

2 курс / Физиология / [ЗАДАЧИ] по физиологии
.pdf28. В эксперименте при раздражении сильным током регистрировали биопотенциалы скелетной мышцы, длительность абсолютной рефрактерной фазы, которой составляла 5,0 мс. Какова реакция мышцы на раздражении различной частоты, какова примерно лабильность мышцы?
1. Период абсолютной рефрактерности соответствует фазе регенеративной деполяризации и реверсии, при этом ткань становится абсолютно невозбудимой и не отвечает на самые сильные раздражители.
2.Какова частота биопотенциалов при раздражении мышцы 100стимулов/с? Объясните результат. 3.С какой частотой будут возникать биопотенциалы при частоте раздражения 200 стимулов/с? 4.Какова реакция мышцы на раздражение 300 стимулов/с?
5.В период абсолютной рефрактерности ткань не отвечает на раздражение.
6.Лабильность – скорость протекания физиологических процессов в возбудимой ткани. Лабильность, как и ПД, определяется скоростью перемещения ионов в клетку и из клетки, которая в свою очередь зависит от скорости изменения проницаемости клеточной мембраны. При этом особое значение имеет длительность рефрактерной фазы - чем больше рефрактерная фаза, тем ниже лабильность ткани. Мерой лабильности является максимальное число ПД, которое ткань может воспроизвести в 1 с. Лабильность мышцы равна -около 200 импульсов в секунду.
29.В контрольном исследовании было установлено, что порог раздражения клетки при замыкании постоянного тока составляет 4В, хронаксия-0,3мс. Однако, при внутриклеточном раздражении током в 8В и длительности пульса в 0,5мс возбуждение не возникло. Объясните результат экс-та при условии, что клетка не была повреждена и находилась в состоянии норм. возбудимости.
1.Сила в 8В является сверхпороговой, ПД возникает.
2.Пороговая сила любого стимула в определенных пределах находится в обратной зависимости от его длительности. Эта зависимость получила название кривой “сила – длительность” или “сила – время”. 3.Аргументируйте.
4.Какова причина отсутствия возбуждения в описанном экс-те? Длительность раздражающего стимула превышает хронаксию.
5.Как изменить условия экс-та для того, чтобы при раздражении клетка возбудилась? Изменить длительность раздражающего стимула.
30.Неповрежденный нерв нервно-мышечного препарата расположен на электродах постоянного тока. При замыкании тока мышца не сократилась, при размыканиисократилась. Объясните результат.
1.Какова по вашему мнению интенсивность раздражающего тока? Слабый(пороговый) или сильный ток.
2.На чем основано ваше заключение? Почему вы исключаете возможность действия тока другой интенсивности?
3.При замыкании раздражающее действие проявляется только под катодом, при размыкании – под анодом. Т.о. мышца расположена ближе к аноду.
4.При замыкании цепи постоянного тока под анодом (допороговый, продолжительный раздражитель) на мембране развивается гиперполяризация за счет перераспределения ионов по обе стороны мембраны (без изменения ионной проницаемости мембраны) и возникающее за ней смещение уровня критической деполяризации в сторону мембранного потенциала. Следовательно, порог уменьшается, возбудимость повышается – анодическая экзальтация.При размыкании цепи мембранный потенциал быстро восстанавливается к исходному уровню и достигает сниженного уровня критической деполяризации, генерируется потенциал действия. Таким образом, возбуждение возникает только при размыкании цепи постоянного тока под анодом.
31.Два различных эфферентных нервных волокон (1 и 2) раздражали надпороговым током, вызывающим возбуждение иннервируемых ими клеток. Однако, при регистрации ПД самих волокон 1 и 2 при помощи микроэлектродов, расположенных на их поверхности, реакция оказалась неодинаковой. В волокне 1 на расстоянии до 0,2 мм от места раздражения ПД не регистровалось. Далее по ходу волокна возбуждение участки чередовались с участками, где ПД отсутствовали. В волокне 2 ПД регистрировались на всем протяжении.
1.Да, так как ПД регистрируется в перехватах Ранвье, и распространяется по мембране волокна сальтаторно 2.Отдельные участки волокна №1 покрыты миелином, имеющим свойства диэлектрика, поэтому в межперехватных участках ПД не регистрируется 3.Волокна 1 и 2 имеют различные физические свойства, так как 1 волокно является миелиновым, а 2 безмиелиновым
4.Участки волокна 1, где регистрируется ПД называется перехватом Ранвье, в этих участках отсутствует миелиновая оболочка, и большое количество натриевых каналов, активация которых приводит к деполяризации, а следовательно и к возникновению ПД 5.Для любого нервного волокна общим механизмом проведения является двухстороннее проведение
возбуждения, изолированное проведение возб-я по нервному волокну и анатомо-физиологическая целостность волокна 6.В миелиновых волокнах (№1) возбуждение возникает пи нанесении раздр – я в перехватах Ранвье и
распространаяется по мембране сальтаторно, при этом может охватывать и несколько перехватов, что является энергитически более выгодным и увеличивает скорость распространения возб-я. В безмиелиновых волокнах (№2)
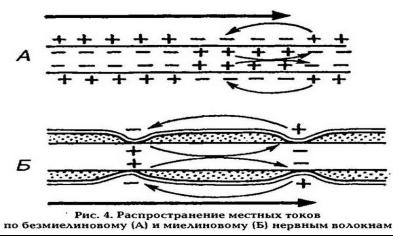
на мембране волокна между возб. И невозб. Участками возникает местный ток, который является раздражителем для соседнего невозб. Участка, но скорость передачи импульса резко снижается.
32. При определении скорости проведения возбуждения в двух нервных волокнах было установлено, что скор проведения в волокне 1 составляет 100м/с, в волокне 2 – 10 м/с.
1.Волокно 1 – миелиновое типа А наибольшего диаметра, скорость 120-70 м/с, длительность ПД минимальная Волокно 2 – миелиновое типа В меньшего диаметра, скорость 3-18 м/с, ПД более длителен
2.Cкорость проведения возбуждения в волокнах 1 и 2 зависит от диаметра волокна
3.Скорость проведения возб-я в волокнах 1 и 2 прямо пропорциональна длине межперехватных участков, а их длина тем больше чем больше диаметр волокна.
33. При определении структурных и функциональных показателей двух афферентных волокон 1 и 2 было установлено, что диаметр волокна 1 - 20 мкм, после пика его пд, развивается гиперполяризация ( 50 мс), диаметр вол 2 - 2 мкм, гиперполяризация – 500 мс
1.Миелиновые и безмиелиновые 2.Миелиновые волокна имеют миелиновую оболочку, а безмиелиновые только шванновскую оболочку.
Миелиновые – диаметр до 25 мкм, скорость 120 м/с, продолжит ПД 0,4-0,5 мсек. Безмиелиновые – до 1,3 мкм, до 2 м/с, 2мсек Безмиелиновые (безмякотные) волокна покрыты только леммоцитами (шванновскими клетками). Между ними и осевым цилиндром (аксоном нейрона) имеется щель с межклеточной жидкостью, поэтому, клеточная мембрана остается неизолированной. Импульс распространяется по волокну со скоростью всего 1-3 м/сек.Миелиновые волокна покрыты спиральными слоями шванновских клеток с прослойкой миелина - жироподобного вещества с высоким удельным сопротивлением. Миелиновая оболочка через промежутки равной длины прерывается, оставляя оголенными участки осевого цилиндра длиной » 1 мкм
3.Основные типы нервных волокон: Аα(первичные афференты мышечных веретен, двигательные волокна скелетным мышц) – 15 мкм, 70-120 м/с; Аβ ( кожные афференты прикосновения и давления)- 8мкм, 30-70 м/с; Аγ ( двигательные волокна мышечных веретен) 5мкм, 15-30 м/с; Аδ ( кожные афференты температуры и боли) < 3мкм, 12-30м/с. В(симпатические преганглионарные волокна) 3 мкм, 15 м/с. С( кожные афференты боли, симпатические постганглионарные волокна (немиелинизированные)) 1 мкм, 0,5-2 м/с
4.Волокно 1 – миелиновое типа Аα, волокно 2 – миелиновые Аδ
5.1- первичные афференты мышечных веретен, 2- кожные афференты температуры и боли
6.Лабильность определяется длительностью рефрактерного периода(фаз абсолютной рефрактерности), чем длиннее рефрактерная фаза, тем меньше лабильность.
7.Фаза абсолютной рефрактерности = фаза деполяризации(восходящая фаза ПД), 8.У волокна 1 лабильность выше, так как короче рефрактерный период, у волокна 2 – наоборот
34. В эксперименте исследовали потенциалы двух нервных волокон 1 и 2, идущих в составе смешанного нерва конечности. Группу волокон 1 составляли волокна, идущие от т.н. проприорецепторов, группу 2 – от болевых рецепторов кожи.
1-3.Группа 1 – волокна типа Аα, а группа 2 – Аδ, следовательно, волокна группы 1 имели наименьшую продолжительность ПД, а группы 2 – чуть более длительный ПД, что обусловлено различной длиной межперехватных участков(?)
4.Законы проведения вобз-я по нервам:
Закон анатомо-физиологической целостности волокна
Закон изолированного проведения возбуждения по нервному волокну Закон двухстороннего проведения возбуждения
5.В двигательных волокнах скелетных мышц, которые так же относятся к типу Аα
35. Частота раздражения нерва 500стимулов/с, возбуждение походит через поврежденный участок, в котором длительность развития пд в рез-те травмы увеличилась с 0.5 мс до 5 мс, следовые потенциалы не характерны. Сопоставить частоту пд, перед поврежденным участком и после него.
1.Совпадает, т.к. чем выше частота раздражения – тем сильнее ответ(выше частота пд)
2.В поврежденном участке возникают потенциалы меньшие по своей амплитуде, а значит и по частоте 3.Наблюдается явление парабиоза-резкого снижения лабильности 4.Лабильность - максимальное кол-во циклов возбуждения, которые может генерировать ткань в единицу
времени. Отсюда следует, что неповрежденный участок обладает большей лабильностью(длительность ПД меньше, а значит и рефрактерный период короче) по сравнению с поврежденным участком.
5.Так как парабиоз – нераспространяющееся возбуждение, то после поврежденного участка частота пд и раздражения совпадают (????)
36. При раздражении нервной ткани был зарегистрирован пд, амплитуда которого по мере увеличения силы раздражения увеличивалась, а характер кривой не менялся.
1.Объектом был смешанный нерв.
2.Если электрод расположен так, что контактирует с целым нервом, то при стимуляции нерва на некотором расстоянии от места отведения электрод сначала зарегистрирует потенциалы действия наиболее быстро проводящих волокон, а после этого–группы потенциалов других, более медленно проводящих волокон. Следовательно, потенциал действия такого нерва состоит из ответов целого спектра групп волокон с разными скоростями проведения.
37. При надпороговом раздражении двигательного нерва были зарегистрированы его биопотенциалы ( ПД) и ПД иннервируемой скелетной мышцы. ПД мышцы не возникли при действии ионов Mg, являющихся антагонистами ионов Са.
1.Возбуждение передается посредством синапса, его основные составляющие – пресинаптическая мембрана, синаптическая щель, постсинаптическая мембрана
2.Mg не действует на первое звено передачи, так как не возник ПД на мышце. Значит Мg не действует на пресинаптическую мембрану нервного окончания.
3.Не действует ни на одно звено
4.Mg не оказывает действия на синапс, а Са действует на пресинаптическую мембрану 5.Са играет ключевую роль в высвобождении квантов медиатора в синаптическую щель-
6.Деполяризация пресинаптической мамбраны – открытие Са-каналов – высвобождение медиатора 7.Высвобождается медиатор, который связывается с лиганд-зависимыми Na-каналами на постсинаптической мембране, открывая их – увеличение проницаемости для натрия – генерация пд на постсинаптической мембране
38 . При надпороговом раздражении двигательного нерва были зарегистрированы его биопотенциалы ( ПД) и ПД иннервируемой скелетной мышцы. ПД мышцы не возникает при действии магния, является антагонистом Са. При прямом раздражении мышцы она продолжала возбуждаться и сокращаться
1.Да
2.Ионы магния не принимают участия в генерации пд
3.Нет 4.Так как мышца продолжала возбуждаться и сокращаться при ее прямом радражении
40. В эксперименте на 2х мышцах ( а и б ) отдельное мышечное волокно раздражали внутриклеточно надпороговым током. Было установлено, что на мышце а – регистрировались пд не только раздражаемого волокна, но и других мышечных волокон, на мышце б – пд только раздражаемого волокна.
1.Между клетками волокон существует система межклеточных контактов(нексусы), через которые распространяется пд 2.А – гладкая висцеральная мышца(унитарная), Б – скелетная

3.В гладкой мышце нет концевых пластиной и отдельных нервных окончаний, по всей длине разветвлений адрен- и холинэргических нейронов имеются варикозные утолщения, содержацие гранулы медиатора. Поэтому по ходу следования нервного волокна могут возбуждаться многие гладкомышечные клетки, а клетки не имеющие контакта с варикозами возбуждаются посредством пд, распространяющегося на них через нексусы.
4.В скелетной мышце нет связей между отдельными мышечными волокнами, поэтому пд не распространяется на соседние волокна
41. Задачей эксперимента являлось получение гладкого и зубчатого тетануса изолированной мышцы. Длительность латентного периода ее сокращения составляла 0.02с, периода укорочения – 0.30с, периода расслабления – 0.50с. Длительность фазы абсолютной рефрактерности данной мышцы – 0.35с, фазы относительной рефрактерности – 0.10с. Каковы результаты эксперимента?
1.1 – момент раздражения, 2 – латентный период, 3 – период укорочения, 4 – период расслабления.
2.Общее условие получения тетанического сокращения
– действие на мышцу ритмических раздражений с такой частотой, что их эффекты суммируются, наступает сильное и длительное сокращение мышцы. ИЛИ наличие абсолютного рефрактерного периода (как сказано в методичке) (?)
3.При нанесении последующего раздражения во время укорочения возникает гладкий тетанус, а во время расслабления – зубчатый тетанус.
4.Минимальный промежуток времени между последовательными эффективными стимулами во время тетануса не может быть меньше рефрактерного
периода. Следовательно, для данной мышцы возможно получение зубчатого тетануса (период расслабления > рефрактерный период), и невозможно получение гладкого (период укорочения < рефрактерный период).
5.Одинаковы.
6.При пороговой силе раздражителя ответ не будет наблюдаться ни во время фазы абсолютной рефрактерности, ни во время относительной. А при максимальной силе возможен будет ответ во время фазы относительной рефрактерности. Однако на частоту раздражения данной мышцы это не повлияет, так как останется: период расслабления > рефрактерный период и период укорочения < рефрактерный период.
42.Сопоставьте динамическую работу, выполненную изолированной мышцей лягушки при однократном подъеме грузов в 10г, 30г, 410г, при условии, что абсолютная сила мышцы составляет 400г.
1.Динамическая работа – работа мышцы, при которой происходит перемещение груза и движение костей в суставах.
2.Учитывая, что абсолютная сила мышцы составляет 400г, а необходимо поднять груз 410г, то в данном случае выполненная работа будет равняться нулю. Поэтому при грузе 10г работа будет больше, чем при грузе 410г.
3.Работа при грузе 30г больше работы при грузе 10г. Высота подъема увеличилась.
4.Исходя из формулы для измерения динамической работы мышц (А = РН, где А – динамическая работа мышцы, кгм, Р – масса груза, кг, Н – высота подъема груза, м.), динамическая работа мышцы прямо пропорциональна массе груза и высоте подъема груза.
5.По мере увеличения груза работа сначала увеличивается, а затем постепенно уменьшается. При очень большом грузе, который мышца неспособна поднять, работа становится равной нулю.
Наибольшую работу мышца совершает при некоторых средних нагрузках.
43. На скелетной мышце были проведены 2 опыта (№1 и №2), в которых регистрировали ее потенциалы возбуждения (ПД). В опыте №1 на мышцах наносили прямое раздражение, в опыте №2 – непрямое. Частота раздражения в двух случаях была одинаковой и составляла 250 стимулов в секунду. Будут ли наблюдаться в опытах №1 и №2 различия в частоте ПД, возникающие в мышце при условии, что лабильность нерва равна 500 имп/с, лабильность мышцы – 300 имп/с.
1.При прямом раздражении мышцы стимулами указанной частоты, примерная частота импульсов, возникающих в мышце –250 имп/с.
2.Поскольку частота прямых раздражений - 250 стимулов в секунду, а лабильность (способность ткани воспроизводить определенное число волн возбуждения в единицу времени в точном соответствии с ритмом наносимых раздражений) мышцы – 300 имп/с.
3.При частоте раздражения нерва 250 стимулов/с, в нерве будет возникать 250 имп/с, поскольку его лабильность - 500 имп/с.
4.При непрямом раздражении указанной частоты в мышце будет возникать около 100-150 имп/с.
5. В данном случае происходит раздражение двигательного нерва, возбуждение от которого передается на мышцу посредством синапса, лабильность которого 100-150 имп/с.
6. Лабильность структур, по которым проходит возбуждение при непрямом раздражении мышцы: нерв – 500имп/с, синапс – 100-150 имп/с, мышца - 300 имп/с.
44.Животному было введено курареподобное вещество, действующее в области мышечнонервного синапса скелетной мышцы. Как доказать точную локализацию действия веществ и его механизм?
1.После введения примененного вещества скелетная мышца не будет реагировать на непрямое надпороговое раздражение.
2.Чтобы доказать, что указанное вещество не действует на двигательный нерв, а именно в области синапса, нужно измерить ПД отведением с данного нерва.
3.Доказать, что данное вещество не действует на сократительный аппарат мышц можно прямым раздражением.
4.Проверить, не действует ли данное вещество в области пресинаптической мембраны необходимо, проверив наличие медиатора ацетилхолина в синаптической щели.
5.Проверить, не оказывает ли данное вещество антихолинэстеразное действие, можно измерив ПД постсинаптической мембраны, т.к. при действии ингибитора холинэстеразы ритмическое раздражение нерва вызывает суммацию потенциала концевой пластинки, что ведет к стойкой деполяризации постсинаптической мембраны. (?)
6.Обладает ли вещество деполяризующим действием нужно проверить, измерив ПД постсинаптической мембраны. (?)
7.Проверить, обладает ли вещество конкурентным действием, можно искусственным введением ацетилхолина. (?)
8.На основании проведенного анализа остается получить результат: данное вещество воздействует на холинорецепторы – курареподобные вещества блокируют их, нарушая открытие Na+каналов на постсинаптической мембране.
45.При раздражении нерва, иннервирующего мышцу, была зарегистрирована гиперполяризация постсинаптической мембраны. К какому виду относится раздражаемая мышца?
1.Описанное изменение заряда потенциала соответствует утомлению мышцы.
2.Следовательно, синапсы, возбуждаемые при раздражении данного нерва – тормозные.
3.Нервные импульсы, приходящие в варикозы аксонов, высвобождают тормозной медиатор. Воздействуя на постсинаптическую мембрану, тормозной медиатор взаимодействует с хемовозбудимыми каналами, обладающими преимущественной проницаемостью для ионов К+. Выходящий поток калия через эти каналы вызывает гиперполяризацию постсинаптической мембраны.
4.Раздражаемая мышца – гладкая, т.к. именно гладкие мышцы могут возбуждаться и тормозиться благодаря наличию адренергических и холинергических волокон.
46.В эксперименте на нервно-мышечном препарате лягушки регистрировали биопотенциалы и сокращение мышечного волокна при увеличении частоты его раздражения от 20 до 200 стимулов в секунду. Использовали два способа раздражения мышц – прямое и непрямое. При одном способе раздражения увеличение частоты сопровождалось увеличением амплитуды сокращения и частоты биопотенциалов; при другом способе при увеличенной частоте раздражения мышца расслаблялась, биопотенциалы не регистрировались.
Объясните результат эксперимента при условии, что лабильность раздражаемой мышцы составляла 250 имп/с.
1.При непрямом раздражении мышечного волокна раздражают двигательный нерв, возбуждение по которому посредством синапса передается на мышцу.
2.Доказать, что лабильность мышцы 250 имп/с можно, увеличивая частоту ее раздражения от 200 до 300 стимулов/с. На раздражение 250 и меньше стимулов/с мышца будет отвечать соответствующим числом сокращений в единицу времени в точном соответствии с ритмом наносимых раздражений. Если частота раздражения превышает величину лабильности, то возникает ее трансформация.
3.Лабильность структур при непрямом раздражении: самая большая – нерв, меньше – мышца, еще меньше – синапс.
4.При частоте прямого раздражения 20 имп/с и 200 имп/с мышца будет генерировать ПД с соответствующей частотой. При частоте непрямого раздражения 20 имп/с мышца будет генерировать ПД соответствующей частотой, а при частоте 200 имп/с – с меньшей частотой.
5.Явление уменьшения частоты биопотенциалов и амплитуды мышечного сокращения вплоть до прекращения генерации ПД и расслабление мышц при увеличении частоты раздражения – утомление.
6.При длительном раздражении в нервных окончаниях уменьшается запас медиатора, а его синтез не поспевает за расходованием. Накапливающиеся продукты обмена в мышце понижают чувствительность постсинаптической мембраны к ацетилхолину, в результате чего уменьшается величина постсинаптического потенциала. Когда он понижается до критического уровня, в мышечном волокне не возникает возбуждения. Снижается скорость ресинтеза АТФ, необходимого для мышечного сокращения.
47.Задачей эксперимента на двух мышцах (А и Б) являлось получение суммарного мышечного сокращения. Предварительно было установлено, что длительность периода укорочения мышцы А – 50 мс, мышцы Б – 500 мс.
1.Для получения слитного суммарного сокращения мышцы А и Б необходимо их раздражать с высокой частотой.
2.Для возникновения суммации необходимо, чтобы интервал между раздражением имел определенную длительность: он должен быть длиннее рефрактерного периода, иначе на второе раздражение не будет ответа, и короче всей длительности сократительного ответа, чтобы второе раздражение подействовало на мышцу раньше, чем она успеет расслабиться после первого раздражения. Причем для возникновения слитного суммированного сокращения раздражение должно приходиться в фазу укорочения.
3.Слитное сокращение мышцы А и мышцы Б – гладкий тетанус.
4.Мышца А относится к медленным красным, Б – к быстрым белым.
5.Продолжительность суммарного сокращение мышцы А будет большей, нежели мышцы Б, т.к. медленные мышечные волокна обладают большей выносливостью, чем быстрые.
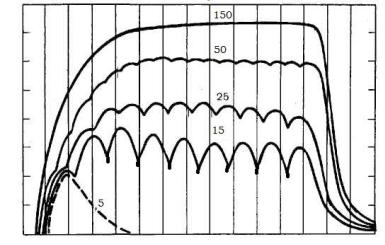
48. На скелетную мышцу лягушки наносили непрямое раздражение и регистрировали сокращения при частоте 5, 15, 25, 50 и 150 стимулов в секунду. Каковы результаты раздражения?
1.Ответ мышцы на 5 стимулов в секунду – 5 одиночных сокращений, поскольку при воздействии на мышцу непрямых раздражений с большим интервалом, при котором всякое следующее раздражение попадает в период после окончания фазы расслабления.
2.Ответ мышцы на 15 стимулов в секунду – зубчатый тетанус, так как подается много стимулов с интервалом, при котором после первого раздражения сокращение еще не закончилось, а уже возникло новое.
3.Ответ мышцы на 25 стимулов в секунду – зубчатый тетанус, поскольку подается много стимулов с интервалом, при котором после первого раздражения сокращение еще не закончилось, а уже возникло новое.
4.Ответ мышцы на 50 стимулов в секунду – гладкий тетанус, т.к. стимулы наносятся с очень малым интервалом. При сравнении с раздражением 25 стимулов/с последующее раздражение приходится на фазу укорочения, а не расслабления.
5.На непрямое раздражение в 150 имп/с мышца ответит гладким тетанусом, т.к. стимулы наносятся с очень высокой частотой.
49.Икроножную мышцу и мышцу желудка лягушки помещали в раствор, содержащий ионы Na+, K+, Ca2+ и Cl- в концентрациях, соответствующих их содержанию в тканях. На мышцы наносили прямое надпороговое раздражение и регистрировали биопотенциалы и сокращения.
Как отразилось на результатах эксперимента прибавление к раствору ионов Mg2+, который, как известно, обладает высоким сродством к активным группам кальциевых каналов и, связываясь с ними, блокирует прохождение Ca2+?
1.При блокаде кальциевых каналов нарушается мышечное сокращение.
2.Прибавление ионов магния не повлияло на генерацию ПД икроножной мышцы.
3.Генерация ПД происходит благодаря ионам Na+, К+ и не зависит от ионов Ca2+.
4.Блокада кальциевых каналов повлияла на сокращение скелетной мышцы – его осуществление стало невозможным.
5.Ионы Ca2+ необходимы для сокращения мышцы: они присоединяются к тропонину, и тот прекращает блокировать взаимодействие актина с миозином.
6.Прибавление ионов магния повлияло на генерацию ПД мышцей желудка во время ее раздражения – заблокировало ее.
7.Ионная природа ПД гладкой мышцы определяется особенностями каналов мембраны гладкой мышечной клетки. Основную роль в механизме возникновения ПД играют ионы Са2+.
8.При действии ионов Mg2+ сократимость мышцы желудка изменилась – стала отсутствовать.
9.Ионы Ca2+ перестали поступать в цитоплазму миоцитов желудка. Данные ионы должны связываться с кальмодулином, активирующим киназу легкой цепи миозина, которая приводит к гидролизу АТФ и запускает цикл образования поперечных мостиков.
10.Прибавление ионов магния не влияет на генерацию ПД исчерченной мышцы, однако блокирование кальциевых каналов данными ионами нарушает способность мышцы сокращаться. В гладкой мускулатуре блокирование кальциевых каналов ионами Mg2+ негативно воздействует и на ПД, и на сокращение.
50.Изолированные икроножные мышцы одной и той же лягушки раздражали с частотой 8 имп/с. Одна из мышц была предварительно утомлена, остальные условия проведения эксперимента были одинаковыми. Оказалось, что сокращения одной из мышц были одиночными, другая отвечала суммарными сокращениями. Какая мышца отвечала на раздражения одиночными, какая – суммарными сокращениями?
1.Неутомленная мышца не может в данных условиях отвечать на раздражение суммарным сокращением, ведь осуществляется воздействие на мышцу раздражений с большим интервалом, при котором всякое следующее раздражение попадает в период после окончания фазы расслабления.
2.Утомленная мышца может в данных условиях отвечать на раздражение суммарным сокращением. При утомлении нарастает латентный период сокращения и удлиняется период расслабления мышцы, следовательно, последующие сокращения могут попадать в фазу расслабления, вызывая суммарное сокращение.
3.Вывод: неутомленная мышца отвечала на раздражения одиночными, а утомленная – суммарными сокращениями.
51.В исследовании на человеке возможную продолжительность работы до полного утомления при подъеме груза в постоянном быстром ритме (например, 120 в минуту)
определяли, а) в условиях произвольного сгибания конечности; б) при непосредственной электрической стимуляции мышц сгибателей. Какие основные звенья прохождения импульсов от места его возникновения до мышцы в вариантах, а) и б)? Что такое утомление, где оно в первую очередь развивается в организме? Одинаково ли время утомления (продолжительность работы) в обеих случаях? Почему? В чем физиологическое значение этого? Какое значение для утомления имеет быстрый ритм работы, указанный в задаче? Как изменится результат при медленном выполнении работы (1 раз в минуту)?
1.В случае а. импульс проходит от коры головного мозга в составе пирамидных путей до мотонейронов спинного мозга и от них к мышечному волокну. В случае Б мышца раздражается непосредственно без передачи импульса по нервным волокнам.
2.Утомлением называется временное снижение или утрата работоспособности органа наступающее после нагрузок.
3.Наиболее утомляемыми при выполнении работы будут двигательные центры ЦНС и нервно-мышечные синапсы(которое связано со снижением чувствительности постсинаптической мембраны мышечного волокна к ацетилхолину.). Сам двигательный орган(мышца) будет наимение утомляем( утомление в самой мышце связано с нарушением процессов ресинтеза АТФ), именно поэтому в опыте А продолжительность работы будет гораздо меньше,чем в опыте Б( в опыте Б – нет передачи через нервно-мышечные синапсы).
4.Развивающаяся в нервно-мышечном синапсе блокада к прохождению нервного импульса является механизмом, который не позволяет истощаться энергетическим ресурсам в самой мышце,что объясняет роль утомления в организме.
5.Быстрота, с которой наступает утомление, зависит от степени нагрузки на работающие мышцы и ритма, с которым производится работа. Соответственно, при большей частоте(120 раз в минуту) утомление наступит быстрее.

ЦНС
1.В эксперименте с помощью внутриклеточного отведения исследовали функцию клеток,аксоны которых образует передние корешки спинного мозга. Что могли зарегистрировать исследователи?
Передние корешки спинного мозга образованы аксонами мотонейронов(альфа,гамма) и преганглионарных нейронов ВНС( находящихся в боковых рогах СМ)
Они иннервируют соответственно: экстрафузальные мышечные волокна,интрафузальные МВ,внутренние органы.
При раздражении проприорецепторов свою активность могли изменять гамма мотонейроны.
От влияния РФ зависела активность мотонейронов СМ через ретикулоспинальный экстрапирамидный путь.
2.При морфологическом исследовании передних и задних корешков спинного мозга установлено, что число волокон задних корешков в 20 раз больше, чем в передних. О чем свидетельствует эта закономерность?
Задние корешки спинного мозга являются афферентным входом – чувствительные. Передние корешки – эфферентный выход,двигательные.
В спинном мозге расположены 2 типа нейронов : интернейроны и эфферентные(мотонейроны и нейроны боковых рогов),При этом количество интернейронов превосходит количество эфферентных нейронов.
Тк количество интернейронов , и количество волокон задних корешков преобладает,можно сделать вывод о том,что здесь присутствуют процессы интеграции, а двигательный нейрон – общий конечный путь для импульсов от разных рецепторов.
3.В эксперименте исследовали функцию мотонейронов спинного мозга. Какие функции в организме они обеспечивают?
Тела мотонейронов располагаются в передних рогах серого вещества спинного мозга.Их аксоны входят в составе передних корешков спинного мозга. Мотонейроны составляют 3% от общего колва нейронов спинного мозга.
Различают: альфа-мотонейроны(иннервируют экстрафузальные мышечные волокна).Большие альфа-мотонейроны отвечают за фазные сокращения, а малые – за тонические.гаммамотонейроны (иннервируют интрафузальные мышечные волокна).Гамма мотонейроны не имеют прямых контактов с афферентными путями.
4.В нейроморфологических исследованиях установлено, что самыми многочисленными в спинном мозге являются интернейроны. Какие процессы в спинном мозге они обеспечивают?
Вставочные нейроны могут быть как возбуждающими,так и тормозными.
Спинальные интернейроны ветвятся в пределах нескольких смежных сегментов, образуя внутрисегментарные и межсегментарные связи.
Проэкционные интернейроны - это клетки, длинные аксоны которых формируют восходящие пути спинного мозга.
Тормозные интернейроны спинного мозга учавствуют в координации спин. рефлексов, регуляции уровня возбудимости моторных нейронов.
Тормозные нейроны(Клетки Реншоу) включены в рефлекторную спинальную дугу иннервации мышц-антагонистов. Это процесс саморегуляции альфа моторных нейронов (пресинаптическое торможение с помощью аксо-аксональных синапсов).
5.Сразу после разрыва спинного мозга на уровне нижних грудных сегментов у больного прекратились произвольные движения, и произошла потеря чувствительности, а также невозможность осуществления соматических и вегетативных рефлексов, дуги которых замыкаются ниже уровня травмы. Может ли произойти восстановление произвольных движений нижних конечностей?
При разрыве спинного мозга ниже 4-6 шейных позвонков наблюдается явление спинального шока,характеризующегося отсутствием рефлесов.У людей это состояние длится от нескольких недель до 4-5 мес.
Механизмом обеспечивающим этот состояние является отсутствие возбуждающего влияния РФ.
Через время происходит снижение порога и КУД,восстанавливается уровень возбудимости( в некоторых случаях – гиперрефлексия). Но для восстановления функций понадобится пару лет(отростки уцелевших нервных клеток восстанавливаются медленно).
Восстановление произвольных движений нижних конечностей свидетельствует о том,что при поражении спинного мозга мотонейроны не подверглись атрофии и их отростки смогли восстановиться ,после разрыва.

6.При стоянии у человека колени под действием силы тяжести постоянно стремятся согнуться. Какой рефлекторный механизм обеспечивает выпрямление конечностей?
Данный рефлекс является полисинаптическим шейным позвоночно-тоническим рефлексом.
Рецепторы шейных рефлексов содержатся в мышцах шеи, они возбуждаются при повороте или наклоне головы. Рефлекторная дуга замыкается на уровне I-III шейных сегментов. Импульсы от этих сегментов передаются на мышцы туловища и конечностей, вызывая перераспределение их тонуса, повышая в целом тонус мышц-разгибателей.
Рецепторы мышечных веретен – рефлекторное поле для поддержания мышечного тонуса. Основное различие в возбуждении сухожильного органа Гольджи по сравнению с мышечным веретеном заключается в том, что веретено определяет длину мышцы и изменение длины мышцы, тогда как сухожильный орган определяет напряжение мышцы, которое изменяет собственное напряжение рецептора. аким образом, сухожильные рецепторы Гольджи обеспечивают нервную систему непрерывной информацией о степени напряжения любого небольшого сегмента каждой мышцы.
Когда рецепторы Гольджи мышечного сухожилия стимулируются при увеличении напряжения связанной с ними мышцы, сигналы передаются к спинному мозгу, вызывая рефлекторный ответ соответствующей мышцы. Этот рефлекс полностью тормозной. Он обеспечивает механизм отрицательной обратной связи, предупреждающий развитие слишком сильного напряжения мышцы.
В чем особенность миотатических рефлексов при раздражении первичных окончаний мышечных веретен и с чем она связана?
7.Верхние конечности человека в покое находятся в состоянии легкого сгибания в локтевом суставе. Чем объяснить данное явление?
При постоянном растяжении двуглавой мышци под действием силы тяжести происходит возбуждение мышечных рецепторов.
Соответственно в состоянии покоя происходит рефлекторное сокращение бицепса(волокна которого будут иметь за счет рефлекса растяжения повышенный тонус).
При растяжении мышци вместе в альфа-мотонейронами активируются гаммамотонейроны,которые иннервируют интрафузальные мышечные волокна.Они играют важную роль в поддержании тонуса мышц.
При одностороннем нарушении задних корешков ,иннервирующих верхние конечности, произойдет нарушение чувствитльности на как на поврежденной,так и на неповрежденной сторонах, а также паралич в связи с непоступлением импульсов на мотонейроны от вышележащих отделов.Произойдет разрыв положительной обратной связи.
8.Спинальное животное-это животное, у которого произведена перерезка спинного мозга с сохранением дыхания и устранением проявлений спинального шока. Какие особенности жизнедеятельности для него характерны?
1) Для сохранения дыхания перерезка спинного мозга (у спинального животного) осуществляется не выше 4-6 шейных сегментов
2) Спинальное животное не может сохранять позу и произвольно ее изменять.
3) Спинальное животное не может стоять, так как пересечены связи мотонейронов спинного мозга со структурами ствола, обеспечивающими тонические рефлексы — рефлексы, направленные на сохранение естественной позы.
4) Спинальное животное не способно к произвольным движениям.
5) Будут осуществляться спинальные рефлексы мочеиспускания, сгибания, разгибания, растяжения, ритмические рефлексы. Рефлексы, способствующие изгнанию каловых масс сохраняются, но не могут осуществляться, так как животное не может создать внутрибрюшное давление
9.Томографическое исследование показало очаг одностороннего повреждения структур заднего мозга. Какие нарушения чувствительности и двигательной функции можно наблюдать у больного
вобласти туловища и головы?
1) Задний мозг включает продолговатый мозг и варолиев мост.
2) В задний мозг поступают афферентные сигналы от рецепторов кожи и мышц головы, вестибулярных и слуховых рецепторов, вкусовых рецепторов, рецепторов внутренностей.