

У всех черепных животных передний конец мозговой трубки преобразуется в головной мозг. На ранних стадиях эмбрионального развития головной мозг у миног представлен тремя утолщениями стенок переднего конца мозговой трубки, из которых впоследствии развиваются три отдела мозга: передний, средний и задний, или ромбовидный. Передний мозг—prosencephalon—находится в связи с органом обоняния и является, следовательно, обонятельным мозгом; средний мозг—mesencephalon—имеет отношение к органу зрения и, таким образом, функционально является зрительным мозгом и, наконец, задний, или ромбовидный, мозг—rhombencephalon—связан с развитием органов VlIl и X пар черепномозговых нервов, т. е. с органами равновесия и органом боковой линии, а также с внутренними органами, вследствие чего он и является высшим моторным центром, координирующим через спинной мозг весь двигательный аппарат и управляющим работой внутренних органов. У всех остальных вышестоящих черепных животных эмбриональный головной мозг представлен пузыревидным расширением нервной трубки, лежащим впереди хорды. Этот прехордальный пузырь называется первичным мозговым пузырём — archencephalon (рис. 156—7). Вентральной складкой—plica ventralis (13)—он вскоре отделяется от вторичного мозгового пузыря—deaterencephalon (12),—располагающегося эпихордально (над хордой).
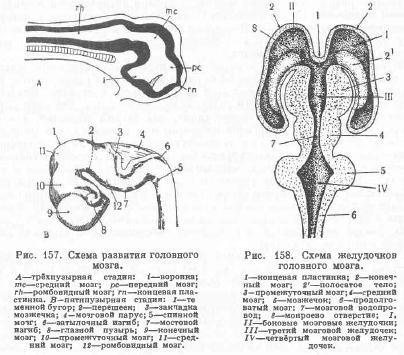
Одновременно с появлением вентральной складки вследствие вентрального изгиба прехордального мозгового пузыря от вторичного мозгового пузыря обособляется третий мозговой пузырь, или средний мозг,—mesencephalon (рис. 157, гпс); прехордальный пузырь получает название переднего мозга—prosencephalon (рс),—а эпихордальный пузырь—ромбовидного мозга—rhombencephalon (rh). С дорзальной поверхности все три отдела головного мозга довольно резко отграничиваются друг от друга поперечными коммиссурами или спайками, из нервных волокон, как впереди, так и позади среднего мозга. В дальнейшем из переднего мозга по обе стороны от концевой пластинки (на месте невропора) возникает в виде парного мозгового пузыря—конечный мозг—telencephalon (В—9),—а остающаяся часть переднего мозга формирует промежуточный мозг—diencephalon (10). Почти одновременно из ромбовидного мозга выделяется задний мозг—metencephalon, из которого впоследствии развивается мозжечок, а у млекопитающих, кроме того, и мост, а главная масса ромбовидного мозга образует продолговатый мозг—myelencephalon, или medulla oblongata. Полости первичных трёх мозговых пузырей называются мозговыми желудочками. Из полости конечного мозга обособляются парные боковые мозговые желудочки (рис. 158—I и II); полость промежуточного мозга называется третьим мозговым желудочком (III). Он сообщается с боковыми посредством межжелудочковых, или монроевых, отверстий (8). Желудочек среднего мозга в результате разрастания стенок мозгового пузыря превращается в канал—мозговой водопровод (7); он соединяет третий мозговой желудочек с четвёртым мозговым желудочком (IV).Последний размещается в ромбовидном мозге, между продолговатым мозгом и мозжечком, и каудально сообщается с центральным каналом спинного мозга. Из описанных мозговых пузырей формируется мозг взрослого животного; при этом отдельные участки их то очень значительно разрастаются, то, напротив, пожизненно сохраняют своё эмбриональное состояние. Ближайшим фактором, обусловливающим развитие головного мозга, является диференциация органов чувств, особенно таких, как органы обоняния, зрения и слуха (у наземных), и органов произвольного движения, что сопровождается появлением различных координирующих центров с их проводящими путями. Так, у селяхий, ориентирующихся при своём движении главным образом при помощи обоняния, максимального развития в головном мозге достигают обонятельные доли конечного мозга. Напротив, у костистых рыб преобладающее значение имеют органы зрения, а в соответствии с этим сильнее развиваются зрительные центры в среднем мозге, в то время как обонятельный мозг усложняется мало. Задний мозг (мозжечок) совершеннее у животных, быстро двигающихся, а у медленно двигающихся он довольно элементарен или даже отсутствует. У наземных животных головной мозг усложняется в своём строении вследствие возникновения новых (высших) центров в конечном мозге и их связей как с различными отделами головного и спинного мозга, так и со всеми органами чувств (экстеро- и интерореценторами). В результате в конечном мозге развивается отдел, называемый корой больших полушарий, являющихся высшим центром, координирующим и контролирующим всю деятельность животного организма во внешней среде и все процессы, протекающие внутри него, так как в коре полушарий замыкаются все возникающие рефлексы.
Ромбовидный мозг
Ромбовидный мозг состоит из продолговатого мозга и заднего мозга, представленного мозжечком и мостом.
Продолговатый мозг
Продолговатый мозг—medulla oblongata s. myelencephalon (рис. 159 и 160—5)—построен наиболее однообразно у животных, так как у всех у них от него отходят черепномозговые нервы, имеющие непосредственное отношение к жаберному аппарату с его производными, к органу равновесия и слуха, внутренностям и сердцу. Он является непосредственным продолжением в сторону головы спинного мозга, однако резко отличается от последнего своей структурой. Продолговатый мозг сформировался в результате разрастания боковых стенок эмбриональной мозговой трубки. Так как боковые пластинки при этом отодвинулись друг от друга и расположились латерально от основных пластинок, то полость ромбовидного мозга значительно расширилась и превратилась в четвёртый мозговой желудочек. Дно желудочка у высших животных принимает форму ромбовидной ямки, на поверхности которой, даже у взрослых млекопитающих, хорошо заметна граница между боковой и основной пластинками в виде пограничной борозды—sulcus limitans. Боковые пластинки особенно значительны у рыб; они формируют латеральные слуховые доли—lobus acusticolateralis,—которые краниально обособляются даже в ушки ромбовидного мозга—auriculae. Они содержат ядра VIII пары нервов, находящиеся в связи с органом равновесия, со всей мускулатурой тела (через спинной мозг) и с мозжечком. Из них особого внимания заслуживает ядро Дейтерса, как наиболее постоянное у животных (рис. 163—1). Вследствие расхождения дорзальных краёв боковых пластинок пластинка покрышки сильно растягивается, но даже у взрослых высших позвоночных она сохраняется в эмбриональном состоянии в виде эпителиальной пластинки—lamina epithelialis,—состоящей из одного слоя клеток эпендимы. Она срастается с сосудами мягкой мозговой оболочки и у взрослых животных именуется каудальным мозговым парусом, или сосудистым сплетением четвёртого мозгового желудочка,—velum medullare caudale, resp. plexus chorioideus ventriculi quarti (рис. 157—B, 4). B нём находятся отверстия, через которые полость желудочка сообщается с подоболочечным пространством. Раздвиганием в стороны боковых пластинок объясняется и своеобразный выход нервов из продолговатого мозга: моторные нервы располагаются более медиально, а чувствительные—более латерально. Так как продолговатый мозг лежит между спинным мозгом и передними отделами головного мозга, составляющими большой мозг, от которого он отделяется сужением—перешейком—isthmus (2),—то центральные проводящие пути, идущие в ту или другую сторону, разбивают серое мозговое вещество его на ряд отдельных ядер—скоплений нервных клеток. Одни из ядер являются клеточными центрами отходящих от продолговатого мозга черепномозговых нервов с V по XIl пару. Другие, напротив, служат промежуточными центрами для проводящих путей, направляющихся в головной мозг из спинного или обратно. К числу таковых относятся ядра дорзальных канатиков и древнейший двигательный центр—моторное ядро чепца, имеющееся уже у рыб. Волокна из ядер дорзального канатика следуют у водных в мозжечок и средний мозг, а у наземных, кроме того, в промежуточный мозг. За счёт клеток моторного ядра чепца у наземных животных обособляются так называемые оливы: назальные (верхние) и каудальные (нижние), становящиеся ассоциационными центрами. Назальные оливы появляются впервые у амфибий в связи с диференциацией улитки (орган слуха). Они вставлены в латеральный лемниск, т. е. в петлю перекрещивающихся проводящих путей, соединяющих ядра улиткового нерва (VIII пары) со зрительными буграми. Пучок этих волокон лежит поверхностно на переднем конце продолговатого мозга; он более сильно развит у млекопитающих и известен под названием трапецоидного тела— corpus trapezoideum (рис. 175—14). Каудальные оливы хорошо выражены у птиц и особенно у млекопитающих; они имеют своеобразную складчато-подковообразную форму и получают импульсы из ядер дорзального канатика, из спинного мозга, из зрительных бугров и посылают импульсы обратно в спинной мозг и в мозжечок (рис. 163). Двигательные проводящие пути, происходящие из коры полушарий большого мозга, формируют у млекопитающих на вентральной продолговатого мозга пучки, известные под названием пирамид. Максимальных размеров они достигают у человека.