит, как в кардиомиоците. Так, в скелетных мышцах теплокровных потенциал действия короткий (2-3 мс) и медленный поток ионов кальция в них отсутствует. В этих клетках сильно развита Т-система поперечных трубочек, подходящих непосредственно к саркомерам близко к z-дискам. Изменения мембранного потенциала во время деполяризации через Т-систему передается в таких клетках непосредственно на мембрану СР, вызывая залповое высвобождение ионов Са2+ и дальнейшую активацию сокращения (3, 4, 5). Временной ход описанных процессов показан на рис. 7.12.
Рис. 7.12. Временное соотношение между потенциалом действия кардиомиоцита (а) и одиночным сокращением (б) в этих клетках. Ордината слева - мембранный потенциал, справа - сила. φп - потенциал покоя
Общим для любых мышечных клеток является процесс освобождения ионов Са2+ и внутриклеточных депо - саркоплазматического ретикулума и дальнейшая активация сокращения. Ход кальциевого выброса из СР экспериментально наблюдается с помощью люминесцирующего в присутствии ионов Са2+ белка экворина, который был выделен из светящихся медуз. Задержка начала развития сокращения в скелетных мышцах составляет 20 мс, а в сердечной - несколько больше (до 100 мс).
8. БИОФИЗИКА НЕРВНОГО ИМПУЛЬСА
Нормальное функционирование организма невозможно без обмена информацией между клетками, одним из способов которого является возможность генерации и восприятия клетками нервного импульса. В организме существуют так называемые возбудимые клетки, к которым относятся мышечные, нервные и секреторные. Эти клетки способны откликаться каким-либо образом на их возбуждение. Так, мышечные клет-
127

ки сокращаются, секреторные выделяют биологически активные вещества, а нервные генерируют электрические колебания — нервный импульс.
8.1. Потенциал покоя.
На мембране клетки всегда существует разность потенциалов, то есть электрический потенциал внутри клетки отличается от наружного. Эта разность потенциалов называется потенциалом покоя. Если наружный потенциал принять равным нулю, то внутренний составит от -50 до - 90 мВ, в зависимости от вида клеток.
Причиной возникновения потенциала покоя является наличие градиента концентраций ионов К+, Na+ и С1– на клеточной мембране (табл. 8.1). Допустим, что мембрана проницаема только для ионов калия и непроницаема для остальных, тогда потенциал покоя будет определяться равновесным калиевым потенциалом и описываться уравнением Нерн-
ста:
Δθ = |
RT |
ln |
[K+]o |
(8. |
F |
+ |
|||
|
|
[K ]i |
1) |
|
|
|
|
|
где R — универсальная газовая постоянная; Т — абсолютная температура; F— число Фарадея; [К+]0 и [К+]i— наружная и внутренняя концентрации ионов калия соответственно. Если внутриклеточная концентрация К+ в 50 раз превышает внеклеточную, то потенциал покоя при температуре 27 °С составляет -100,7 мВ.
Величина потенциала покоя всегда несколько ниже равновесного калиевого потенциала, что объясняется просачиванием внутрь клетки ионов натрия, которые приносят туда свой положительный заряд. Диффузия отрицательно заряженных ионов хлора, наоборот, способствует увеличению Δθ. Таким образом, вклад в формирование потенциала покоя вносят потоки всех ионов, коэффициенты проницаемости которых отличны от нуля.
Вычислим зависимость потенциала покоя от концентрации и коэффициентов проницаемости ионов. В дальнейших расчетах будем учитывать только ионы К+, Na+, Cl–.
В стационарном состоянии сумма потоков ионов через мембрану
равна нулю: |
|
JNa +JK – JCl = 0 |
(8 |
.2)
хотя потоки отдельных ионов могут быть и отличны от нуля. Так как ионы хлора заряжены отрицательно, то перед потоком ионов хлора JС1 ставится знак «-».
128
Таблица 8.1 Концентрация ионов К+, Na+ и С1– внутри и снаружи клетки, значения равновесных потенциалов, потенциалов покоя и потенциалов дейст-
вия некоторых клеток
|
Клетки |
Значения концентраций ио- |
|
Равновесный |
Измеренные по- |
||||||||||||||||||||
|
|
|
нов внутри (i) и снаружи (о) |
|
потенциал для |
тенциалы, мВ |
|||||||||||||||||||
|
|
|
|
|
|
|
клетки, мМ |
|
разных ионов, мВ |
|
|
|
|
||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
Кi+/Кo+ |
Nai+/Nao+ |
|
Cli–/Clo– |
|
К+ |
|
|
Na+ |
Сl– |
покоя |
на макси- |
|||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
муме по- |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
тенциала |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Гигантский |
340/10,4 |
49/463 |
|
114/592 |
|
-88 |
+57 |
-42 |
-60 |
|
|
+ 50 |
||||||||||||
|
аксон |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Гигантский |
360/10,0 |
69/425 |
|
157/496 |
|
-90 |
+46 |
-29 |
-60 |
|
|
+ 35 |
||||||||||||
|
аксон каль- |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Мышечное |
(48:1) |
(1:7) |
|
(1:64) |
|
-98 |
+49 |
-105 |
-88 |
|
|
+ 34 |
||||||||||||
|
волокно |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Моторный |
150/5,5 |
15/150 |
|
9/125 |
|
-90 |
+60 |
-70 |
-90 |
|
|
+ 30 |
||||||||||||
|
нейрон кош- |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
ки |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Выразим из уравнения Гольдмана (см. формулу 6.26) потоки для |
||||||||||||||||||||||||
|
ионов К+, Na+, Cl– с учетом их заряда z. |
|
|
|
|
|
|
|
|
|
|
||||||||||||||
|
|
|
|
|
FΔθРK [К+]i e FΔθ/RT – [К+]o |
|
|
|
|
(8.3) |
|||||||||||||||
|
JK = |
|
RT |
|
|
· |
|
|
|
1 – e FΔθ/RT |
|
|
|
|
|
|
|
||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||
|
|
|
|
|
FΔθРNa |
|
|
[Na+]i e FΔθ/RT – [Na+]o |
|
|
|
|
(8.4) |
||||||||||||
|
JNa = |
|
RT |
|
|
· |
|
|
|
|
1 – e FΔθ/RT |
|
|
|
|
|
|
|
|||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||
|
|
|
|
|
FΔθРCl |
|
|
|
[Cl–]i e FΔθ/RT – [Cl–]o |
|
|
|
|
(8.5) |
|||||||||||
|
JCl = |
|
RT |
|
|
· |
|
|
|
1 – e FΔθ/RT |
|
|
|
|
|
|
|
||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||
|
и подставим |
их |
в |
уравнение (8.2). |
Сократив на |
общий множитель |
|||||||||||||||||||
|
FΔθ/RT, запишем: |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
PK[К+]i e FΔθ/RT – PK[К+]o |
+ |
PNa[Na+]i e FΔθ/RT – PNa[Na+]o |
|
+ |
(8.6) |
||||||||||||||||||||
|
1 – e FΔθ/RT |
|
|
|
|
|
|
|
|
|
|
1 – e FΔθ/RT |
|
|
|
|
|||||||||
+ |
|
PCl[Cl–]ie FΔθ/RT – PCl[Cl–]o |
= 0; |
|
|
|
|
|
|||||||||||||||||
|
|
|
|
|
1 – e FΔθ/RT |
|
|
|
|
|
|
|
|||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
129 |
|
|
|
|
|
|
|
|
|
|
||
Разделив числитель и знаменатель последнего слагаемого на множитель e FΔθ/RT:
PK[К+]i e FΔθ/RT – PK[К+]o |
+ |
|
PNa[Na+]i e FΔθ/RT – PNa[Na+]o |
+ |
(8.7,а) |
||||||||||||||
|
|
|
|
|
|
|
|
|
1 – e FΔθ/RT |
|
|||||||||
|
1 – e FΔθ/RT |
|
|
|
|||||||||||||||
+ |
|
PCl[Cl–]i – PCl[Cl–]oe FΔθ/RT |
|
= 0; |
|
|
|
|
|
||||||||||
|
|
|
|
|
e FΔθ/RT – 1 |
|
|
|
|
|
|
||||||||
получаем: |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||
PK[К+]i e FΔθ/RT – PK[К+]o + PNa[Na+]i e FΔθ/RT – PNa[Na+]o – |
(8.7,б) |
||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
– |
PCl[Cl–]i – PCl[Cl–]oe FΔθ/RT = 0; |
|
|
|
||||||||||||||
или |
|
|
|
PK[К+]о + PNa[Na+]o + PCl[Cl–]i |
|
|
|
||||||||||||
|
FΔθ |
|
|
|
|
|
(8.7,в) |
||||||||||||
|
|
|
= ln |
|
|
|
|
|
|
|
|
||||||||
|
RT |
|
PK[К+]i + PNa[Na+]i + PCl[Cl–]o |
|
|
|
|
|
|||||||||||
В результате получаем: |
|
|
|
|
|
|
|
|
|
||||||||||
Δθ = |
RT |
|
PK[К+]о + PNa[Na+]o + PCl[Cl–]i |
|
|
|
|||||||||||||
|
|
ln |
|
|
|
|
|
(8.8) |
|||||||||||
F |
|
|
PK[К+]i + PNa[Na+]i + PCl[Cl–]o |
|
|
|
|||||||||||||
Данное |
|
выражение |
называется |
уравнением |
Гольдмана— |
||||||||||||||
Ходжкина—Катца.
В состоянии покоя мембранные проницаемости для ионов К+, Na+ и С1– относятся друг к другу как
К+ : Na+ : С1– = 1 : 0,004 : 0,45 (8.9)
Так как в покое проницаемость клетки для ионов калия намного больше ее проницаемости для других ионов, то потенциал покоя определяется преимущественно разностью концентраций ионов калия.
Поддержание разности концентраций ионов осуществляется при помощи работы ионных насосов, использующих энергию АТФ. Na+,K+- нacoc, кроме того, способствует увеличению трансмембранного потенциала, так как выводит три иона натрия и закачивает в клетку только два иона калия, что увеличивает положительный заряд межклеточной среды, а следовательно, увеличивает потенциал покоя.
Увеличение трансмембранной разности потенциалов называется ги-
перполяризацией, уменьшение — деполяризацией. В последнем случае может даже измениться знак внутриклеточного потенциала.
Нарушение работы АТФ-азы, которое можно вызвать, например, действием сердечного гликозида уабаина, уменьшает потенциал покоя. К такому же эффекту приводит действие различных ядов (например батра-
130
хотоксина), увеличивающих проницаемость клеточной мембраны для ионов натрия. Деполяризацию вызывает также торможение процессов, обеспечивающих в клетке синтез АТФ.
8.2. Потенциал действия.
При воздействии на клетку какого-либо раздражителя ее трансмембранный потенциал изменяется, возникает так называемый потенциал действия, или спайк. Причиной такого колебания потенциала покоя является изменение проницаемости мембраны для натрия, что, в свою очередь, вызвано открытием натриевых ионных каналов. При возбуждении проницаемости мембраны для ионов калия и натрия относятся друг к другу как
Рк : PNa = 1 : 20. |
(8.10) |
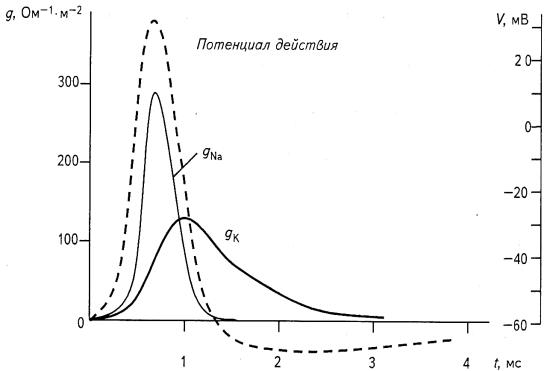
Рис. 8.1. Зависимость проводимости мембраны для ионов натрия и калия (gNa и gK) от времени t для гигантского аксона кальмара во время генерации потенциала действия V
В результате поток ионов натрия в клетку начинает превышать поток ионов калия из клетки (рис. 8.1). Если раньше потенциал на мембране был близок к равновесному калиевому потенциалу, то теперь он стремится к равновесному натриевому, но не достигает его только вследствие того, что проводимости для ионов калия и хлора отличны от нуля.
131

В клетке происходит деполяризация мембраны: отрицательный потенциал клетки приближается к нулю, а потом и вовсе меняет знак на противоположный (рис. 8.2). Последний процесс называется реверсией мембранного потенциала. Максимальное значение потенциала действия составляет обычно 30...40 мВ. Увеличение натриевой проводимости длится доли миллисекунд. Далее она начинает снижаться, а калиевая — возрастать, в результате чего в клетке восстанавливается потенциал покоя. Этот процесс называется реполяризацией клеточной мембраны.
Рис. 8.2. Фазф потенциала действия:
1 – фаза деполяризации; 2 – фаза реполяризации; 3 – следовой потенциал: а – отрицательный, б – положительный.
Для восстановления потенциала покоя клетке требуется некоторое время. В процессе реполяризации сначала происходит быстрое приближение к нормальному значению потенциала покоя, затем скорость изменения потенциала уменьшается, и клетка некоторое время пребывает в деполяризованном состоянии. Это называется следовой деполяризацией. В некоторых клетках, наоборот, возникает следовая гиперполяризация, то есть в процессе реполяризации разность потенциалов начинает превышать обычную. Оба эти отклонения от нормального значения потенциала покоя называются следовыми потенциалами (рис.8.2).
Длительность потенциала действия отличается для различных клеток (колеблется в пределах от 0,5 до 3 мс) и существенно зависит от температуры. При ее уменьшении на 10°С время существования потенциала действия увеличивается в три раза. Длительность реполяризации обычно превышает длительность деполяризации.
132