

kossitsky
.pdfДля удобства описания и изучения каждый цикл ходьбы подразделяют на периоды: двухопорный период, когда обе ноги стоят на опоре; одноопорный период для правой ноги и переносный для левой, снова двухопорный и затем одноопорный для левой ноги и переносный для правой. Во время опорной фазы происходит перекатывание стопы с пятки на носок. В опорных реакциях ходьбы выделяют передний и задний толчки, первый — при переносе нагрузки на опорную ногу, второй — при отталкивании ноги от опоры. В осуществлении ходьбы участвуют мышцы стопы, голени, бедра и тазового пояса (рис. 99).
Сопоставление изменений межзвенных углов в тазобедренном, коленном и голеностопном суставах с распределением во времени активности мышц показывает, что кинематика ходьбы является результатом взаимодействия мышечных и немышечных сил. Так, задний толчок, т. е. отталкивание стопы от опоры в результате подошвенного сгибания стопы, осуществляется напряжением задней группы мышц голени, а опускание стопы после соприкосновения пятки с опорой — под влиянием силы тяжести. Определенные фазы движения в коленном суставе также происходят под влиянием немышечных сил: сгибание в начале фазы переноса осуществляется по инерции в результате заднего толчка, последующее разгибание — сначала вследствие силы тяжести, а затем под влиянием инерции в результате активного движения бедра.
Анализ работы мышц при ходьбе показывает, что в разные фазы шага они сокращаются в разных режимах — концентрическом, т. е. с укорочением (например, мышцы, осуществляющие подошвенное сгибание стопы в опорном периоде), эксцентрическом,
151
т. e. с удлинением (например, сгибатели коленного сустава, тормозящие разгибание
вконце переносного периода), изометрическом, т. е. без изменения длины (мышцы тазобедренного сустава во время переката через пятку). В последнем случае наблюдается одновременное напряжение мышц-антагонистов, и их функциональная задача при этом заключается в фиксации сустава. В некоторые фазы ходьбы движение имеет баллистический характер, т. е. соответствующая мышца активна лишь в начале перемещения звена, а затем движение продолжается по инерции.
Повторяемость параметров движений в последовательных циклах при ходьбе не абсолютная: они обладают некоторой вариативностью. Наименьшая вариативность у кинематической картины ходьбы, наибольшая — в работе мышц, проявляющаяся в вариативности электромиограмм. Это отражает корригирующую деятельность ЦНС, которая
вкаждом шаге вносит в стандартную иннервационную структуру ходьбы поправки, необходимые для обеспечения относительного постоянства ее кинематики.
Бег отличается от ходьбы тем, что нога, которая находится позади, отталкивается
от опоры раньше, чем другая нога опускается на нее. В результате в беге имеется безопорный Период — период полета. В беге благодаря большим, чем при ходьбе, скоростям перемещения более значительную роль играют баллистические компоненты движения — перемещение звеньев ноги по инерции.
РАБОЧИЕ ДВИЖЕНИЯ
Рабочими движениями в широком смысле слова могут быть названы самые различные целенаправленные движения и действия, совершаемые как в процессе труда, так и
вповседневной жизни.
Учеловека основным рабочим органом является рука, причем для выполнения дви-
гательной задачи обычно наиболее важно положение кисти, которая в результате движения должна в определенный момент оказываться в определенном месте пространства. Благодаря большому количеству степеней свободы верхней конечности кисть может попасть в нужную точку по разным траекториям и при различных соотношениях углов в плечевом, локтевом и лучёзапястном суставах. Это многообразие возможностей позволяет выполнять двигательную задачу начиная движение из различных исходных поз. ' Электромиографическое исследование ряда рабочих движений показало сложную картину работы мышц, однако в этой картине часто можно выделить устойчивые сочетания активности нескольких мышц, используемые в различных движениях. Это так называемые Синергии, основанные на врожденных или выработанных в процессе двигательного опыта связях, которые, являясь устойчивыми компонентами движений, упрощают управление сложными двигательными актами.
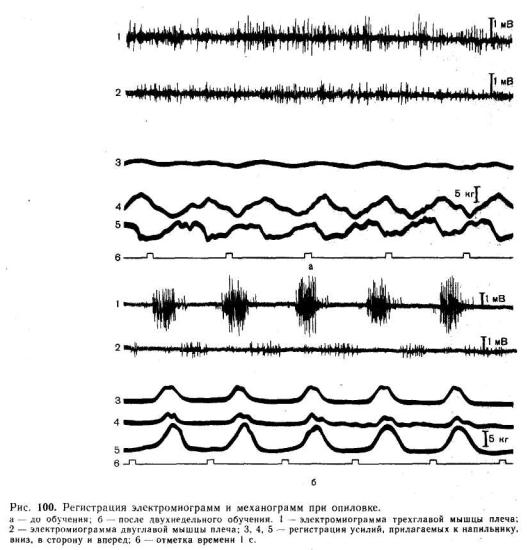
При совершении одного и того же, даже простого движения организация мышечной Деятельности в сильной степени зависит от вмешательства немышечных сил, в частности внешних по отношению к человеку. Так, при ударе молотком, когда к массе предплечья добавляется масса молотка и, следовательно, возрастает роль инерции, разгибание предплечья совершается по типу баллистического движения — мышцы-раз-
гйбатёли активны только в начале разгибания, которое дальше совершается по инерции, а в конце притормаживается мышцами-антагонистами. Аналогичное по кинематике движение При работе напильником, когда основная внешняя сила — трение, совершается путем непрерывной активности мышцы на протяжении всего разгибания (рис. 100). Если в первом из этих Двух движений преобладает роль программы, то во втором велика роль обратных связей.
ПОЗА
У млекопитающих животных и человека поддержание позы обеспечивается тем же механизмом сокращения мышц, что и движение. Отличие заключается в том, что при «позной» деятельности мышц сила их сокращения обычно невелика, режим близок к
152
изометрическому, а длительность сокращения значительна, В «позной» деятельности мышц участвуют преимущественно низкопороговые, медленные, устойчивые к утомлению двигательные единицы.
Поза и движение не существуют самостоятельно, поскольку движение всегда происходит в условиях некоторой позы, а смена поз осуществляется путем движения.
Одна из основных задач «позной» деятельности мышц — удержание в нужном положении суставов и звеньев тела в поле силы тяжести (удержание головы от свисания, голеностопных суставов от тыльного сгибания при стоянии и др.). Кроме того, «познан» активность может быть направлена на фиксацию суставов, не принимающих участия в данном движении. В трудовой деятельности удержание позы бывает связано с преодолением внешних сил.
Типичный пример позы — стояние человека. Сохранение равновесия при стоянии возможно в том случае, если проекция центра тяжести тела находится в пределах площади, занимаемой на плоскости опоры стопами (рис. 101). При стоянии вертикаль, иду-
153
щая через общий центр тяжести тела, проходит несколько впереди оси голеностопных и коленных суставов и несколько позади оси тазобедренных суставов. Следовательно, на эти суставы действуют моменты сил тяжести вышерасположенных звеньев тела, а это делает необходимым для удержания позы стояния напряжение многих мышц туловища и ног. Активность этих мышц невелика. Некоторые из них, например прямая мышца живота и двуглавая мышца бедра, поддерживают постоянную активность, другие, например мышцы голени, активируются периодически. Последнее связано с небольшими колебаниями центра тяжести тела, постоянно происходящими при стоянии как в сагиттальной, так и во фронтальной плоскости. Эти колебания могут быть зарегистрированы специальным прибором — стабилографом, работающим с помощью
тензодатчиков (рис. 102). Мышцы голени противодействуют отклонениям тела, возвращая его в вертикальное положение (рис. 103). Таким образом, поддержание позы
— это активный процесс, осуществляющийся, как и движение, с участием обратных связей от рецепторов. Среди последних в поддержании позы важную роль играет вестибулярный аппарат.
Поза стояния у человека энергетически относительно экономна, так как моменты силы тяжести невелики вследствие близости проекции центра тяжести тела от осей основных суставов ног. Менее экономно стояние у многих животных, например, у кошки и собаки, которые стоят на полусогнутых конечностях.
Поддержание постоянной позы — только частный случай «позной» активности мышц. Обычно в процессе двигательной деятельности происходит смена поз, так как движение связано с изменением положения центра тяжести тела и его звеньев. Сохранение устойчивости при движениях обеспечивается тем, что перераспределение «позной» активности мышц обычно предшествует движению.
Кпонятию позы примыкает понятие мышечного тонуса. Термин «тонус» многозначен,
вприменении к скелетным мышцам им обозначают комплекс явлений. В покое мышечные волокна обладают тургором, определяющим их сопротивление давлению и растяжению. Это составляет тот компонент тонуса, который не связан со специфической нервной активацией мышцы, обусловливающей ее сокращение. Однако в естественных условиях большинство мышц обычно в некоторой степени активируются нервной системой, в частности, для поддержания позы («позный тонус»). Полное расслабление мышцы, когда
электромиографическим методом с помощью как накожных, так и внутримышечных электродов в ней не регистрируется никаких потенциалов действия, достигается только

в условиях полного покоя и при исключении задачи поддержания позы в поле силы тяжести (исследуемая часть тела лежит на опоре).
Другой важный компонент тонуса — рефлекторный, который определяется рефлексом на растяжение. При исследовании человека он выявляется по сопротивлению растяжению мышцы при пассивном повороте звена конечности в суставе. Этот прием широко используется в медицине. Если в процессе такого исследования записать электромиограмму, то в растягиваемой мышце регистрируется электрическая активность, свидетельствующая об активации двигательных единиц. У здорового человека рефлекс на растяжение путем пассивного движения наблюдается только в процессе самого растяжения и притом при достаточно большой скорости растяжения. Тонический компонент рефлекса на растяжение, т. е. активность в растянутой мышце, обычно отсутствует, о чем можно судить по отсутствию колебаний потенциала на электромиограмме.
Нарушения тонуса при ряде заболеваний проявляются в увеличении ответа на растяжение, снижении порога скорости, появлении тонического компонента тонуса (мышечная гипертония). При некоторых заболеваниях наблюдается, наоборот, снижение рефлекторного тонуса (гипотония).
ВЫРАБОТКА ДВИГАТЕЛЬНЫХ НАВЫКОВ
Совершенствование двигательной функции человека в процессе онтогенеза происходит как вследствие продолжающегося в первые годы после рождения созревания отделов нервной системы и врожденных механизмов, участвующих в координации движений, так и в результате обучения, т. е. формирования новых связей, ложащихся в основу программ тех или иных конкретных двигательных актов. Координация новых, непривычных движений имеет характерные черты, отличающие ее от координации тех же движений после обучения.
Выше уже говорилось, что обилие степеней свободы в опорно-дигательном аппарате, влияние на результат движения сил тяжести и инерции осложняют выполнение любой двигательной задачи. Связанные с этим трудности особенно влияют на выполнение новых движений. На первых этапах обучения двигательный аппарат справляется с этими трудностями, нейтрализуя помехи путем развития дополнительных мышечных напряжений. Мышечный аппарат жестко фиксирует суставы, не участвующие в движении, активно тормозит инерцию быстрых движений. Такой путь преодоления помех, возникающих в ходе движения, энергетически невыгоден и утомителен. Использование обратных связей еще несовершенно — коррекционные посылки, возникающие на их основе, несоразмерны и вызывают необходимость новых коррекций.
На электромиограммах видно, что мышцы-антагонисты даже тех суставов, в которых совершается движение, активируются одновременно, в циклических движениях мышцы почти не расслабляются. Возбуждены также многие мышцы, не имеющие прямого отношения к данному двигательному акту. Движения, совершаемые в таких условиях, напряжены и неэстетичны (например, движения человека, впервые вышедшего на.коньках на лед).
155
Как показал в своих исследованиях Н. А. Бернштейн, по мере обучения вырабатывается такая структура двигательного акта, при которой немышечные силы включаются в его динамику, становятся составной частью движения. Излишние мышечные напряжения при этом устраняются, движение 'становится более устойчивым против помех. На электромиограммах видна концентрация возбуждения мышц во времени и пространстве: периоды активности работающих мышц укорачиваются, а количество мышц, вовлеченных в возбуждение, уменьшается. Это приводит к экономизации мышечной деятельности, а движения делаются более плавными, точными и непринужденными.
Ведущую роль в обучении движениям играет рецепция, особенно проприорецепция. В процессе этого обучения обратные связи используются не только для коррекции движе-
ния по его ходу, но и для коррекции программы следующего движения на основе ошибок предыдущего.
УТОМЛЕНИЕ
При длительной физической работе наступает утомление, которое, в частности, проявляется в изменении координации мышечной деятельности. Возбуждение каждой работающей мышцы становится менее концентрированным во времени. В работу вовлекаются другие мышцы, сначала синергисты, компенсирующие снижение силы основных мышц, а затем, по мере нарастания дискоординации — и другие мышцы, в частности антагонисты. Движения становятся все менее точными, темп их замедляется.
Картина мышечной активности во время движений, совершаемых на фоне утомления, во многом напоминает картину, наблюдаемую при выполнении новых, непривычных движений.
КРОВОСНАБЖЕНИЕ МОЗГА И ЛИКВОР
КРОВОСНАБЖЕНИЕ МОЗГА
Кровоснабжение головного мозга обеспечивается двумя сонными и двумя позвоночными артериями, которые образуют артериальный круг большого мозга (виллизиев круг); от него отходят артериальные ветви, питающие мозговую ткань.
Непрерывное снабжение головного мозга кровью — основное условие его нормальной деятельности. Никакие другие клетки так быстро, как нервные, не перестают функционировать при прекращении или резком уменьшении кровоснабжения; временное обескровливание мозга приводит к обморочному состоянию. Причиной такой чувствительности мозга к кровоснабжению является большая потребность его в кислороде и питательных веществах, в частности в глюкозе.
При усиленной работе коры больших полушарий головного мозга, в частности при решении арифметических задач, чтении и т. д.,увеличивается ее кровоснабжение вследствие расширения мозговых сосудов. Это наблюдалось у людей, у которых в результате перенесенной травмы имелось отверстие в костях черепа, сквозь которое можно было регистрировать пульсацию мозга и его кровенаполнение. Это же отмечалось с помощью современных электронных приборов (реоплетизмографы и др.), позволяющих исследовать кровоснабжение мозга без повреждения черепа. В хронических опытах на животных можно наблюдать кровообращение мозга непосредственно, заменив естественную крышку черепа на искусственную прозрачную — из пластмассы.
ЛИКВОР
Внутри мозга и под его оболочками имеются большие пространства, заполненные спинномозговой жидкостью, или ликвором (Liquor cerebrospinalis). Ликвор заполняет мозговые желудочки, центральный канал спинного мозга и подпаутинное пространство (cisterna subarachnoidalis) как головного, так и спинного мозга.
156
Желудочковый и подпаутинный ликвор сообщаются между собой посредством отверстий: непарного foramen Luschka и парных foramina Mageinlie, расположенных в области продолговатого мозга.
Количество ликвора у человека равно 120;—150 мл, причем большая его часть приходится на подпаутинные пространства и только 20—40 мл содержится в желудочках.
Ликвор прозрачен и бесцветен. Удельный вес его 1,005—1,007; реакция слабо щелочная и рН близок к рН крови (7,4). В ликворе содержится очень небольшое количество клеток — лимфоцитов (1—5 мкл). Ликвор по своему составу отличается от крови и лимфы меньшим содержанием белков (содержание белков в крови около 7—8%, в лимфе — 0,3—0,5%, а в ликворе только 0,02%). Содержание неорганических солей в ликворе примерно такое же, как в плазме крови. В ликворе отсутствуют ферменты и иммунные тела.
Ряд данных говорит о том, что образование ликвора происходит в результате активной секреции клеток, выстилающих сосудистое сплетение желудочков мозга.
Так как количество ликвора более или менее постоянно, а его образование происходит непрерывно, следовательно, непрерывным должно быть и его всасывание. Отток ликвора происходит главным образом в венозную систему из подпаутинных пространств.
Ликвор — внутренняя среда мозга, поддерживающая его солевой состав и осмотическое давление. Кроме того, ликвор является гидравлической подушкой мозга, создающей надежную механическую защиту нервных клеток.
Нарушение циркуляции ликвора ведет к расстройствам деятельности ЦНС. Значение ликвора состоит в том, что он в известной мере является'питательной средой мозга. На это указывает большое содержание сахара в ликворе желудочков (где ликвор образуется) по сравнению с ликвором подпаутинного пространства 1где он всасывается). Значение ликвора заключается еще и в том, что с его током удаляются из мозга и поступают в кровь продукты распада, образующиеся в процессе обмена веществ тканей мозга.
ГЕМАТОЭНЦЕФАЛИЧЕСКИЙ БАРЬЕР
Состав ликвора в большей степени зависит от свойств так называемого гематоэнцефалического барьера (Л. С. Штерн). Таким барьером являются стенки капилляров, разделяющие кровь и ликвор, а также, возможно, некоторые нейраглиальные клетки — астроциты. О наличии этого барьера свидетельствует ряд фактов, в частности различие в составе крови и ликвора. Многие вещества, находящиеся в крови или искусственно вводимые в кровь, совершенно отсутствуют в ликворе, между тем как другие вещества, близкие к первым по своему химическому строению, содержатся как в крови, так и в ликворе в равных или почти равных концентрациях Это свидетельствует о том, что мембраны, разделяющие кровь и ликвор, обладают избирательной проницаемостью.
Внорме из крови в ликвор не проходят и, следовательно, задерживаются барьером соединения йода, соли азотной кислоты, соли салициловой кислоты, метиленовая синь, все коллоиды, иммунные тела, антибиотики (пенициллин и стрептомицин). Легко проходят через барьер в ликвор алкоголь, хлороформ, стрихнин, морфин, столбнячный токсин (этим, по всей вероятности, объясняется быстрое действие на нервную систему этих веществ после их поступления в кровь).
Всвязи с тем что многие лекарственные или биологически активные вещества не про-
ходят в ликвор и потому не оказывают влияния на нервные центры, Л. С. Штерн рекомендовала вводить их непосредственно в ликвор, минуя гематоэнцефалический барьер. Для этого производят субокципитальный или люмбальный прокол и вводят лекарственные вещества непосредственно в ликвор.
Этот способ введения лекарственных веществ использовали при лечении некоторых инфекционных заболеваний мозга в том случае, если лекарственные средства, например антибиотики, не проходили через гематоэнцефалический барьер, а также при необходимости непосредственного воздействия некоторых химических веществ на ЦНС для изменения ее деятельности.
157
Характер действия на организм некоторых веществ при их введении в кровь или ликвор может быть совершенно различен. Если в кровь вводится вещество, которое не проникает через гематоэнцефалический барьер, то реакция организма на его введение будет зависеть только от того, как на введенное вещество реагируют периферические органы. Если же это вещество вводится непосредственно в ликвор, реакция на его введение в первую очередь зависит от действия вещества на нервные центры. Этим объясняется, например, следующий факт: введение АТФ в кровь влечет за собой падение артериального давления вследствие расширения артерий и артериол: при введении же этого вещества непосредственно в ликвор субокципитальным проколом происходит повышение артериального давления в результате возбуждающего действия АТФ на сосудодвигательный центр продолговатого мозга. В последнее время получены данные, свидетельствующие о возможности естественной регуляции функционального состояния мозга путем изменения состава ликвора. Биологически активные вещества, действующие на нервные и глиальные клетки ЦНС, могут выделяться самими нервными клетками (нейросекреция) либо поступать из крови через гематоэнцефалический барьер. Как показала Л. С. Штерн, избирательное изменение проницаемости гематоэнцефалического барьера для некоторых биологически активных веществ, имеющихся в крови, может быть способом регуляции функционального состояния мозга. Так как проницаемость гематоэнцефалического барьера регулируется ЦНС, то в данном случае имеет место один из способов, с помощью которых мозг может регулировать собственное функциональное состояние.
Гл а в а 7
НЕРВНАЯ РЕГУЛЯЦИЯ ВЕГЕТАТИВНЫХ ФУНКЦИЙ
Со времени французского физиолога Биша — с начала XIX столетия — функции ор-
ганизма разделяют на анимальные, или соматические, и вегетативные. К анимальным,
или соматическим, функциям относятся восприятие внешних раздражений и двигательные реакции, осуществляемые скелетной мускулатурой. Вегетативными функциями называют те, от которых зависит осуществление обмена веществ в целостном организме (пищеварение, кровообращение, дыхание, выделение и т, д.), а также рост и размножение. В соответствии с этим разделением функций различают соматическую и вегетативную нервную систему. Соматическая нервная система обеспечивает экстероцептивные сенсорные и моторные функции организма. Вегетативная нервная система обеспечивает регуляцию деятельности внутренних органов, сосудов и потовых желез, а также трофическую иннервацию скелетной мускулатуры, рецепторов и самой нервной системы.
Вегетативная нервная система отличается от соматической локализацией своих ядер в ЦНС, очаговым выходом волокон из мозга, отсутствием сегментарности их распределения на периферии и малым диаметром волокон. Помимо этого, для вегетативной нервной системы характерно, что ее волокна, направляющиеся из мозга к внутренним органам, обязательно прерываются в периферических вегетативных ганглиях, образуя синапсы на нейронах, расположенных в этих ганглиях. Аксоны ганглионарных нейронов оказывают влияние на внутренние органы.
Ганглии являются не только образованиями, передающими импульсы из ЦНС на органы и ткани. Они представляют собой вынесенные на периферию рефлекторные центры, способные регулировать функции внутренних органов в известной мере независимо от ЦНС посредством периферических рефлексов, замыкающихся в этих ганглиях.
Вегетативная нервная система делится на симпатический и парасимпатический отделы. Они отличаются по локализации центров в мозге, по характеру влияний на внутренние органы, а также тем, что ганглии парасимпатического отдела расположены в самих иннервируемых ими органах (интрамурально), в то время как ганглии симпатического отдела локализованы в пограничном стволе (truncus sympaticus).
158
Выделяя регуляцию вегетативных функций, надо отметить, что в целостных реакциях организма сенсорные, моторные и вегетативные компоненты тесно связаны между собой.
Соматические компоненты реакций организма, осуществляемые скелетной мускулатурой, в отличие от вегетативных могут быть произвольно вызваны, усилены или заторможены; они находятся в течение всего хода реакции под контролем сознания.
Вегетативные же компоненты, как правило, произвольно не контролируются. На этом основании вегетативную нервную систему называют автономной, или непроизвольной. Однако представление об автономности вегетативной нервной системы является весьма условным.
Многочисленные опыты К. М. Быкова и сотрудников, показавшие возможность условнорефлекторной регуляции всех внутренних органов и всех вегетативных функций, позволяют считать, что кора больших полушарий регулирует деятельность всех органов, иннервированных вегетативной нервной системой, и координирует их деятельность в соответствии с текущими потребностями организма, в зависимости от характера его реакций при изменениях внешней и внутренней среды организма. Это обстоятельство свидетельствует о принципиальной возможности произвольного управления вегетативными функциями, что удается осуществить после специальной целенаправленной тренировки (например, по системе индийских йогов).
ОБЩИЙ ПЛАН СТРОЕНИЯ И ОСНОВНЫЕ ФИЗИОЛОГИЧЕСКИЕ СВОЙСТВА
ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
Центры вегетативной нервной системы расположены в мозговом стволе и спинном мозге. 1. В среднем мозге находятся мезэнцефальные центры парасимпатического отдела вегетативной нервной системы; вегетативные волокна от них идут в составе глазодвигательного нерва. 2. В продолговатом мозге расположены бульварные центры парасимпатического отдела нервной системы; эфферентные волокна от них проходят в составе лицевого, языкоглоточного и блуждающего нервов. 3. В грудных и поясничных сегментах-спин- ного мозга (от I грудного до II — IV поясничного) находятся тораколюмбальные центры симпатического отдела вегетативной нервной системы: вегетативные волокна от них выходят через передние корешки спинномозговых сегментов вместе с отростками моторных нейронов. 4. В крестцовых сегментах спинного мозга находятся сакральные центры парасимпатического отдела вегетативной нервной системы, волокна от них идут в составе тазовых нервов.
Таким образом, центры вегетативной нервной системы расположены в четырех отделах ЦНС. Ядра, находящиеся в мезэнцефальном, бульварном и сакральном отделах, образуют парасимпатическую часть вегетативной нервной системы, а находящиеся в тораколюмбальном отделе — ее симпатическую часть.
Все уровни вегетативной нервной системы подчинены высшим вегетативным центрам, расположенным в промежуточном мозге — в гипоталамусе и полосатом теле. Эти центры координируют функции многих органов и систем организма. Они в свою очередь подчинены коре больших полушарий, которая обеспечивает целостное реагирование организма, объединяя его соматические и вегетативные функции в единые акты поведения.
Симпатические нервные волокна имеют значительно более широкое распространение, чем парасимпатические. Симпатические нервы иннервируют фактически все органы и ткани организма; напротив, парасимпатические же нервы не иннервируют скелетную мускулатуру, ЦНС, большую часть кровеносных сосудов и матку.
Распространение волокон парасимпатического отдела вегетативной нервной системы представлено на рис. 104. Ко многим органам парасимпатические волокна проходят в составе блуждающих нервов, которые иннервируют бронхи, сердце, пищевод, желудок, печень, тонкий кишечник, поджелудочную железу, надпочечники, почки, селезенку, часть толстого отдела кишечника.
159

Распространение волокон симпатического отдела вегетативной нервной системы показано на рис. 105. Верхние сегменты симпатического отдела вегетативной нервной системы посылают свои волокна через верхний шейный симпатический узел к органам головы; следующие сегменты посылают их через нижележащие симпатические узлы к органам грудной полости и верхним конечностям; далее следует ряд грудных сегментов, посылающих волокна через солнечное сплетение и верхний брыжеечный узел к органам брюшной полости,- и, наконец, от поясничных сегментов волокна направляются через нижний брыжеечный узел в основном к органам малого таза и нижним конечностям.
ДВУХНЕЙРОННАЯСТРУКТУРАЭФФЕРЕНТНЫХ СИМПАТИЧЕСКИХИПАРАСИМПАТИЧЕСКИХПУТЕЙ
Периферическая часть всех эфферентных симпатических и парасимпатических нервных путей построена из двух последовательно расположенных нейронов. Тело первого нейрона находится в ЦНС, его аксон направляется на периферию и оканчивается в том или ином нервном узле. Здесь находится тело второго нейрона, на котором аксон первого нейрона образует синаптические окончания. Аксон второго нейрона иннервирует соот-
160