

Ретикулярная формация имеет связи с многочисленными структурами ЦНС. Афферентные входы поступают в латеральные области РФ преимущественно от трех источников:
-
температурных и болевых рецепторов по волокнам спиноретикулярного тракта и тройничного нерва импульсация поступает в ретикулярные ядра продолговатого мозга и моста;
-
сенсорной и, частично, других зон коры головного мозга по кортико-ретикуляр- ным путям импульсация поступает в ядра, дающие начало ретикулоспинальным трактам (гигантоклеточное ядро, оральное и каудальное ядра моста), а также в ядра, которые проецируются на мозжечок (парамедианное ядро и ядро покрышки моста);
-
ядер мозжечка по мозжечково-ретикулярному пути импульсация поступает в гигантоклеточное и парамедианное ядра и ядра моста.
Эфферентные выходы формируются преимущественно в медиальных областях РФ и
проецируются:
-
к спинному мозгу по латеральному рети- кулоспинальному тракту (от гигантокле- точного ядра) и по медиальному ретику- лоспинальному тракту (от каудального и орального ядер моста);
-
к верхним отделам головного мозга (не специфическим ядрам таламуса, заднему гипоталамусу, полосатому телу) идут вос ходящие пути, начинающиеся в ядрах продолговатого мозга (гигантоклеточном, латеральном и вентральном) и в ядрах моста;
-
к мозжечку идут пути, которые начинают ся в латеральном и парамедианном ретикулярных ядрах и в ядре покрышки моста. Многообразие связей и структур РФ обусловливает ее многочисленные функции. Эти функции можно объединить в триглавные группы: соматические — двигательные, сенсорные восходящее влияниена большой мозг) и вегетативные.
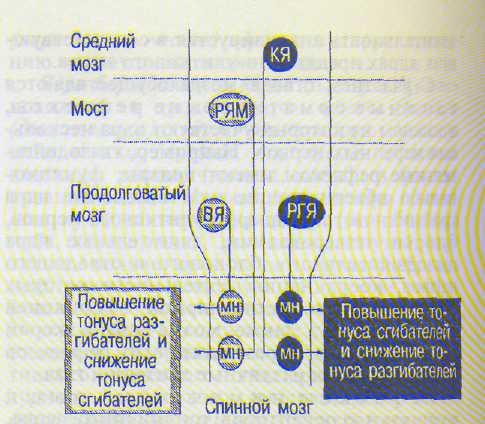
Рис. 8.5. Основные двигательные центры ствола мозга и двигательные пути, регулирующие тонус мышц туловища и конечностей.
КЯ — красное ядро и руброспинальный тракт; РЯМ — ретикулярные ядра моста и медиальный ретикулоспиналь-ный тракт; ВЯ — вестибулярные ядра и вестибулоспи-нальный тракт; РГЯ — ретикулярное гигантоклеточное ядро и латеральный ретикулоспинальный тракт; МН -спинальные мотонейроны.
Соматические функции РФ проявляются, во-первых, в ее влиянии на двигательные ядра черепных нервов, моторные спинальные центры и активность мышечных рецепторов (мышечных веретен).
Ретикулярная формация среднего мозга осуществляет координацию функции ядер глазодвигательных (111, IV, VI) нервов, обеспечивая содружественные движения глаз в горизонтальном и вертикальном направлениях. В парамедианной РФ продолговатого мозга имеются нейроны, управляющие горизонтальными движениями глаз. Эти же нейроны, но опосредованно, через нейроны РФ среднего мозга, управляют вертикальными движениями глаз. При двусторонних поражениях этих отделов РФ становится невозможным произвольное выполнение горизонтальных и вертикальных движений глаз.
Нисходящие влияния РФ на моторные спинальные центры (рис. 8.5). От ретикулярного гигантоклеточного ядра (РГЯ) продолговатого мозга идет частично перекрещенный латеральный ретикулоспинальный тракт, волокна которого оканчиваются на вставочных нейронах спинного мозга. Через эти интернейроны они возбуждают а- и у-нейроны мышц—сгибателей конечностей и реципрок-но тормозят нейроны мышц—разгибателей.
124
Основным источником лиановидных волокон являются нейроны нижней оливы продолговатого мозга. Информация к ним поступает от мышечных, кожных рецепторов и нейронов основания задних рогов спинного мозга (по спинооливному пути), а также от двигательной коры головного мозга. Каждое лиановидное волокно устанавливает еинаптические контакты с дендритами (медиатор аспартат), как правило, одной клетки Пуркинье, но число их может достигать трехсот, что объясняет исключительно сильное возбуждающее действие этого афферентного входа (большой ВПСП, на вершине которого формируется 3—5 потенциалов действия). Кроме того, они оказывают на них и тормозное (более слабое) влияние через корзинча-тые и звездчатые клетки поверхностного (молекулярного) слоя коры мозжечка.
По моховидным волокнам в кору мозжечка поступает информация от коры большого мозга (преимущественно ассоциативной, по корково-мостомозжечково-му пути), проприорецепторов опорно-двигательного аппарата (по спинно-мозжечковым путям), вестибулярных рецепторов и ретикулярной формации. Моховидные волокна образуют возбуждающие синапсы на дендритах клеток-зерен внутреннего (гранулярного) слоя коры. Через аксоны клеток-зерен этот афферентный вход оказывает непосредственное возбуждающее влияние (медиатор — глу-тамат) на тормозные клетки Пуркинье и опосредованно (через корзинчатые и звездчатые клетки) тормозное действие на клетки Пуркинье. Активность клеток-зерен регулируется через тормозные клетки Гольджи внутреннего слоя по типу возвратного торможения (медиатор — ГАМ К).
Третью, афферентную (моноаминергичес-кую) систему мозжечка начали изучать в последнее время. Через эти пути передается влияние на мозжечок некоторых структур ствола головного мозга, например голубого пятна и ядер шва. Нейроны голубого пятна осуществляют влияние на мозжечок через медиатор норадреналин, а ядер шва — через серото-нин. Стимуляция этих ядер тормозит фоновую активность клеток Пуркинье и модулирует их ответы на активацию моховидных и лиановидных афферентов. Как прямое (через лиановидные волокна), так и опосредованное (через моховидные волокна и клетки-зерна) афферентное влияние на клетки Пуркинье является возбуждающим.
Но поскольку клетки Пуркинье являются тормозными нейронами (медиатор ГАМК), то с их помощью кора мозжечка превращает
возбуждающие сигналы на входе в тормозные сигналы на выходе. Таким образом, эфферентное влияние коры мозжечка на последующее нейронное звено (в основном это внутримозжечковые ядра) осуществляется не по механизму запуска нейронной активности, а по механизму более или менее сильного сдерживания этой активности, т.е. в мозжечке доминирует тормозный характер управления [Фанарджян В.В., 1992].
При изучении связей коры мозжечка с его ядрами были выделены три симметричные вертикальные зоны. Клетки Пуркинье медиальной червячной зоны и флоккулонодулярной доли проецируются на ядра шатра. Нейроны этих ядер связаны с моторными центрами ствола, от которых к спинальным моторным центрам идут вестибуло-, рубро- и ретику-лоспинальные тракты, регулирующие тонус мышц. Клетки Пуркинье промежуточной зоны мозжечка проецируются на промежуточные (шаровидное и пробковидное) ядра. Аксоны нейронов этих ядер идут к красному ядру среднего мозга, от которого начинается руброспинальный тракт, стимулирующий через спинальные моторные центры тонус мышц-сгибателей.
Есть путь от промежуточного ядра к вент-ролатеральному ядру таламуса и оттуда к двигательной коре. Клетки Пуркинье латеральной зоны коры проецируются на зубчатые ядра мозжечка, от которых мощные тракты направляются к вентролатеральному ядру таламуса, нейроны которого в свою очередь проецируются на моторные зоны коры большого мозга. Итак, три парных ядра мозжечка являются главным эфферентным выходом мозжечка на стволовые и корковые моторные центры. Нейроны этих ядер имеют высокую фоновую активность (50—100 имп/с). Они находятся под тормозным влиянием клеток Пуркинье и возбуждающим влиянием афферентных входов, которые по коллатеральным ответвлениям поступают в эти ядра.
8.4.2. ФУНКЦИИ МОЗЖЕЧКА
Функции мозжечка формируют три главных его влияния на организм [Орбели Л.А., 1940]: на двигательный аппарат, афферентные системы и вегетативную нервную систему.
А. Двигательные функции мозжечка заключаются в регуляции мышечного тонуса, позы и равновесия (1), координации позы и выполняемого целенаправленного движения (2), программировании целенаправленных движений (3).
133
-
Регуляция мышечного тонуса, позы и равновесия осуществляется преимуществен но древним мозжечком (флоккулонодуляр- ная доля) и частично старым мозжечком, входящими в медиальную червячную зону. Получая и обрабатывая импульсацию от вес тибулярных рецепторов (по вестибуломоз- жечковому пути), от проприорецепторов ап парата движения и рецепторов кожи (по спинно-мозжечковым путям), от зрительных и слуховых рецепторов (через четверохолмие по тектоцеребеллярным путям), мозжечок способен оценить состояние мышц, положе ние тела в пространстве и через ядра шатра, используя вестибуло-, ретикуло- и руброс- пинальный тракты, произвести перераспре деление мышечного тонуса, изменить позу тела и сохранить равновесие. Нарушение равновесия является наиболее характерным симптомом поражения архицеребеллума. Из связей древнего мозжечка со структурами ствола мозга наиболее выражены его двусто ронние связи с вестибулярными ядрами. Мозжечок осуществляет тонкую настройку вестибулярных рефлексов, в том числе реф лекторное поддержание антигравитацион ной (вертикальной) позы. Функцию древне го мозжечка врач оценивает, определяя рав новесие у пациента в позе Ромберга: верти кальная поза со сдвинутыми ступнями и за крытыми глазами, вытянутыми горизонталь но (вперед) руками.
-
Координация позы и выполняемого целе направленного движения осуществляется ста рым и новым мозжечком, входящими в про межуточную (околочервячную) зону. В кору этой части мозжечка поступает импульсация от рецепторов аппарата движения, а также импульсация от моторной коры (программа произвольного движения). Анализируя ин формацию о программе движения (из мотор ной коры) и выполнении движения (от про приорецепторов), мозжечок способен через свое промежуточное ядро, имеющее выходы на красное ядро и моторную кору, осущест вить координацию позы и выполняемого це ленаправленного движения в пространстве, а также исправить направление движения. Об этом свидетельствует то, что нейроны про межуточного ядра продолжают генерировать импульсы в течение всего времени выполне ния движения. Нарушение координации дви жения (атаксия) является наиболее характер ным симптомом нарушения функции про межуточной зоны мозжечка. Эта фун кция мозжечка может быть исследована, напри мер, палыхеносовой или пяточно-коленной пробами.
-
Участие в программировании целена правленных движений осуществляется новым мозжечком — той частью полушарий мозжеч ка, которая входит в латеральную зону. Кора этой части мозжечка получает импульсацию преимущественно из ассоциативных зон коры большого мозга через ядра моста. Эта информация характеризует замысел движе ния. В коре нового мозжечка (а также в ба- зальных ганглиях) она перерабатывается в программу движения, которая через зубчатое ядро мозжечка и вентральное латеральное ядро таламуса попадает в кору, обрабатывает ся в премоторной и моторной коре большого мозга и через пирамидную, экстрапирамид ную системы осуществляется как сложное целенаправленное движение. Контроль и коррекция более медленных программиро ванных движений осуществляется мозжечком на основе обратной афферентации преиму щественно от проприорецепторов, а также от вестибулярных зрительных, тактильных ре цепторов. Коррекция быстрых (баллистичес ких) движений из-за малого времени их вы полнения осуществляется по другому меха низму. Если не достигается результат, кор рекция движений осуществляется путем из менения их программы в латеральном моз жечке, т.е. на основе обучения и предшеству ющего опыта. Об этом говорит и прекраще ние импульсации в зубчатых ядрах мозжечка сразу после начала движения. К таким дви жениям относятся многие спортивные дви жения (например, бросок мяча), игра на му зыкальных инструментах, «слепой» метод пе чатания и др. Врач может оценить эту функ цию мозжечка пробой с адиадохокинезом (например, быстрой сменой ротационных движений рук), при которой необходимо бы стро менять программу движения.
-
Наряду с этим полушария мозжечка осуществляют функцию инициации движения. Было показано, что изменение активности нейронов мозжечка (зубчатое и промежуточ ное ядра, клетки Пуркинье) на 0,1—0,3 с предшествует началу движения, а охлаждение зубчатого ядра задерживает на 0,1 с актива цию нейронов моторной коры и начало дви жения. Эти данные объясняют затруднения вызова движений, которые испытывают больные с поражением мозжечка. Вместе с тем основные симптомы нарушения функ ций мозжечка в большей или меньшей степе ни выявляются при поражении каждого из трех отделов мозжечка, что свидетельствует об их функциональном перекрытии.
Из изложенного выше видно, что нейроны мозжечка не имеют прямого выхода на спи-
134
нальные мотонейроны, а действуют на них через корково-стволовые моторные центры. С этим, вероятно, связана высокая степень пластичности головного мозга по компенсации нарушенных функций мозжечка. Известны случаи врожденного отсутствия мозжечка или медленного разрушения его опухолью, при которых у человека не определялись симптомы нарушения движения.
Б. Афферентная функция мозжечка. Выше изложены в основном афферентные связи мозжечка с проприорецепторами, кожными и вестибулярными рецепторами. Известно мнение Ч. Шеррингтона о мозжечке как «главном ганглии» проприоцептивной чувствительности. Однако, как показали электрофизиологические исследования, изменения активности клеток Пуркинье возникали при стимуляции практически всех рецепторов — зрительных, слуховых, висцеральных и др. Вместе с тем исследования Л.А. Орбели людей с ранениями мозжечка показали изменения пороговых величин различных видов чувствительности. Эти данные свидетельствуют о сложных двусторонних связях мозжечка и сенсорных систем. В реализации влияния мозжечка на афферентные системы организма, без сомнения, большую роль играют проекции ядер мозжечка на специфические и неспецифические ядра таламуса как главного центра переключения в сенсорных системах. Можно предположить, что механизмы влияния мозжечка на сенсорные функции связаны с его влиянием на эфферентный контроль активности рецепторного аппарата и центров переключения в сенсорных системах.
В последнее время исследуется участие мозжечка в процессах мышления. Обнаружено, что при мыслительных операциях (например, арифметическом счете) увеличивается локальный кровоток не только в ассоциативной коре, но и в полушариях мозжечка. Локальная биоэлектрическая активность в полушариях мозжечка была отмечена в процессах слышания и видения. Предполагается, что мысль и движение контролируются одними и теми же нейронными системами [Ito М., 1993].
В. Роль мозжечка в регуляции вегетативных функций изложена в разделе 9.9.
8.5. ЛИМБИЧЕСКАЯ СИСТЕМА
Термин «лимбическая система» (limbus — край) был предложен П. Мак-Л ином (1952) и отражает особенность расположения ее в ви-
де кольца на границе новой коры, отделяющей ее от ствола мозга. Под лимбической системой понимают функциональное объединение различных структур конечного, промежуточного и среднего мозга, обеспечивающее эмоционально-мотивационные компоненты поведения и интеграцию висцеральных функций организма. В эволюционном аспекте лимбическая система сформировалась в процессе усложнения форм поведения организма, перехода от жестких, генетически запрограммированных форм поведения к пластичным, основанным на обучении и памяти.
8.5.1. СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ ЛИМБИЧЕСКОЙ СИСТЕМЫ
В более узком понимании в лимбическую систему включают образования древней коры (обонятельная луковица и бугорок, периа-мигдалярная и препириформная кора), старой коры (гиппокамп, зубчатая и поясная извилины), подкорковые ядра (миндалина, ядра перегородки). По отношению к гипоталамусу и ретикулярной формации ствола этот комплекс рассматривается как более высокий уровень интеграции вегетативных функций. В настоящее время преобладает понимание лимбической системы в более широком плане; кроме вышеназванных структур, в нее также включают зоны новой коры лобной и височной долей, гипоталамус и РФ среднего мозга.
Важнейшие афферентные входы в лимбическую систему осуществляются от различных областей головного мозга, а также через гипоталамус от РФ ствола, которая считается главным источником ее возбуждения. Важным стимулирующим афферентным входом являются также импульсы от обонятельных рецепторов, поступающие по волокнам обонятельного нерва (п. olfactorii, I) в структуры лимбической системы — обонятельные луковицу и бугорок, параобонятельную зону Брока, препириформную кору, миндалину. В лимбической системе, таким образом, находится корковый отдел древнего — обонятельного — анализатора. Главные эфферентные выходы из лимбической системы осуществляются через гипоталамус (особенно его мамиллярные тела) на ниже лежащие вегетативные и соматические центры ствола и спинного мозга. Другой эфферентный выход проводит возбуждение из лимбической системы в новую кору (преимущественно ассоциативную). Через нее лимбическая система
135
Функционально и топографически латеральный ретикулоспинальный тракт сходен с руб-роспинальным и кортико-спинальным трактами; они образуют нисходящую латеральную флексорную (сгибательную) систему. Вместе с тем раздражение нейронов центральной части РГЯ в эксперименте вызывает подавление всех спинальных рефлексов (сги-, бательных и разгибательных) за счет активации тормозных интернейронов в соответствующих сегментах спинного мозга. По этому же механизму осуществляется открытое И.М. Сеченовым (1862) центральное торможение рефлексов спинного мозга.
От каудальных и оральных ретикулярных ядер моста идет неперекрещенный медиальный ретикулоспинальный тракт с быстропроводя-щими (до 120 м/с) волокнами, оканчивающимися на интернейронах спинного мозга. Через них осуществляется стимуляция а- и у-нейро-нов мышц—разгибателей осевой мускулатуры тела (туловища и проксимальных отделов конечностей) и через тормозные нейроны тормозятся центры сгибателей. В функциональном и топографическом отношении этот тракт сходен с вестибулоспинальными трактами; они составляют медиальную нисходящую экстензорную (разгибательную) систему.
Ретикулярная формация ствола мозга через у-мотонейроны спинальных и бульбар-ных моторных центров осуществляет центральный контроль активности мышечных рецепторов [Гранит Р., 1955], при этом тормозные влияния на них легче осуществляются при стимуляции РФ на уровне продолговатого мозга, а облегчающие влияния — на уров-I не моста и среднего мозга.
Восходящие влияния РФ па большой мозг преимущественно активирующие. Импульсы ретикулярных нейронов продолговатого мозга (гигантоклеточное, латеральное и вент-ральное ретикулярные ядра), моста (особенно каудальное ретикулярное ядро) и среднего мозга поступают к неспецифическим ядрам тшмуса и после переключения в них проецируются в различные области коры. Кроме таламуса, восходящие влияния следуют также в задний гипоталамус, полосатое тело, прозрачную перегородку.
Изучение активирующего влияния восходящей РФ началось с опыта по перерезке ствола мозга между верхними и нижними буграми четверохолмия — изоляции переднего мозга [Бремер Ф., 1935]. У экспериментального животного не нарушалось поступление в кору большого мозга возбуждения по важнейшим сенсорным системам — зрительной и обонятельной. Однако животное вело
себя как спящее: у него был нарушен контакт с внешним миром, оно не реагировало на световые и обонятельные раздражители (спящий мозг по Бремеру). На электроэнцефалограмме (ЭЭГ) у таких животных преобладали медленноволновые регулярные ритмы. У человека аналогичные ритмы возникают при спокойном бодрствовании и в дремотном состоянии. Такое же состояние головного мозга остается, если при перерезке ствола мозга сохранить основные афферентные пути, составляющие медиальную петлю (например, от лица по тройничному нерву) и повредить только восходящие пути РФ.
Прямое доказательство активирующего влияния РФ по восходящим путям на состояние мозга было получено Г. Мегуном и Дж, Моруцци (1949) в хронических опытах с раздражением РФ через погружные электроды у сонных животных. Стимуляция РФ вызывала пробуждение животного. На ЭЭГ медленные ритмы сменялись высокочастотными ритмами (реакция десинхронизации), свидетельствующими об активированном состоянии коры головного мозга. На основании полученных данных сложилось представление о том, что важнейшей функцией восходящей РФ является регуляция цикла сон/бодрствование и уровня сознания.
Тормозное влияние РФ на большой мозг изучено значительно хуже. Работами В. Гесса (1929), Дж. Моруцци (1941) было показано, что раздражением некоторых точек РФ ствола мозга можно перевести животное из бодрствующего состояния в сонное, при этом на электроэнцефалограмме возникает реакция синхронизации ритмов ЭЭГ.
Вегетативные функции РФ осуществляются через ее влияния на вегетативные центры ствола и спинного мозга. Ретикулярная формация входит в состав жизненно важных центров продолговатого мозга — сердечно-сосудистого и дыхательного.
8.2.3. СТВОЛОВЫЕ РЕФЛЕКСЫ ПОДДЕРЖАНИЯ ПОЗЫ
Рефлексы поддержания позы осуществляются с рецепторов вестибулярного аппарата, проприорецепторов шеи, рецепторов сетчатки и тактильных рецепторов. Эфферентные влияния из моторных центров ствола распространяются на мышцы конечностей и туловища по четырем нисходящим трактам (см. рис. 8.5): вестибулоспинальному, руброспи-нальному, латеральному и медиальному рети-кулоспинальному.
125
Вестибулоспинальный тракт начинается преимущественно от вестибулярного латерального ядра (Дейтерса), а также от верхнего и медиального вестибулярных ядер, получающих афферентную импульсацию от рецепторов отолитового аппарата и ампул полукружных каналов. Его волокна оказывают возбуждающее действие на спинальные а-и у-мотонейроны мышц—разгибателей конечностей, туловища, шеи, а также реци-прокное тормозящее влияние на мотонейроны сгибателей. Аналогично на мышечный тонус влияет описанный выше медиальный ретикулоспинальный тракт. Вместе они составляют 'медиальную нисходящую разгиба-тельную систему.
Противоположное действие на мышечный тонус оказывает руброспинальный тракт (Монакова), который начинается от красного ядра покрышки среднего мозга и идет в спинной мозг, возбуждая а- и у-нейроны сгибателей и реципрокно тормозя мотонейроны разгибателей. Аналогично на мышечный тонус действуют латеральный ретикулоспинальный и пирамидный кортико-спиналь-ный тракты. Вместе они составляют латеральную нисходящую сгибательную систему. Таким образом, в стволе мозга имеется четыре главных парных моторных центра и тракта, регулирующих тонус мышц туловища и конечностей (см. рис. 8.5).
Взаимоотношения между этими центрами показаны в опытах с перерезкой ствола мозга между продолговатым и средним мозгом [Шеррингтон Ч., 1896). У таких животных возникает резкое преобладание тонуса мышц-разгибателей над тонусом сгибателей конечностей, туловища, шеи, хвоста (деце-ребрационная ригидность). Причина ее связана с тем, что влияние структур медиальной нисходящей разгибательной системы не уравновешивается влияниями латеральной нисходящей сгибательной системы, в частности красного ядра, оказывающегося выше перерезки. Частный механизм децеребраци-онной ригидности связан с активацией как у-мотонейронов, так и а-мотонейронов мышц-разгибателей.
* Через стволовые моторные центры и их тракты осуществляется регуляция установки тела в пространстве, направленной на сохранение нормальной позы тела и равновесия. Эти рефлексы изучены голландским физиологом Р. Магнусом (1924), который разделил их на две группы: статические и статокинети-ческие.
А. Статические рефлексы обусловливают положение тела и его равновесие в покое.
Магнус разделил их на рефлексы позы (положения тела), выпрямительные рефлексы и рефлексы компенсаторного положения глаз.
1. Рефлексы позы возникают при измене ниях положения головы (сдвиг центра равно весия) и направлены на создание удобной позы. Афферентные импульсы поступают с рецепторов отолитового аппарата и проприо- рецепторов мышц шеи. Шейные рефлексы описаны выше. Для выявления вестибуляр ных рефлексов на шею животного, располо женную на одной линии с туловищем, накла дывают гипсовую повязку, чтобы исключить шейные рефлексы. Нейтральное положение головы горизонтальное, при этом импульса- ция с рецепторов лабиринта минимальна. Для вызова вестибулярных рефлексов живот ное поворачивают через ось висков. Если жи вотное располагают спиной вниз, тонус мышц-разгибателей становится максималь ным, если спиной вверх — минимальным. При вертикальной позе человека (голова вверх, шея наклонена несколько вперед) про исходит повышение тонуса разгибателей нижних конечностей и сгибателей верхних конечностей. При этом шейные и лабиринт ные рефлексы усиливают друг друга.
-
Выпрямительные рефлексы — более сложный вариант статических рефлексов. Благодаря этим рефлексам организм спосо бен принимать естественную позу при ее на рушении, например при положении живот ного на спине. Рефлексы осуществляются с рецепторов шейных мышц, лабиринта, кожи, сетчатки при обязательном участии среднего мозга. Важными компонентами этих рефлек сов являются шейный, вестибулярный и оп тический установочные рефлексы, а первой двигательной реакцией — восстановление нормального положения головы. Далее воз никает цепь рефлекторных реакций с пере распределением тонуса мышц конечностей и туловища — восстанавливается нормальная ориентация тела в пространстве.
-
Рефлексы компенсаторного положения глаз обеспечивают правильное видение при различных положениях головы относительно тела и в пространстве. Например, если на клонить голову к правому плечу, глазное яб локо поворачивается влево, как бы стремясь удержать первоначальное поле зрения.
Б. Статокинетические рефлексы возникают при ускорениях прямолинейного и вращательного движений организма. Сокращения мыщц при этом направлены на преодоление действующих на человека ускорений, сохранение нормальной позы, равновесия и ориентации в пространстве. Для их осущест-
126
вления необходимо сохранение функции моторных центров ствола мозга не ниже уровня среднего мозга. Эти рефлексы запускаются с рецепторов вестибулярного аппарата: с рецепторов отолитового аппарата возникают рефлексы прямолинейного ускорения, с рецепторов ампул полукружных каналов — рефлексы вращения.
Примером рефлекса прямолинейного ускорения является лифтный рефлекс. В начале быстрого подъема лифта (или остановки движущегося вниз лифта) в нижних конечностях повышается тонус разгибателей, что обеспечивает сохранение позы — преодоление сил гравитации, хотя наблюдается некоторое сгибание конечностей под влиянием сил гравитации. В начале опускания лифта (или остановки движущегося вверх лифта) динамика изменения мышечного тонуса сгибателей и разгибателей и положение конечностей противоположные, однако имеется и противоположная точка зрения, базирующаяся только на внешних признаках, — сгибание или разгибание конечностей.
Статокинетические рефлексы вращения включают мышцы тела и глазные мышцы. Движение глазных яблок (глазной нистагм) при этом способствует сохранению зрительной ориентации. Нистагм имеет две фазы. При ускорении вращения сначала происходит медленное отклонение глазных яблок в сторону, противоположную вращению. После этого они быстро отклоняются обратно (по ходу вращения). Медленное отклонение вызывается с рецепторов полукружных каналов, быстрый компонент связан с влиянием корковых центров (при наркозе он исчезает). Во врачебной практике направление нистагма обозначают по быстрому компоненту. При замедлении вращения, напротив, медленное движение глаз направлено в сторону вращения, а быстрое — против направления вращения. Движения глаз при нистагме (вправо-влево, вверх-вниз, круговые движения) зависят от возбуждения рецепторов соответственно горизонтального, сагиттального и фронтального полукружных каналов. Исследование нистагма (степень его выраженности, длительность) используется для оценки функционального состояния вестибулярного аппарата. Глазной нистагм может возникнуть без вращения, если происходит раздражение рецепторов полукружных каналов патологическим процессом. •
Кроме регуляции мышечного тонуса, позы и равновесия, структуры ствола головного мозга участвуют в контроле спинального автоматизма шагания и, следовательно, в
контроле локомоции (например, ходьбы). Важное значение в осуществлении этой функции придают клинообразному ядру в задненижнем отделе среднего мозга, который обозначают как локомоторную область среднего мозга (ЛОСМ), имеющую эфферентный выход к ядрам моста. У кошки с перерезкой ствола между средним и промежуточным мозгом разрушение ЛОСМ полностью прекращает способность к бегу и резко нарушает ходьбу. Без разрушения ЛОСМ эти функции восстанавливаются через несколько недель после перерезки. Для осуществления локомоции важна также область гипоталамуса под мамиллярными телами. Однако эти структуры составляют только часть системы управления локомоцией. У человека, например, она без участия корковых отделов невозможна.
Роль различных отделов ствола мозга в регуляции движения можно выявить в опытах на животных с перерезкой ствола или удалением вышележащих отделов мозга. В медицинской практике подобные ситуации возникают при торможении вышележащих отделов мозга в результате тяжелых патологических процессов (например, при коме). При перерезке в эксперименте между продолговатым и средним мозгом формируется бульварный организм. Наиболее яркой особенностью его двигательных функций является децеребра-ционная ригидность (см. выше). Такой организм имеет статические позные рефлексы (шейные, лабиринтные), но у него отсутствуют выпрямительные рефлексы, локомоция, произвольные движения.
Двигательные функции мезэнцефалическо-го организма (перерезка между средним и промежуточным мозгом) характеризуются нормальным мышечным тонусом, наличием статических и статокинетических рефлексов (сохранение равновесия, возврата в нормальную позу), способностью к координированным движениям (например, ходьбе, бегу) у животных, но при этом нарушаются сложные целенаправленные движения.
8.2.4. ФУНКЦИИ ГОЛУБОГО ПЯТНА
Голубое пятно имеется только у млекопитающих, расположено в каудальном отделе среднего мозга (около мезэнцефалического ядра V нерва), является основным норадренерги-ческим образованием среднего мозга, передающим импульсные влияния через медиатор норадреналин. Аксоны нейронов голубого пятна (ГП) связаны с корой большого мозга,
127
большинством ядер ствола мозга, промежуточного мозга, моторными центрами спинного мозга. Афферентные входы ГП получает от ядер тройничного нерва, ядра одиночного пути, гипоталамуса, РФ ствола, черного вещества.
Моторные функции ГП. Аксоны его нейронов идут в переднем и боковом канатиках к а-мотонейронам передних рогов спинного мозга. Предполагают, что норадреналин является тормозным медиатором для а-мото-нейронов. Импульсная активность нейронов ГП повышена в фазе быстрого сна, что выключает мышечный тонус и физические движения мускулатуры туловища и конечностей. При одностороннем разрушении ГП возникают вращательные движения в сторону, противоположную разрушению, что, как предполагают, связано с нарушением функциональных связей ГП с базальными ганглиями и черным веществом.
Гомеостатическая функция ГП. В функциональном плане ГП тесно связано с чувствительными ядрами тройничного, языкогло-точного и блуждающего нервов; вместе они составляют основную мозговую структуру, обеспечивающую гомеостазис организма. Эта способность связана с тем, что ГП, с одной стороны, способно реагировать на изменение газового состава крови и ликвора, а с другой — имеет многочисленные эфферентные выходы на гипоталамус, РФ и вегетативные центры, обеспечивающие нейрогуморальную регуляцию состава внутренней среды организма.
8.2.5. ПРОВОДНИКОВАЯ ФУНКЦИЯ СТВОЛА МОЗГА
Выполняется восходящими и нисходящими путями.
А. Восходящие пути являются частью проводникового отдела анализаторов, передающих информацию от рецепторов в проекционные зоны коры. В стволе мозга выделяют две восходящие системы — специфическую и неспецифическую.
1. Специфическую восходящую систему составляет лемнискоталамический путь, в котором выделяют медиальную и латеральную петли. Медиальная петля образуется преимущественно из аксонов нейронов тонкого (Голля) и клиновидного (Бурдаха) ядер, которые проводят от рецепторов конечностей, туловища и шеи проприоцептивную, тактильную и висцеральную чувствительность. В состав медиальной петли включают также во-
локна вентрального спиноталамического тракта, передающие тактильную чувствительность, и часть волокон тригеминального тракта (от ядер V нерва), проводящих от головы проприоцептивную и тактильную чувствительность. К медиальной петле на подходе к таламусу присоединяется часть волокон от ядра одиночного пути, проводящих вкусовую и висцеральную чувствительность, поступающую по волокнам языкоглоточного, блуждающего и лицевого нервов, а также небольшая часть волокон латерального спиноталамического тракта, проводящих острую локализованную боль. Волокна медиальной петли переключаются в вентральных задних специфических ядрах таламуса.
Латеральная петля (син. слуховая петля) составлена аксонами нейронов ядер трапециевидного тела и верхней оливы моста, входящих в проводниковый отдел слухового анализатора. Латеральная петля переключается в медиальном коленчатом теле таламуса и нижних буграх четверохолмия.
К специфической проводящей системе относятся проводящие пути зрительного и вестибулярного анализаторов.
Функциональной особенностью специфической проводящей системы является высокая скорость проведения возбуждения, ее нейроны имеют небольшие рецептивные поля, они преимущественно мономодальны. В центрах переключения афферентных нейронов (в стволе, таламусе) имеется выраженная топографическая проекция периферии. В результате обеспечивается быстрая передача информации с тонким различением свойств раздражителя. При этом в корковом отделе соответствующего анализатора с коротким латентным периодом возникают вызванные потенциалы — первичные ответы.
2. Неспецифические (экстралемнисковые) восходящие пути — в основном это волокна латерального спиноталамического и спино-ретикулярного трактов, проводящих температурную и болевую чувствительность. Спи-норетикулярный тракт переключается на нейроны РФ ствола, и в составе ее восходящих волокон импульсация поступает в неспецифические (интраламинарные, ретикулярное) ядра таламуса и далее проецируется в различные области коры (особенно лобную орбитальную кору). Неспецифическая система получает коллатеральные волокна от специфической системы, что обеспечивает связь этих двух восходящих систем. Функциональной особенностью неспецифической системы является относительно медленное проведение информации. Рецептивные поля ней-
128
ронов большие, нейроны полимодальные, связанные с несколькими видами чувствительности, топография проекции периферии в центрах не выражена. При проведении возбуждения через эту систему на ЭЭГ фиксируются с большим латентным периодом вызванные потенциалы —вторичные ответы.
Кроме проведения возбуждения по специфической и неспецифической восходящим системам, часть сенсорной импульсации поступает в мозжечок. После соответствующей обработки она проецируется в центры тала-муса и ствола мозга. Через ствол мозга в мозжечок проходят дорсальный спинно-мозжеч-ковый тракт Флексига, проводящий импуль-сацию от рецепторов мышц и связок, и вентральный спинно-мозжечковый тракт Говер-са — от рецепторов сухожилий, кожи и внутренних органов, а также вестибуломозжечко-вый тракт, несущий информацию от вестибулярных рецепторов. Из коры мозжечка через его зубчатые и промежуточные ядра обработанная проприоцептивная информация передается по церебеллоталамическому тракту в вентральные (переднее и латеральное) ядра таламуса, далее она проецируется в сомато-сенсорную, моторную и премоторные зоны коры.
Б. Нисходящие проводниковые пути ствола мозга в функциональном отношении можно объединить в несколько групп. Двигательные пирамидные пути, начинающиеся от клеток Беца коры прецентральной извилины, ин-нервируют мотонейроны передних рогов спинного мозга (кортико-спинальные пути), мотонейроны двигательных ядер черепных нервов (кортико-бульбарный путь), обеспечивая произвольные сокращения мышц конечностей, туловища, шеи и головы.
Моторные центры ствола мозга и их пути являются важнейшим компонентом экстрапирамидной системы, основной функцией которой является регуляция мышечного тонуса, позы и равновесия. К этой системе на уровне ствола мозга относятся кортико-красноядер-ный и кортико-ретикулярный тракты, оканчивающиеся на моторных центрах ствола, от которых идут рассмотренные выше рубро-, ретикуло- и вестибулоспинальный пути.
В стволе мозга проходят нисходящие пути, обеспечивающие двигательные функции мозжечка. К ним относится кортико-мостомоз-жечковый путь, по которому в мозжечок поступает импульсация от двигательной и других областей коры. Обработанная в коре мозжечка и его ядрах информация поступает на моторные ядра ствола (красное, вестибулярные, ретикулярные).
Через ствол мозга проходит начинающийся в четверохолмии тектоспинальный тракт, который обеспечивает двигательные компоненты ориентировочных зрительных и слуховых рефлексов.
8.3. ПРОМЕЖУТОЧНЫЙ МОЗГ
Промежуточный мозг (diencephalon) расположен между средним и конечным мозгом, вокруг III желудочка мозга. Он состоит из та-ламической области и гипоталамуса. Талами-ческая область включает в себя таламус, ме-таталамус (коленчатые тела) и эпиталамус (эпифиз).
8.3.1 ТАЛАМУС
Таламус (зрительный бугор) представляет собой парный ядерный комплекс, занимающий преимущественно дорсальную часть промежуточного мозга. Таламус составляет основную массу (около 20 г) промежуточного мозга, наибольшее развитие имеет у человека. В таламусе выделяют до 40 парных ядер, которые в функциональном плане можно разделить на следующие три группы: релейные, ассоциативные и неспецифические. Все ядра таламуса в разной степени обладают тремя общими функциями — переключающей, интегративной и модулирующей.
А. Релейные ядра таламуса (син.: переклю-чательные, специфические) разделяют на сенсорные и несенсорные. К главным сенсорным релейным ядрам относят вентральные задние ядра — n.VP (вентральное заднелатеральное — п. VPL и вентральное заднемедиальное — n.VPM), вентральное промежуточное ядро (n.VIM), латеральное (n.GL) и медиальное (n.GM) коленчатые тела. Главной функцией этих ядер является переключение потоков афферентной импульсации в сенсорные зоны коры. Наряду с этим происходят перекодирование и обработка информации.
Вентральные задние ядра (син. вентроба-зальный комплекс) являются главным реле для переключения соматосенсорной афферентной системы, импульсы которой поступают по волокнам медиальной петли и примыкающим к ней волокнам других афферентных путей. В них переключаются тактильная, проприоцептивная, вкусовая, висцеральная, частично температурная и болевая чувствительность. В этих ядрах имеется топографическая проекция периферии; различные виды чувствительности от головы,
129
лица, языка проецируются в вентральные заднемедиальные ядра (n.VPM), а от туловища, шеи, конечностей — в вентральные зад-нелатеральные ядра (n.VPL). При этом язык, лицо имеют большую зону представительства. Вентральные задние ядра проецируются в соматосенсорную кору постцентральной извилины (поля 3, 1, 2), где формируются соответствующие ощущения. Электр о стимуляция вентральных задних ядер вызывает парестезии (ложные ощущения) в разных частях тела, иногда нарушение «схемы тела» (искаженное восприятие частей тела). Стереотак-сическое разрушение участков этих ядер используется для ликвидации тяжелых болевых синдромов, характеризующихся острой локализованной болью и фантомными болями.
Латеральное коленчатое тело является реле для переключения зрительной импульсации в затылочную кору (поле 17), где она используется для формирования зрительных ощущений. Кроме корковой проекции, часть зрительной импульсации направляется в верхние бугры четверохолмия. Эта информация используется для регуляции движения глаз, в зрительном ориентировочном рефлексе. Медиальное коленчатое тело является реле для переключения слуховой импульсации в височную кору задней части сильвиевой борозды (извилины Гешля, поля 41, 42).
Переключение в таламусе афферентной импульсации от вестибулярного аппарата точно не показано. Возможно, она переключается в вентральном промежуточном ядре (п.VIM) и проецируется в нижнюю часть постцентральной извилины (поле 3). Некоторые исследователи считают, что переключение происходит в медиальном коленчатом теле с дальнейшей проекцией в кору верхней и средней височной извилин (поля 21 и 22).
В релейных ядрах обнаружено несколько типов нейронов. Таламокортикальные (релейные) нейроны, имеющие длинный аксон, непосредственно обеспечивают переключение поступающей афферентной импульсации на нейроны сенсорной коры. Особенностью этой передачи является ее высокая точность с минимальным искажением входного сигнала. Регуляция передачи возбуждения через релейные ядра осуществляется с помощью тормозных и возбуждающих вставочных нейронов. Тормозные интернейроны возбуждаются как от таламокортикальных нейронов ядра, осуществляя возвратное торможение, так и от корковых нейронов, проецирующихся в данное ядро (корковый контроль проведения возбуждения). Возбуждающие интернейроны имеют фоновую ритмическую ак-
тивность, которая подавляется при активации тормозных интернейронов и активируется при выходе из тормозной паузы.
К несенсорным релейным ядрам таламуса относятся передние и вентральные ядра. Они переключают в кору несенсорную импульса-цию, поступающую в таламус из разных отделов головного мозга. В передние вентральное, медиальное и дорсальное ядра (п.А V, пАМ, n.AD) импульсация поступает в основном по мамиллоталамическому пути из мамилляр-ных тел гипоталамуса. Нейроны передних ядер проецируются в лимбическую кору (поля 23, 24, 29, 32). От нее аксонные связи идут к гиппокампу и опять к гипоталамусу, в результате чего образуется нейронный круг, реверберация возбуждения по которому участвует в формировании эмоций («эмоциональное кольцо Пейпеца»). В связи с этим передние ядра таламуса рассматриваются как часть лимбической системы и иногда обозначаются как «лимбические ядра таламуса».
Вентральные переднее (п. VA) и латеральное (п. VL) ядра участвуют в регуляции движения, выполняя таким образом моторную функцию. В них переключается импульсация от базальных ганглиев, зубчатого ядра мозжечка, красного ядра среднего мозга, импульсы после этого проецируются в моторную и пре-моторную кору (поля 4 и 6). Таким образом, через эти ядра таламуса происходит передача в моторную кору сложных двигательных программ, образованных в мозжечке и базальных ганглиях.
Наряду с корковыми проекциями релейных ядер каждое из них получает нисходящие корковые волокна из той же проекционной зоны, что создает структурную основу для взаиморегулирующих отношений между корой и таламусом.
Б. К ассоциативным ядрам таламуса относятся ядра подушки (п. pulvinares), медиодор-сальное ядро (n.MD) и латеральные ядра -дорсальное (n.LD) и заднее (n.LP). Волокна к этим ядрам приходят не от проводниковых путей анализаторов, а от других ядер таламуса. Эфферентные выходы от этих ядер направляются главным образом в ассоциативные поля коры. В свою очередь кора мозга посылает волокна к ассоциативным ядрам, регулируя их функцию. Главной функцией этих ядер является интегративная функция, которая выражается в объединении деятельности как таламических ядер, так и различных зон ассоциативной коры полушарий мозга. Электростимуляция ассоциативных ядер вызывает в коре мозга биоэлектрическую реакцию в виде вторичного ответа.
330
Подушка получает главные входы от коленчатых тел и неспецифических ядер тала-муса. Эфферентные пути от нее идут в височ-но-теменно-затылочные зоны коры, участвующие в гностических (узнавание предметов, явлений), речевых и зрительных функциях (например, в интеграции слова со зрительным образом), а также в восприятии «схемы тела». Электростимуляция подушки доминантного полушария приводит к речевым нарушениям типа аномии (нарушение называния предметов), разрушение подушки вызывает нарушение «схемы тела», устраняет тяжелые боли.
Медиодорсальное ядро получает входы от гипоталамуса, миндалины, гиппокампа, тала-мических ядер, центральнбго серого вещества ствола. Проекция этого ядра распространяется на ассоциативную лобную и лимби-ческую кору (поля 12, 18). Оно участвует в формировании эмоциональной и поведенческой двигательной активности, а также, возможно, в механизмах памяти. Разрушение этих ядер устраняет у больных страх, тревогу, напряженность, страдание от боли, но возникает лобный синдром: снижение инициативы, безразличие, гипокинезия.
Латеральные ядра получают зрительную и слуховую импульсацию от коленчатых тел и соматосенсорную импульсацию от вентрального ядра. Интегрированная сенсорная информация от этих источников далее проецируется в ассоциативную теменную кору и используется в ее функциях гнозиса, праксиса, формирования схемы тела.
В. Неспецифические ядра составляют эво-йюционно более древнюю часть таламуса, включающую интралолшнарную (внутриплас-тмчатую) ядерную группу: центральное срединное ядро (п.СМ), парафасцикулярное fn.Pf) и субпарафасцикулярное (n.sPf) ядра, центральное латеральное (n.Cl) и медиальное (п.Cm) ядра, парацентральное ядро (п.Рс). Кроме того, к неспецифическим ядрам относят ретикулярное ядро (n.R). Эти ядра содержат преимущественно мелкие многоотрост-чатые нейроны и функционально рассматриваются как производное ретикулярной формации ствола мозга.
Неспецифические ядра имеют многочисленные входы как от других ядер таламуса, так и внеталамические: по латеральному спиноталамическому, спиноретикуло-тала-мическому трактам, текто- и тегментотала-мическому трактам, проводящим преимущественно болевую и температурную чувствительность. В неспецифические ядра поступает непосредственно или через ретикуляр-
ную формацию также часть импульсации по коллатералям от всех специфических сенсорных систем. Кроме того, в неспецифические ядра поступает импульсация из моторных центров ствола (красное ядро, черное вещество), ядер мозжечка (шатра, проб-кообразного), от базальных ганглиев и гиппокампа, а также от коры мозга, особенно лобных долей. Неспецифические ядра имеют эфферентные выходы на другие тала-мические ядра, кору большого мозга как непосредственно, так и через ретикулярные ядра, а также нисходящие пути к другим структурам ствола мозга.
Благодаря этим связям неспецифические ядра таламуса выступают в роли интегрирующего посредника между стволом мозга и мозжечком, с одной стороны, и новой корой, лимбической системой и базальными ганглиями, с другой стороны, объединяя их в единый функциональный комплекс. На кору мозга неспецифический таламус оказывает преимущественно модулирующее влияние. Разрушение неспецифических ядер не вызывает грубых расстройств эмоций, восприятия, сна и бодрствования, образования условных рефлексов, а нарушает только тонкую регулировку поведения. В связи с этим модулирующее влияние неспецифических ядер таламуса, обеспечивающее «плавную настройку» высшей нервной деятельности, считается их главной функцией.
8.3.2. ГИПОТАЛАМУС
Гипоталамус является вентральной частью промежуточного мозга. Макроскопически он включает в себя преоптическую область и область перекреста зрительных нервов, серый бугор и воронку, сосцевидные тела. Микроскопически в гипоталамусе выделяют, по данным разных авторов, от 15 до 48 парных ядер, которые подразделяются на 3—5 групп. Многие авторы выделяют в гипоталамусе 4 основные области, включающие в себя несколько ядер (по О.Г.Баклаваджану, 1981):
-
преоптическая область — медиальное и латеральное преоптические ядра;
-
передняя область — супрахиазматическое, супраоптическое, паравентрикулярное и переднее гипоталамическое ядра;
-
средняя (или туберальная) область —дор- сомедиальное, вентромедиальное, аркуат- ное (инфундибулярное) и латеральное ги- поталамические ядра;
-
задняя область — супрамамиллярное, пре- мамиллярное, латеральное и медиальное
131
мамиллярные ядра, заднее гипоталамичес-кое и перифорниатное ядра, субталами-ческое ядро Луиса.
Важной физиологической особенностью гипоталамуса является высокая проницаемость его сосудов для различных веществ, в том числе и для крупных полипептидов. Это обусловливает большую чувствительность гипоталамуса к сдвигам во внутренней среде организма и способность реагировать на колебания концентрации гуморальных веществ. В гипоталамусе по сравнению с другими структурами головного мозга имеются самая мощная сеть капилляров (1100—2600 капилляров/мм2) и самый большой уровень локального кровотока.
Ядра гипоталамуса образуют многочисленные связи друг с другом (ассоциативные), с парными одноименными ядрами противоположной стороны (комиссуральные), а также с выше- и нижележащими структурами ЦНС (проекционные). Главные афферентные пути гипоталамуса идут от лимбической системы, коры больших полушарий, базаль-ных ганглиев и ретикулярной формации ствола. Основные эфферентные пути гипоталамуса идут в ствол мозга — его ретикулярную формацию, моторные и вегетативные центры, в вегетативные центры спинного мозга, от мамиллярных тел к передним ядрам таламуса и далее в лимбическую систему, от супраоптического и паравентрикулярного ядер к нейрогипофизу, от вентромедиального и инфундибулярного ядер к аденогипофизу, а также имеются эфферентные выходы к лобной коре и полосатому телу.
Гипоталамус является многофункциональной системой, обладающей широкими регулирующими и интегрирующими влияниями. Однако важнейшие функции гипоталамуса трудно соотнести с его отдельными ядрами. Как правило, отдельно взятое ядро имеет несколько функций, а отдельно взятая функция локализуется в нескольких ядрах. В связи с этим физиология гипоталамуса рассматривается обычно в аспекте функциональной специфики его различных областей и зон.
Гипоталамус является важнейшим центром интеграции вегетативных функций, регуляции эндокринной системы, теплового баланса организма, цикла «бодрствование — сон» и других биоритмов; велика его роль в организации поведения (пищевого, полового, агрессивно-оборонительного), направленного на реализацию биологических потребностей.
8.4. Мозжечок
Мозжечок — отдел головного мозга, образующий вместе с мостом задний мозг. Составляя 10 % массы головного мозга, мозжечок включает в себя более половины" всех нейронов ЦНС. Это свидетельствует о больших возможностях обработки информации и соответствует главной функции мозжечка как органа координации и контроля сложных и автоматизированных движений. В осуществлении этой функции важную роль играют обширные связи мозжечка с другими отделами ЦНС и рецепторным аппаратом.
8.4.1. ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ И СВЯЗИ МОЗЖЕЧКА
Выделяют три структуры мозжечка, отражающие эволюцию его функций (по Дж. Джан-сенду, А. Бродалу, 1954).
Црёвжй мозжечок (архицеребеллум) состоит из клочка и узелка (флоккулонодулярная доля) и нижней части червя. Он гомологичен мозжечку круглоротых, передвигающихся в воде с помощью змеевидных движений тела.
Старый мозжечок (палеоцеребеллум) включает в себя верхнюю часть червя и пара-флоккулярный отдел. Он гомологичен мозжечку рыб, передвигающихся с помощью плавников.
Новый мозжечок (неоцеребеллум) состоит из полушарий и появляется у животных, передвигающихся с помощью конечностей.
М^жнейронные связи в коре мозжечка, его афферентные входы и эфферентные выходы многочисленны. Грушевидные нейроны (клетки Пуркинье), "'образующие средний (ганглиозныи) слой коры, являются главной функциональной единицей. Ее структурной основой являются многочисленные ветвящиеся дендриты, на которых в одной клетке может быть до 100 тыс. синапсов. Количество ^клетоК-ГТуркинь^ v человека, по разным источникам, — от 7 до 30 млн. Они являются единственными эфферентными нейронами коры мозжечка и непосредственно связывают ее с внутримозжечковыми и вестибулярными ядрами. В связи с этим функциональное влияние мозжечка существенным образом зависит от активности клеток Пуркинье, что в свою очередь связано с афферентными входами этих клеток. Главными мозжечковыми афферентами являются системы лиано-видных и моховидных волокон. В последнее время изучается и третья система — моноа-минергические афференты.
132

А — круг Пейпеца; Б — круг черед миндалину; МТ — мамиллярные тела.
включается в регуляцию высших психических функций.
Характерным свойством лимбической системы является наличие хорошо выраженных кольцевых нейронных связей, объединяющих различные ее структуры (рис. 8.6). Эти связи дают возможность длительной циркуляции (реверберации) возбуждения, которая является механизмом его пролонгирования, повышения проводимости синапсов и формирования памяти. Реверберация возбуждения создает условия для сохранения единого функционального состояния структур замкнутого круга и навязывания этого состояния другим структурам мозга. Важнейшим циклическим образованием лимбической системы является лимбический круг Пейпеца (1937), идущий от гиппокампа через свод к мамиллярным телам, от них через мамилло-таламический пучок — к передним ядрам та-ламуса, от него в поясную извилину и от нее через парагиппокамповую извилину — обратно к гиппокампу. Этот круг играет большую роль в формировании эмоций, научения и памяти. Другой лимбический круг (от миндалины через терминальную полоску к мамиллярным телам гипоталамуса, от них — к лимбической области среднего мозга и обратно к миндалинам) имеет важное значение для формирования агрессивно-оборонительных, пищевых и сексуальных реакций.
8.5.2. ФУНКЦИИ ЛИМБИЧЕСКОЙ СИСТЕМЫ
Получая информацию о внешней и внутренней среде организма, лимбическая система после сравнения и обработки этой информа-
ции запускает через эфферентные выходы вегетативные, соматические и поведенческие реакции, обеспечивающие приспособление организма к внешней среде и сохранение внутренней среды на определенном уровне.
А. Регуляция висцеральных функций. Лим-бическую систему иногда называют «висцеральным мозгом». Эта функция осуществляется преимущественно через деятельность гипоталамуса, который является диэнце-фалическим звеном лимбической системы. О тесных эфферентных связях лимбической системы (через гипоталамус) с внутренними органами свидетельствуют разнообразные изменения их функций при раздражении лим-бических структур, особенно миндалины. При этом эффекты имеют различный знак в виде активации или угнетения висцеральных функций: происходит повышение или понижение частоты сердечных сокращений, моторики и секреции желудка и кишечника, секреции различных гормонов аденогипофи-зом (особенно АКЛТ и гонадотропинов).
Б. Формирование эмоций — переживаний, в которых отражается субъективное отношение человека к предметам внешнего мира и результатам собственной деятельности. В свою очередь эмоции являются субъективным компонентом мотиваций — состояний, запускающих и реализующих поведение, направленное на удовлетворение возникших потребностей. Через механизм эмоций лимбическая система улучшает приспособление организма к изменяющимся условиям среды.
В иерархии мозговых структур гипоталамус является критической зоной для возникновения эмоций. В структуре эмоций выделяют собственно эмоциональные пережива-
136
ния и его периферические (вегетативные и соматические) проявления. Эти компоненты эмоций могут иметь относительную самостоятельность в проявлениях: выраженные субъективные переживания могут сопровождаться небольшими периферическими проявлениями, и, напротив, при псевдоэмоциях яркие периферические проявления (например, мимические и вегетативные реакции при плаче или смехе) могут протекать без существенных субъективных переживаний. Гипоталамус в этом плане представляется структурой, ответственной преимущественно за вегетативные проявления эмоций.
Кроме гипоталамуса, к структурам лимби-ческой системы, наиболее тесно связанным с эмоциями, принадлежат поясная извилина и миндалина. Электрическая стимуляция миндалины у человека вызывает преимущественно отрицательные эмоции — страх, гнев, ярость. Напротив, двустороннее удаление миндалин в эксперименте на обезьянах резко снижает их агрессивность, повышает тревожность, неуверенность в себе. У таких животных нарушается способность оценивать информацию (особенно зрительную и слуховую), поступающую из окружающей среды, и связывать эту информацию со своим эмоциональным состоянием. В результате нарушается нормальное взаимодействие организма с окружающей средой, в том числе зоосоци-альные отношения с другими особями в группе. Наряду с этим миндалина участвует в процессе сравнения конкурирующих мотива-ций, выделения из них доминирующей мотивации и, следовательно, влияет на выбор поведения. Поясная извилина, имеющая многочисленные связи как с новой корой, так и со стволовыми центрами, выполняет, по мнению некоторых авторов, роль главного интегратора различных систем мозга, участвующих в формировании эмоций.
Важную роль в регуляции эмоций играет вентральная лобная кора, имеющая хорошо выраженные связи с миндалиной. Поражение лобной коры вызывает резкие нарушения эмоций у человека. Характерным является возникновение эмоциональной тупости. В первую очередь нарушаются эмоции, связанные с социальными отношениями людей, творчеством, и растормаживаются эмоции, связанные с удовлетворением биологических потребностей.
В. Участие в формировании памяти и обучения. Эта функция преимущественно связана с основным лимбическим кругом Пейпеца (см. рис. 8.6). Вместе с тем в одной из форм обучения (однократном обучении) важную
роль играет миндалина благодаря ее свойству индуцировать сильные отрицательные эмоции, что способствует быстрому и прочному формированию временной связи.
Среди структур лимбической системы, ответственных за память и обучение, весьма важную роль играют гиппокамп и связанные с ним задние зоны лобной коры. Их деятельность необходима для консолидации памяти — перехода кратковременной памяти в долговременную. Повреждение гиппокампа у человека вызывает резкое нарушение усвоения новой информации, образования долговременной памяти.
Электрофизиологической особенностью гиппокампа является то, что в ответ на сенсорное раздражение, стимуляцию ретикулярной формации и заднего гипоталамуса в гип-покампе развивается синхронизация электрической активности в виде тета-ритма (4— 7 Гц). При этом в новой коре, напротив, возникает десинхронизация в виде бета-ритма (14—30 Гц). Пейсмекером тета-ритма считают перегородку — ее медиальное ядро. Вопрос о поведенческих проявлениях гиппо-кампального тета-ритма до сих пор не решен. Его считают индикатором участия гиппокампа в ориентировочных рефлексах, реакциях внимания, настороженности, развитии эмоционального напряжения (ритм напряжения, по П.К. Анохину). Однако большинство исследователей считают, что он связан с обработкой мозгом информации и организацией памяти. Другой электрофизиологической особенностью гиппокампа является его уникальная способность на стимуляцию отвечать длительной (в течение часов, дней и даже недель) посттетанической потенциацией, которая приводит к облегчению синаптической передачи и является основой формирования памяти. Ультраструктурным проявлением участия гиппокампа в процессах памяти является увеличение числа шипиков на дендри-тах его пирамидных нейронов, что свидетельствует об усилении синаптической передачи возбуждающих и тормозных влияний.