

Коллоквиумы / Коллоквиум_возбудимые_ткани
.pdfКонтрольные вопросы по теме «Физиология возбудимых тканей»:
1. Каково строение клеточной мембраны?
Плазматическая мембрана – обязательный компонент любой клетки. По химическому составу мембрана представляет из себя белковолипидное образование с приблизительно равным весовым соотношением данных компонентов.
Структурную основу мембран составляют молекулы липидов, в непрерывный бислой которых включены отдельные белковые молекулы.
Основу билипидного слоя составляют фосфолипиды. В состав липидного слоя эукариот входят гликолипиды и стерины.
Основную массу липидов в мембране эукариотических клеток составляют фосфолипиды, которые составляют 80% всех липидов. Кроме липидов и белков в мембране присутствуют углеводы.
Мембранные белки связаны с липидным бислоем различными способами. Мембранные белки представлены 3-я разновидностями:
1.Периферические.
2.Интегральные (трансмембранные).
3.Полуинтегральные.
•Периферические: располагаются на поверхности билипидного слоя и связаны с интегральными белками и полярными головками липидных молекул электростатическими, водородными связями, солевыми мостиками; периферические белки никогда не образуют сплошного слоя; они, в основном, растворимы в воде, легко отделяются от мембраны без ее разрушения; некоторые периферические белки обеспечивают связь между мембранами и цитоскелетом.
•Интегральные белки мембран нерастворимы в воде; один из доменов интегрального белка встроен в гидрофобную часть бислоя мембраны, поэтому интегральный белок, не может быть удален из мембраны без ее разрушения. Интегральные белки полностью располагаются в билипидном слое.
•Полуинтегральные белки погружены в билипидный слой частично. Весь набор белковых молекул распределен в мембране мозаично и легко перемещается в ее плоскости с участием элементов цитоскелета, которые образуют связи с интегральными белками.
2. В чем заключаются функции клеточных мембран?
Функции мембран:
•Барьерные - обеспечивает регулируемый, избирательный, пассивный и активный обмен веществ с окружающей средой
•Механические – связана с цито скелетом и поддерживает форму клетки
•Транспортные - транспорт через мембраны обеспечивает: доставку питательных веществ, удаление конечных продуктов обмена, секрецию различных веществ, создание ионных градиентов, поддержание в клетке оптимального pH и концентрации ионов, которые нужны для работы клеточных ферментов.
•Осмотические – поддержание тургорного давление клетки
•Электрические – участие в передаче зарядов (например, межклеточные контакты)
•Осуществление генерации и проведения биопотенциалов. С помощью мембраны в клетке поддерживается постоянная концентрация ионов: концентрация иона К+ внутри клетки значительно выше, чем снаружи, а
концентрация Na+ значительно ниже, что очень важно, так как это обеспечивает поддержание разности потенциалов на мембране и генерацию нервного импульса.
•Энергетические - при фотосинтезе в хлоропластах и клеточном дыхании в митохондриях в их мембранах действуют системы переноса энергии, в которых также участвуют белки.
•Рецепторные и другие - некоторые белки, находящиеся в мембране, являются рецепторами (молекулами, при помощи которых клетка воспринимает те или иные сигналы).
3. Какие существуют виды транспорта веществ через мембрану?
Пассивный (идущий по градиенту концентрации и/или заряда без затрат энергии)
1) Простая диффузия – прохождение веществ непосредственно между фосфолипидами
МОГУТ
мелкие гидрофобные (кислород)
средние гидрофобные (стероиды)
мелкие полярные (вода)
НЕ МОГУТ
средние полярные (глюкоза)
мелкие заряженные (протоны)
средние заряженные (аминокислоты)
крупные молекулы (белки)
2)Облегченная диффузия (вещество связывается с переносчиком и такой комплекс диффундирует через мембрану, например – транспорт глюкозы)
3)Канальный транспорт (через белки-каналы, например, калиевые и натриевые каналы, участвующие в создании потенциала действия)
Активный (идущий против градиента концентрации и/или заряда с затратами энергии АТФ)

1)Первичноактивные транспортные белки (Na-К АТФ-аза с затратой энергии АТФ выкачивает из клетки ионы натрия и закачивает ионы калия)
2)Вторичноактивные транспортные белки (работают за счет градиента другого вещества, который создается за счет первичноактивного транспорта, например, аминокислоты в кишечнике всасываются за счет симпорта с натрием).
3)Векторный транспорт (вещество, зашедшее пассивно, внутри клетки с затратой энергии модифицируется, так что его концентрация внутри остается низкой, например, глюкоза превращается в гликоген)
4)Эндоцитоз (фаго- и пиноцитоз) происходит за счет активности цитоскелета, лежащего под мембраной.
4. Какие ткани относятся к возбудимым?
Возбудимые ткани — специально приспособленные к осуществлению быстрых ответных реакций на действие раздражителя нервная ткань, железистая ткань и мышечная ткань.
5.Какими свойствами обладают возбудимые ткани?
1)Раздpажимость
2)Возбудимость
3)Пpоводимость
4)Память
Раздpажимость - это способность клетки, ткани или оpгана воспpинимать действие pаздpажителя изменением метаболизма, стpуктуpы и функций.
Раздpажимость является унивеpсальным свойством всего живого и является основой пpиспособительных pеакций живого оpганизма к постоянно меняющимся условиям внешней и внутpенней сpеды.
Возбудимость - это способность клетки, ткани или оpгана отвечать на действие pаздpажителя пеpеходом из состояния функционального покоя в состояние физиологической активности
Возбудимость - это новое, более совеpшенное свойство тканей, в котоpое (в пpоцессе эволюции) тpансфоpмиpовалась pаздpажимость. Разные ткани обладают pазличной возбудимостью: неpвная> мышечная> железистая
Меpой возбудимость является поpог pаздpажения
Поpог pаздpажения - это минимальная сила pаздpажителя, способная вызвать pаспpостpоняющееся возбуждение
Возбудимость и поpог pаздpажения находятся в обpатной зависимости (чем
>возбудимость, тем < поpог pаздpажения)
6.Дайте определение понятия «потенциал покоя».
Потенциал покоя — это разность электрических потенциалов между наружной и внутренней средой клетки.
7. Опишите механизмы, которые участвуют в поддержании мембранного потенциала покоя.
Механизм формирования потенциалов покоя. Непосредственная причина потенциала покоя — это неодинаковая концентрация анионов и катионов внутри и вне клетки.
Во-первых, такое расположение ионов обосновано разницей проницаемости.
Во-вторых, ионов калия выходит из клетки значительно больше, чем натрия. Создание потенциала покоя обеспечивается двумя основными процессами — неравномерным распределением неорганических ионов между внутри- и внеклеточным пространством и неодинаковой проницаемостью для них клеточной мембраны. Анализ химического состав вне- и внутриклеточной жидкости свидетельствует о крайне неравномерном распределении ионов.
Всостоянии покоя внутри клетки много анионов органических кислот и ионов К+, концентрация которых в 30 раз больше, чем снаружи; ионов Na+, наоборот, снаружи клетки в 10 раз больше, чем внутри; СIтакже больше снаружи.
Впокое мембрана нервных клеток наиболее проницаема для К+, менее — для СI- и очень мало проницаема для Na+/ Проницаемость мембраны нервного волокна для Na+ B покое в 100 раз меньше, чем для K+. Для многих анионов органических кислот мембрана в покое совсем непроницаема.
Всилу градиента концентраций К+ выходит на наружную поверхность клетки, вынося свой положительный заряд. Высокомолекулярные анионы не могут следовать за К+ из-за непроницаемости для них мембраны. Ион Na+ также не может возместить ушедшие ионы калия, ибо проницаемость мембраны для него значительно меньше. СIпо градиенту концентраций может перемешаться только внутрь клетки, увеличивая тем самым отрицательный заряд внутренней поверхности мембраны. Вследствие такого перемещения ионов возникает поляризация мембраны, когда наружная ее поверхность заряжается положительно, а внутренняя — отрицательно.
Электрическое поле, которое создастся на мембране, активно вмешивается в распределение ионов между внутренним и наружным содержимым клетки. По мере возрастания положительного заряда на наружной поверхности клетки иону К+ как положительно заряженному становится все труднее перемещаться изнутри наружу. Он движется как бы в гору.
Чем больше величина положительного заряда на наружной поверхности, тем меньшее количество ионов К+ может выходить на поверхность клетки. При определенной величине потенциала на мембране количество ионов К+, пересекающих мембрану в том и другом направлении, оказывается равным, т.е. концентрационный градиент калия уравновешивается имеющимся на мембране потенциалом. Потенциал, при котором диффузионный поток ионов
становится равным потоку одноименных ионов, идущих в обратном направлении, называют потенциалом равновесия для данного иона. Для ионов К+ потенциал равновесия равен -90 мВ. В миелинизированных нервных волокнах величина потенциала равновесия для ионов СIблизка к значению мембранного потенциала покоя (-70 мВ). Поэтому, несмотря на то что концентрация ионов СIснаружи волокна больше, чем внутри его, не отмечается их одностороннего тока в соответствии с градиентом концентраций. В этом случае разность концентраций сбалансирована потенциалом, имеющимся на мембране.
Ион Na+ по градиенту концентраций должен был бы входить внутрь клетки (его потенциал равновесия составляет +60 мВ), и наличие отрицательного заряда внутри клетки не должно было бы препятствовать этому потоку. В этом случае входящий Na+ нейтрализовал бы отрицательные заряды внутри клетки. Однако этого в действительности не происходит, так как мембрана в покое малопроницаема для Na+.
Важнейшим механизмом, поддерживающим низкую внутриклеточную концентрацию ионов Na+ и высокую концентрацию ионов К+, является натрий-калиевый насос (активный транспорт). Известно, что в клеточной мембране имеется система переносчиков, каждый из которых связывается, стремя находящимися внутри клетки ионами Na+ и выводит их наружу. С наружной стороны переносчик связывается с двумя находящимися вне клетки ионами К+ которые переносятся в цитоплазму. Энергообеспечение работы систем переносчиков обеспечивается АТФ. Функционирование насоса по такой системе приводит к следующим результатам:
поддерживается высокая концентрация ионов К+ внутри клетки, что обеспечивает постоянство величины потенциала покоя. Вследствие того, что за один цикл обмена ионов из клетки выводится на один положительный ион больше, чем вводится, активный транспорт играет роль в создании потенциала покоя. В этом случае говорят об электрогенном насосе, поскольку он сам создает небольшой, но постоянный ток положительных зарядов из клетки, а потому вносит прямой вклад в формирование отрицательного потенциала внутри нее. Однако величина вклада электрогенного насоса в общее значение потенциала покоя обычно невелика и составляет несколько милливольт;
поддерживается низкая концентрация ионов Na+ внутри клетки, что, с одной стороны, обеспечивает работу механизма генерации потенциала действия, с другой — обеспечивает сохранение нормальных осмолярности и объема клетки;
поддерживая стабильный концентрационный градиент Na+, натрийкалиевый насос способствует сопряженному К+, Na+ -транспорту аминокислот и Сахаров через клеточную мембрану.
Таким образом, возникновение трансмембранной разности потенциалов (потенциала покоя) обусловлено высокой проводимостью клеточной мембраны в состоянии покоя для ионов К+, СI-, ионной асимметрией концентраций ионов К+ и ионов СI-, работой систем активного транспорта (Na+/K+ -АТФаза), которые создают и поддерживают ионную асимметрию.
8. Что такое потенциал действия?
Потенциал действия — это возбуждение клетки, быстрое колебание мембранного потенциала вследствие диффузии ионов в клетку и из клетки.
9. Охарактеризуйте фазы развития потенциала действия.
При действии раздражителя на клетки возбудимой ткани сначала очень быстро активируются и инактивируются натриевые каналы, затем с некоторым опозданием активируются и инактивируются калиевые каналы.
Вследствие этого ионы быстро диффундируют в клетку или из нее согласно электрохимическому градиенту. Это и есть возбуждение. По изменению величин и знака заряда клетки выделяют три фазы:
1-я фаза — деполяризация. Уменьшение заряда клетки до нуля. Натрий движется к клетке согласно концентрационному и электрическому градиенту. Условие движения: открыты ворота натриевого канала;
2-я фаза — инверсия. Изменение знака заряда на противоположный. Инверсия предполагает две части: восходящую и нисходящую.
Восходящая часть. Натрий продолжает двигаться в клетку согласно концентрационному градиенту, но вопреки электрическому градиенту (он препятствует).
Нисходящая часть. Калий начинает выходить из клетки согласно концентрационному и электрическому градиенту. Открыты ворота калиевого канала;
3-я фаза — реполяризация. Калий продолжает выходить из клетки согласно концентрационному, но вопреки электрическому градиенту.
Суть механизма генерации ПД состоит в том, что рецепторный потенциал вызывает возникновение локальных круговых токов между деполяризованной рецепторной мембраной немиелинизированной части нервного окончания и мембраной первого перехвата Ранвье. Эти токи, носителями которых являются ионы Na+, К+, СI- и другие минеральные ионы, «протекают» не только вдоль, но и поперек мембраны нервного волокна в области перехвата Ранвье. В мембране перехватов Ранвье в отличие от рецепторной мембраны самого нервного окончания имеется большая плотность ионных потенциалзависимых натриевых и калиевых каналов.
При достижении на мембране перехвата Ранвье величины деполяризации около 10 мВ происходит открытие быстрых потенциалзависимых натриевых каналов и через них в аксоплазму по электрохимическому градиенту устремляется поток ионов Na+. Он обусловливает быструю деполяризацию и перезарядку мембраны перехвата Ранвье. Однако одновременно с открытием быстрых потенциалзависимых натриевых каналов в мембране перехвата Ранвье открываются медленные потенциалзависимые калиевые каналы и из аксоилазмы начинают выходить ионы К+ Их выход запаздывает по отношению ко входу ионов Na+.
Таким образом, входящие с большой скоростью в аксоплазму ионы Na+ быстро деполяризуют и перезаряжают на короткое время (0,3-0,5 мс) мембрану, а выходящие ионы К+ восстанавливают исходное распределение зарядов на мембране (реполяризуют мембрану). В результате во время механического воздействия на тельце Пачини силой, равной или превышающей пороговую, на мембране ближайшего перехвата Ранвье наблюдается кратковременное колебание потенциала в виде быстрой деполяризации и реполяризации мембраны, т.е. генерируется ПД (нервный импульс).
Поскольку непосредственной причиной генерации ПД является рецепторный потенциал, то его в этом случае еще называют генераторным потенциалом. Число генерируемых в единицу времени одинаковых по амплитуде и длительности нервных импульсов пропорционально амплитуде рецепторного потенциала, а, следовательно, силе давления на рецептор. Процесс преобразования информации о силе воздействия, заложенной в амплитуде рецепторного потенциала, в число дискретных нервных импульсов получил название дискретного кодирования информации.
Более подробно ионные механизмы и временная динамика процессов генерации ПД изучены в экспериментальных условиях при искусственном воздействии на нервное волокно электрическим током различной силы и длительности.
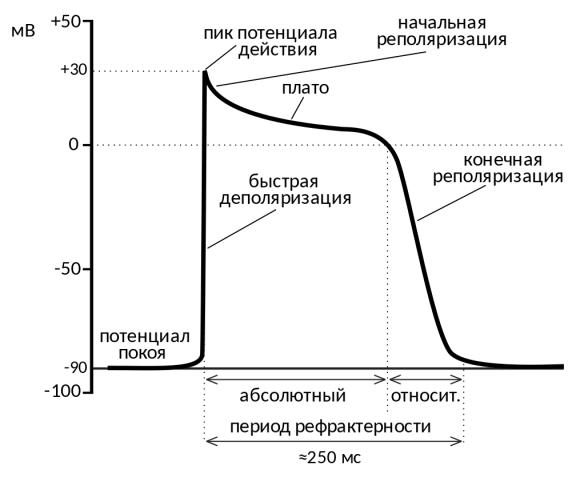
10. Что такое рефрактерные периоды? Какие рефрактерные периоды Вам известны?
Р.П - период времени после возникновения на возбудимой мембране потенциала действия, в ходе которого возбудимость мембраны
снижается, а затем постепенно восстанавливается до исходного уровня.
Абсолютный рефрактерный период — интервал, в течение которого возбудимая ткань не способна генерировать повторный потенциал действия (ПД), каким бы сильным ни был инициирующий стимул.
Относительный рефрактерный период — интервал, в течение которого возбудимая ткань постепенно восстанавливает способность формировать потенциал действия. В ходе относительного рефрактерного периода стимул, более сильный, чем тот, который вызвал первый ПД, может привести к формированию повторного ПД.