

3 КУРС / Разобранные билеты на экзамен по биологии микроорганизмов
.pdf1. Особенности строения прокариотной клетки и ее химический состав.
-нет ядра (нуклеоид)
-нет органелл (мезосомы- локальные впячивания ЦПМ)
-более просто устроены
-клеточная стенка (не всегда)
-меньше размер (1 – 10 мкм)
-рибосомы 70S
-бинарный тип деления
-внехромосомная днк (плазмиды)
1 - гранулы поли-βоксибутирата; 2 - жировые капельки; 3 - включения серы; 4 - трубчатые тилакоиды; 5 - пластинчатые тилакоиды; 6 - хлоросомы; 7 - хроматофоры; 8 - нуклеоид; 9 - рибосомы; 10 - цитоплазма; 11 - базальное тельце; 12 - жгутики; 13 - капсула; 14 - клеточная стенка; 15 - ЦПМ; 16 - мезосома; 17 - газовые вакуоли; 18 - ламеллярные структуры; 19 - гранулы полисахарида; 20 - гранулы полифосфата; 21 - карбоксисомы.
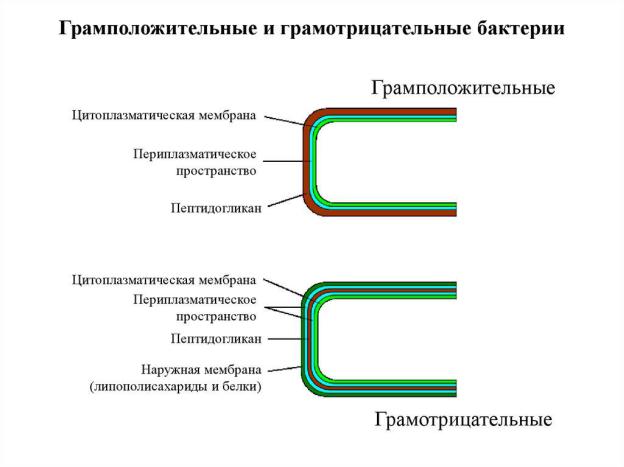
2. Структура и функции клеточной стенки бактерий.
Клеточная стенка бактерий выполняет следующие функции:
•механическую защиту клетки от воздействий факторов окружающей среды;
•обеспечивает поддержание формы бактериальной клетки;
•дает возможность клетке существовать в гипотонических растворах;
•осуществляет транспорт веществ и ионов (характерно для грамотрицательных бактерий, имеющих наружную мембрану, которая является дополнительным барьером для их поступления; основным барьером служит цитоплазматическая мембрана);
•препятствует проникновению в клетку токсических веществ (также более характерно для грамотрицательных бактерий, имеющих наружную мембрану);
•на клеточной стенке находятся рецепторы, на которых адсорбируются бактериофаги и бактериоцины;
•в клеточной стенке находятся антигены (липополисахариды у грамотрицательных бактерий и тейховые кислоты у грамположительных бактерий);
•на клеточной стенке находятся рецепторы, ответственные за взаимодействие клеток донора и реципиента при конъюгации бактерий.
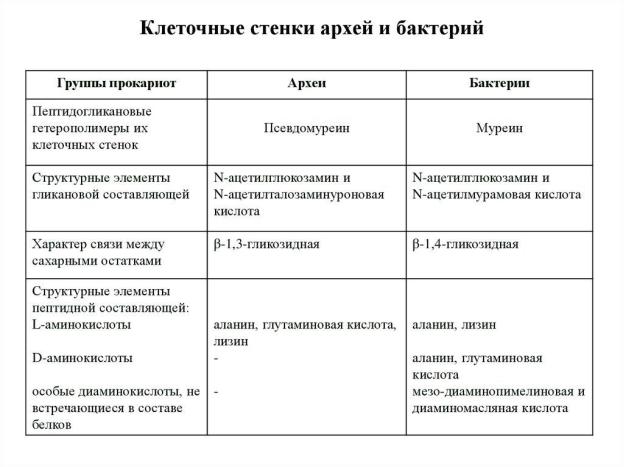
3. Структура и функции клеточной стенки архей.
Признаки по родам:
Псевдомуреин (Methanobacterium)
Кислый гетерополисахарид (Halococcus) Метанохондроитин (Methanosarcina)
Белок или гликопротеин (Pyrodictium occultum)
Отсутствуют клеточные покровы, только ЦПМ (Thermoplasmatales)
S-слои архей состоят из одного типа гликопротеина, способного к самосборке с образованием упорядоченной структуры.
4. Отличия и особенности прокариот без клеточной стенки.
Прокариоты, не имеющие клеточной стенки. Впервые бактерии, не имеющие клеточной стенки, были обнаружены при воздействии на них лизоцимом ферментом из группы гликозидаз, содержащимся в яичном белке, слюнной жидкости и выделяемом некоторыми бактериями.
Бактерии с частично (сферопласты) или полностью (протопласты) утраченной клеточной стенкой можно получать при воздействии определенными химическими веществами в лабораторных условиях. При нару шении синтеза клеточной стенки бактерий под влиянием фермента лизо цима или пенициллина, а также защитных факторов организма образуются клетки с измененной, часто шаровидной, формой (протопласты).
После удаления ингибитора синтеза клеточной стенки такие изме ненные бактерии могут реверсировать, то есть приобретать полноценную клеточную стенку и восстанавливать исходную форму.
Бактерии сферопластного или протопластного типа, утратившие спо собность к синтезу пептидогликана под влиянием антибиотиков или дру гих факторов, но сохранившие способность к размножению, называются L формами. Lформы могут возникать и в результате мутаций. Они пред ставляют собой осмотически чувствительные шаровидные, колбовидные клетки различной величины, в том числе и проходящие через бактериальные фильтры. Lформы могут образовывать многие бактерии возбудите ли инфекционных болезней.
5. Химический состав и функции цитоплазматических мембран прокариот.
На долю ЦПМ приходится 8-15% сухого вещества клеток.
По химическому составу ЦПМ – белковолипидный комплекс: белки – от 50 до 75%, липиды – от 15 до 45%. углеводы – около 5 % Главный компонент – фосфолипиды, производные 3-фосфоглицерина
Фосфолипиды эубактерий:
Насыщенные или мононенасыщенные жирные кислоты С 16-18
У цианобактерий найдены полиненасыщенные жирные кислоты C16:2, C18:2, C18:3,
C15:4
Фосфолипиды архей:
Липиды архей образованы глицерином, связанным с боковой цепью фитанила эфирной связью
У термофильных архей (Thermoplasma, Sulfolobus, Thermoproteus) в бифитанильных тетраэфирах может содержаться 1-4 циклопентановых кольца
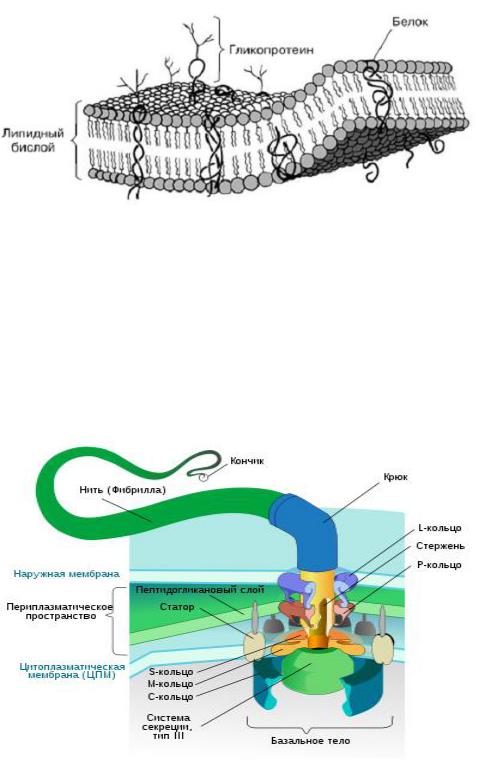
Функции ЦПМ:
1.Барьерная
2.Ферментная
3.Энергетическая
4.Участие в репликации
5.Интегрирующая роль в организме (тилакоиды, хроматофоры, локальные впячивания)
6.Транспортная (Диффузия, активный транспорт, перенос молекул, заряд)
6. Строение и биохимические особенности жгутиков прокариот. Механизмы движения.
-Базальное тело, Крюк, Нить (фибрилла, состоит из белка - флагеллина)
-Наращивание жгутика происходит с дистального конца, куда субъединицы тубулина поступают по внутреннему каналу.
-Движение жгутика обеспечивается энергией трансмембранного электрохимического потенциала. Жгутик вращается за счет M-кольца, другие служат для крепления стержня.S, M –кольца обязательны, L,P- есть только у грам-

7. Запасные вещества прокариот и их назначение.
Полисахариды:
-гликоген -крахмал -гранулеза
Углеводородные гранулы
Липиды (в том числе поли- - оксимасляная кислота)
Полипептиды
Полифосфаты Отложения серы
Кристаллоподобные включения (параспоральные)
8. Генетический аппарат прокариот. Бактериальная хромосома. Внехромосомные элементы.
Нуклеоид:
Длина молекулы – 1 мм и более, диаметр нитей – около 2 нм. Молекулярная масса – 1-3 × 109 Да.
Каждая клетка содержит 1 хромосому, кольцевую ДНК.
При действии неблагоприятных факторов происходит образование копий хромосомы. Нейтрализация зарядов осуществляется взаимодействием ДНК с полиаминами и ионами Mg2+ . Содержание пар оснований А+Т и Г+Ц является постоянным для вида. Молярная доля ГЦ в ДНК – от 23 до 75%.
Плазмиды –замкнутые в кольцо ДНК, способные к независимой репликации, содержат дополнительные гены, необходимые в специфических условиях (устойчивость к антибиотикам)
Транспозоны – мобильные сегменты, способные перемещаться из одной части хромосомы к другой, или во внехромосомные ДНК (в том числе в другие клетки). Эти сегменты неспособны к автономной репликации. Вносят нарушения в существующие гены.
Деление молекулы ДНК (репликация) происходит по полуконсервативному механизму

9. Общая характеристика метаболизма прокариот. Связь катаболических и анаболических путей.
Метаболизм - совокупность протекающих в клетке процессов, обеспечивающих воспроизводство биомассы
Энергетический метаболизм — поток реакций, сопровождающихся мобилизацией энергии и преобразованием ее в электрохимическую ( H + ) или химическую (АТФ) форму.
Конструктивный метаболизм (биосинтез) — поток реакций, в результате которых за счет поступающих извне веществ строится вещество клеток.
Метаболические пути:
1)Периферический метаболизм
2)Промежуточный метаболизм
3)Образование конечных продуктов
10. Потребности прокариот в питательных веществах: органические и неорганические источники углерода.
ИСТОЧНИК УГЛЕРОДА:
Неорганический (CO2, карбонаты ) (автотрофы) Органический (гетеротрофы)
11. Потребности прокариот в питательных веществах: органические и неорганические источники азота.
ИСТОЧНИК АЗОТА:
Восстановленные формы (соли аммония, мочевина, аминокислоты, пептиды) аммонификаторы
Окисленные формы (NO3 , NO2 ) нитрификаторы, денитрификаторы Молекулярный азот (N2 ) азотфиксаторы
12. Потребности прокариот в питательных веществах: микроэлементы и факторы роста. Прототрофные и ауксотрофные бактерии.
Прототрофные микроорганизмы, не требующие (в отличие от ауксотрофов) для своего развития готовых витаминов, аминокислот или др. факторов роста, а синтезирующие их

из минеральных или органических соединений. Один и тот же микроорганизм может быть прототрофным по одному фактору роста, но ауксотрофным по другому.
БАКТЕРИАЛЬНЫЕ ФАКТОРЫ РОСТА, стимуляторы роста бактерий, вещества, необходимые для жизнедеятельности и роста гетеротрофных микроорганизмов, к-рые они не могут синтезировать (совсем или в достаточном кол-ве) и поэтому должны получать из окружающей среды. Отсутствие или недостаток приводит к задержке роста микробов. Для удовлетворения потребностей микробов в Б. ф. р. достаточно присутствия ничтожных кол-в этих веществ.
Потребность бактерий к Б. ф. р. наиболее сильно выражена у патогенных микроорганизмов. К наиболее важным Б. ф. р. относятся витамины: никотиновая к та, пантотеновая к-та, тиамин, рибофлавин, пиридоксин, биотин, дианкобаламин, парааминобензойная к-та, фолиевая к-та, холин; гемин (фактор X); пуриновые и пиримидиновые основания (аденин, гуанин, ксантин, гипоксантин, цитозин, тимин, урацил); аминокислоты: триптсфан, лизин, холин, глутаминовая к-та, аргинин, метионин, валин, лейцин, треонин, гистидин, фенилаланин и др.
Учитывая потребность тех или иных видов микробов в определённых факторах роста, можно регулировать их жизнедеятельность, что важно для биол. пром-сти. Б. ф. р. используют в лабораторной практике при приготовлении селективных питат. сред.
Микроэлементы – это марганец, Молибден активирует целую группу ферментов, ответственных за процессы брожения, фиксации азота, нитрификации; цинк, медь, кобальт, никель, ванадий, бор, хлор, натрий, селен, кремний, вольфрам и другие
13. Универсальные формы энергии в клетке прокариот.
АТФ + Трансмембранный заряд ( H+) - энергия находится в мобильной форме.
Количество освобождающейся энергии определяется градиентом окислительно восстановительных потенциалов при переносе электронов от донора к акцептору.
Энергия, получаемая в процессах брожения, дыхания или фотосинтеза, запасается в определенных формах. (атф)
•Атф образуется в реакциях субстратного фосфорилирования (гликолиз): Субстрат ~ Ф + АДФ =) субстрат + АТФ; Окислительное и фотосинтетическое – АТФ образуется в процессе электронного транспорта при градиенте ионов водорода.
•При переносе электронов на ЦПМ возникает трансмембранный электрохимический
градиент ионов водорода, обозначаемый символом ( H+) +, и измеряемый в вольтах.
H+) +, и измеряемый в вольтах.
14.Основные макроэргические соединения прокариот и их функции.
Макроэргические молекулы (макроэрги) — биологические молекулы, которые способны накапливать и передавать энергию в ходе реакции. При гидролизе одной из связей высвобождается более 20 кДж/моль. По химическому строению макроэрги —
чаще всего ангидриды фосфорной и карбоновых кислот, а также слабых кислот, какими являются тиолы и енолы.
• Соединения с высокоэнергетической фосфатной связью (фосфоенолпируват, 1.3- дифосфоглицерат-соед. Гликолиза)
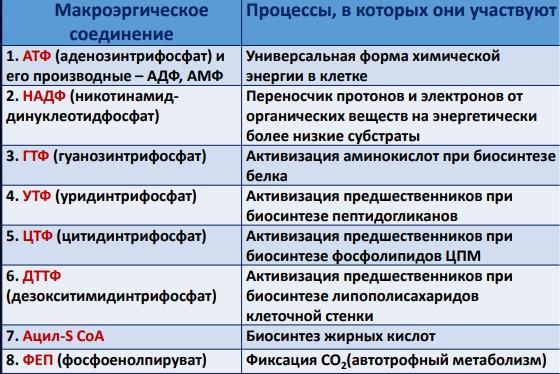
• Соединения с высокоэнергетической тиоэфирной связью (ацетил-коа, сукцинил-коа)
•Нуклеотид ди- и трифосфаты (Аденозинтрифосфат (АТФ), Гуанозинтрифосфат (ГТФ) Уридинтрифосфат( УТФ))
15. Способы получения энергии прокариотами: общая характеристика процессов.
МЕТАБОЛИЧЕСКИЕ ГРУППЫ ПРОКАРИОТ:
1.Фотоавтотрофы (цианобактерии, серные, пурпурные бактерии)
2.Фотогетеротрофы (галобактерии, осуществляющие родопсиновый фотосинтез, пурпурные бактерии, осуществляющие аноксигенный фотосинтез)
3.Хемолитоавтотрофы (метановые бактерии и сульфатредуцирующие - Tiobacteriaceae, Nitrobacteriaceae)
4.Хемолитогетеротрофы (железобактерии, сульфатредуцирующие и метановые бактерии)
5.Хемоорганоавтотрофы (сульфатредукторы, которые в качестве источника энергии используют формиат, реже метанол)
6.Хемоорганогетеротрофы (большинство грам(+) бактерий; Pseudomonas, Azotobacter, E.coli и др. энтеробактерии).
16. Классификация микробных ферментов.
ФЕРМЕНТЫ МИКРООРГАНИЗМОВ:
1. Оксидоредуктазы – окислительно-восстановительные процессы. Коферменты: НАД, НАДФ, ФАД, ФМН, убихинон, глутатион, липоевая к-та Дегидрогеназы – дегидрирование субстрата, акцептор водорода – любые молекулы, кроме кислорода -
Редуктазы – если перенос водорода трудно доказуем - Оксидазы – окисление субстратов, акцептор электронов – молекулярный кислород -
Моно(ди)оксигеназы – внедрение атомов кислорода в молекулу субстрата - Пероксидазы – пероксид водорода в качестве акцепторов
Пример: алькогольдегидрогеназа, сукцинатдегидрогеназа
2.Трансферазы – перенос группировок с одной молекулы на другую, гексокиназа,
аспартатаминотрансфераза
3.Гидролазы – расщепление сложных соединений (за исключением С-С связей) с одновременным присоединением воды. Холинэстераза, таг-липаза
Эстеразы-гидролиз сложноэфирных связей, Липазы-гидролиз жиров, Фосфотазыгидролиз моноэфиров фосфорной кислоты,
Гликозидазы – гидролизуют О- и S гликозидные связи Протеазы и пептидазы– гидролиз белков и пептидов Нуклеазы – гидролиз НК
4.Лиазы – разрыв C-O, C-N, C-C и других связей отщепление группировок от субстрата без участия воды, образование соединений с двойной связью, присоединение группировок по месту двойной связи (альдолаза). Аденилатциклаза, пируватдекарбоксилаза,фумараза
5.Изомеразы – превращение органических соединений в их изомеры
(Фосфоглюкомутаза, Рибулозофосфат-3- эпимераза)
6. Лигазы – реакции синтеза, используют энергию АТФ/ГТФ (Глутаминсинтетаза Пируваткарбоксилаза, Сукцинил-КоА-синтетаза)
Конститутивные – ферменты, синтезируемые в клетке вне зависимости и от наличия в среде субстрата.
Индуцибельные – ферменты, синтезируемые только в том случае, если имеется субстрат, превращение которого они катализируют.
Эндоферменты осуществляют каталитические реакции внутри клетки.
Экзоферменты - выделяются клеткой во внешнюю среду (в основном это гидролазы): - прочно связаны с клеточной стенкой - действуют в периплазме
17. Пути превращения гексоз: гликолиз.
ПУТИ КАТАБОЛИЗМА ГЕКСОЗ:
1.Путь Эмбдена-Мейергофа-Парнаса – фуктозобисфосфатный путь, гликолиз.
2.Путь Варбурга-Диккенса-Хореккера – гексозомонофосфатный путь, пентозофосфатный окислительный путь.
3.Путь Энтнера-Дудорова – 2-кето-3-дезокси-6- фосфоглюконатный (КДФГ).
Гликолиз. Унификация – превращения различных субстратов в один, исходный для дальнейшего метаболизирования.