

ЭЛЕКТРИЧЕСКИЕ
ПРОЯВЛЕНИЯ АКТИВНОСТИ ПУТЕЙ И ЦЕНТРОВ
СЛУХОВОЙ СИСТЕМЫ
Общая
задача современных электрофизиологических
исследований деятельности слуховой
системы может быть сформулирована
как изучение закономерностей описания
параметров звуковых сигналов в
активности нервных элементов, составляющих
слуховую систему. Сопоставление
особенностей такого описания в основных
отделах слуховой системы, начиная от
волокон слухового нерва и кончая
слуховой областью коры, позволяет
подойти к изучению тех закономерностей
обработки информации о сигнале, которые
обеспечивают организму возможность
обнаружения и классификации всего
многообразия звуков внешней среды.
Электрофизиологическое
исследование путей и центров слуховой
системы осуществляется в настоящее
время двумя основными методами. Один
из этих методов состоит в регистрации
и последующей оценке активности
больших совокупностей нервных элементов,
реакции которых получили в литературе
ряд обозначений, употребляющихся
(не вполне строго) как синонимы: суммарные
реакции или ответы, вызванные потенциалы,
первичные ответы. Эти обозначения
характеризуютраз- личные свойства
указанного класса биоэлектрических
явлений: участие суммы нервных элементов
в их генерации, обусловленность их
появления воздействием звукового
стимула, раннее время их появления и
развития по сравнению с другими, более
поздними (например, вторичными) ответами.1
Второй метод состоит в регистрации
активности отдельных нервных элементов
(волокон или клеток) с помощью
микроэлектродов. При микроэлектродном
1
В понятие «суммарные вызванные ответы»
входят также суммарные реакции,
возникающие и протекающие в более
поздние или более длительные периоды
времени по сравнению с первичными
ответами (вторичные реакции,
сдвиги,постоянного потенциала); подробнее
см.: Tsuchitani, Boudreau, 1964; David et al., 1969; Ройтбак,
1969; Радионова, 1971, и др.
210Глава 7
методе
исследования деятельности нервных
элементов слуховой системы в подавляющем
большинстве работ используется
внеклеточное отведение: относительно
малый размер клеток слуховой системы
создает большие методические трудности
для внутриклеточной регистрации
активности, поэтому такие работы
являются единичными (Nelson, Erulkar, 1963;
Gerstein et al., 1968; Starr, Britt, 1970). Следует также
отметить, что исследование как суммарной
активности, так и активности одиночных
нейронов слуховой системы, как
правило, проводится с использованием
адекватной для слуховой системы
стимуляции, т. е. стимуляции звуковыми
сигналами. Вместе с тем метод электрического
раздражения, занимающий важное место
в других областях электрофизиологии,
при изучении электрических реакций
слуховой системы использовался лишь
в очень ограниченном числе работ,
основные из которых упоминаются в
последующем изложении. Работы, выполненные
с применением метода электрического
раздражения при исследовании слуха
человека, выходят за рамки настоящего
раздела и рассмотрены отдельно на
стр. 280.
КРАТКАЯ
ХАРАКТЕРИСТИКА СУММАРНЫХ ЭЛЕКТРИЧЕСКИХ
ОТВЕТОВ
Электрофизиологическое
исследование реакций путей и центров
слуховой системы на звуковые воздействия
было начато в 30-х годах с изучения
суммарных ответов, возникающих при
действии звуковых сигналов в основных
отделах слуховой системы (Derbyshire, Davis,
1935; Kemp et al., 1937; Гершуни, 1940). В дальнейшем
были подробно описаны основные
характеристики суммарных ответов всех
отделов слухового пути (рис. 68, 69, табл.
6, 7). Необходимо подчеркнуть, что данные,
приведенные на рис. 68, 69 и табл. 6, 7, по
существу не вполне однозначны, для
проводниковых (слуховой нерв, латеральная
петля) и клеточных отделов слуховой
системы (кохлеарные ядра, верхняя олива,
задние холмы, внутреннее коленчатое
тело, слуховая область коры больших
полушарий). В первом случае речь идет
о суммарной импульсной реакции
волокон, во втором же — о суммарных
первичных вызванных ответах слуховых
ядер, в которых, по-видимому, могут
находить отражение как синаптические,
так и импульсные реакции совокупности
нейронов (Bishop, McLeod, 1954; Jungert, 1958; Альтман
и др., 1963; Вартанян, 1966а; Серков, 1970, и
др.).
Исследование
суммарных ответов позволяет изучить
целый ряд существенных сторон в
организации деятельности слуховой
системы. Прежде всего были установлены
те области мозга, в которых могут быть
зарегистрированы суммарные ответы при
действии звука. Были определены фокусы
максимальной
14*
211
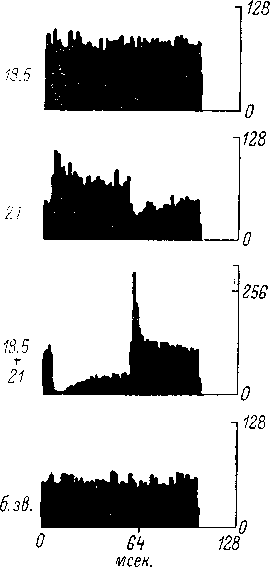
Рис.
68. Суммарные вызванные ответы различных
отделов слу-
ховой системы, возникающие
при действии звукового щелчка
(ориг.;
данные для верхней оливы — реконструкция
по: Galam-
bos et al., 1959).
CH
—
слуховой нерв (регистрация от круглого
окна улитки); НЯ
—
кохлеарные ядра; ВО
—
комплекс верхней оливы; ЛП
—
латеральная петля; ЗХ
—
задний холм; ВКТ — внутреннее коленчатое
тело; СН—
слуховая кора (зона AI, поверхностное
отведение). Точкой
отмечены компоненты суммарного ответа,
усредненные характеристики которых
приведены в табл. 6, 7. Нижняя
осциллограмма для СН,
полученная в условиях хлоралозного
наркоза, демонстрирует, помимо первичного,
также и поздние компоненты суммарного
ответа. Калибровка амплитуды ответов:
100 мкв (СН,
НЯ, ВО)
и 25 мкв (ЛП,
ЗХ, ВНТ, СН).
Калибровка времени: 5 мсек. (левый
столбец)
и 20 мсек, (правый
столбец и СК, внизу).
Негативность вверх. Полоса пропускания
усилителя: 10 гц—10 кгц (СН,
НЯ)и
10—1000 гц (ЛП,
ЗХ, ВНТ, СН).
Момент подачи щелчка отмечен импульсом
в случаях СН,
НЯ, ЛП, ЗХ, ВНТ, СН
или соответствует началу развертки
луча в случае ВО.


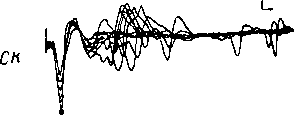
активности
различных отделов слуховой системы в
зависимости от частоты звукового
сигнала и стороны, с которой он
предъявляется. В частности,
характеристики суммарных ответов при
сопоставлении с морфологическими
данными позволили в общих чертах
определить характер конвергенции
билатерального потока импульсации на
разных уровнях слуховой системы
(Rosenzweig, Rosenblith, 1953), а также выделить ряд
самостоятельных функциональных зон
в слуховой области коры (Woolsey, 1961).
Рис.
69. Зависимость скрытых периодов (Л) и
амплитуд (В)
вызванных
потенциалов разных отделов слуховой
системы от интенсивности звуковых
сигналов (щелчков) (по: Альтман,
Марусева, 1965).
1
—
улитка; 2
—
кохлеарное ядро; з
—
задний холм; 4
—
внутреннее
коленчатое тело; 5
—
слуховая зона коры. По
оси абсцисс —
интен-
сивность в дб; по
оси ординат'
на А — скрытый период в мсек., на Б
—
амплитуда ответа в мкв.
Регистрация
суммарных ответов позволяет также
измерить порог в том или ином отделе
слухового пути и оценить его изменения
при различных воздействиях. С помощью
метода регистрации суммарных ответов
было определено время распространения
возбуждения по ходу слуховой системы
(по величинам скрытых периодов реакций)
и его изменение при изменении интенсивности
стимула (рис. 69, А).
Были установлены также амплитудные
зависимости суммарных ответов от
интенсивности стимула (рис. 69, Б).
В
частности, оценка амплитудной зависимости
компонента Н1
суммарного
ответа слухового нерва позволила
предположить наличие двух групп
элементов, реагирующих в различных
диапазонах интенсивности стимула
(Радионова, 1971).
С
помощью суммарных ответов оценивается
ряд существенных временных характеристик
в деятельности слуховой системы. Так,
длительность суммарного ответа может
являться свидетельст-
213
Таблица
6
Количественные
характеристики суммарных ответов*
основных отделов слуховой системы**
при
действии звукового щелчка |
Порог обнаружения ответа в дб относительно порога слышимости человека |
Скрытый период в мсек. |
Максимальная амплитуда в МКВ |
Длительность в мсек. |
|
при пороговой интенсивности |
при максимальной интенсивности |
||||
Слуховой нерв |
—5 |
1.2—1.4 |
0.45—0.5 |
,2000 |
0.5 |
Кохлеарные ядра |
-5 |
3.2—3.6 |
2.0—2.4 |
600 |
3-5 |
Комплекс верхней оливы |
Не иссл. |
Не иссл. |
3 4*** |
300—500 |
4—6 |
Латеральная петля |
Не иссл. |
3.0-3.5 |
2.0-2.5 |
Не иссл. |
2.5—3.0 |
Задние холмы |
—2 |
4.8-6.3 |
3.2—3.6 |
450 |
4.5-5.5 |
Внутреннее коленчатое тело . . . |
-6 |
8.0 |
5.0 |
200-300 |
5-6 |
Слуховая зона коры |
—2 |
10.5—12.0 |
8.0-8.5 |
200—350 |
10 |
*
Компоненты суммарных электрических
ответов, для которых измерялись
представленные в табл. 6 и 7 количественные
характеристики, отмечены точками
на рис. 68. Эти компоненты, согласно
существующим представлениям, отражают
либо суммарную актив; ность волокон
(слуховой нерв, латеральная петля), либо
постсинаптическую реакцию клеточных
элементов остальных отделов слуховой
системы (см., например: Jungert, 1958; Galambos et
al., 1959; Гершуни, 1962; Альтман и др., 1963;
Альтман, Лебедева, 1965; Альтман, Ма-
русева, 1965; Вартанян, Ратникова, 1967;
Ройтбак, 1969; Серков, 1970; Радионова, 1971).
**
Данные по комплексу верхней оливы взяты
из работы на кошке (Galambos et al., 1959), остальные
данные взяты из работы Я. А. Альтмана,
А. М. Марусевой (1965) на кошке, за исключением
данных для латеральной петли, полученных
на крысе (Вартанян, 1966а).
***
Скрытый период при средней интенсивности
раздражения.
Таблица
7
Характеристики
суммарных ответов слуховой системы
в связи с временными параметрами
звуковых сигналов |
Время восстановления в мсек, между щелчками |
Предел воспроизведения ритма раздражения в гц |
Суммационные свойства*** |
Критическая постоянная времени сигнала (предел исчезновения суммарного ответа, мсек). |
||
появление отдельного ответа на второй щелчок |
полное восстановление |
глубина суммации в дб |
критическое время суммации в мсек. |
|||
Слуховой нерв |
1.4 (Альтман, Марусева, 1965). |
20—25 (Альтман, Марусева, 1965). |
3000—4000** (Гершуни, 1940; Peake et al., 1962). |
0 (Забоева, не- опубликовано). |
0 (Забоева, нс- опубликовано). |
10(Забоева, 1966). |
Кохлеарные ядра |
2 (Альтман и др., 1963). |
20—25* (Альтман и др. 1963). |
4000—5000 (Worden, Marsh, 1968; Радионова, 1971). |
2**** (Вартанян, 1966а). |
1.5—3**** (Вартанян, 1966а), |
160 (Альтман и др., 1963). |
Комплекс верхней оливы . . . |
— |
— |
4300 (Boudreau, 1965). |
— |
— |
— |
Латеральная петля**** .... |
— |
25—30 (Вартанян, 1966а). |
|
1.5 (Вартанян, 1966а). |
2 (Вартанян, 1966а), |
40 (Вартанян, 1971). |
«Задние холмы |
2.2—3 (Альтман, Марусева, 1965). |
40 (Альтман, Марусева, 1965). |
1200—1500 (Kemp et al., 1937; Гершуни, 1940). |
11**** (Вартанян, 19666). |
5—8**** (Вартанян, 19666), |
60—100**** (Вартанян, 1971). |
Внутреннее коленчатое тело . . |
10—14 (Альтман, Марусева, 1965). |
75—110* (Альтман, Марусева, 1965). |
1200—1500 (Гершуни, 1940). |
— |
— |
— |
Слуховая зона коры |
30—50 (Альтман, Марусева, 1965). |
400* (Альтман, Марусева, 1965). |
100(Гершуни, 1940; Goldstein et al., 1957). |
10—12 (Гершуни, 1963). |
8—10 (Гершуни, 1963). |
268 (Гершуни, Забоева, 1962). |
*
Восстановление до 85—90%, после чего
наблюдается цикличность процесса.
**
Эта цифра, по-видимому, занижена, о чем
свидетельствуют данные о синхронизации
импульсов одиночных волокон слухового
нерва с фазой звукового раздражения
до 5000 гц (см. «Слуховой нерв»).
***
Суммационные свойства оценивались при
изменении длительности звука от величин
порядка 0.5 до 100 мсек. Под глубиной
суммации понимается величина
изменения порога при увеличении
длительности звука. Критическое время
суммации характеризуется той максимальной
длительностью сигнала, которая еще
приводит к дальнейшему снижению порога.
****
Данные получены на крысе, остальные
данные получены на кошке.
вом
степени синхронизированности афферентного
потока на различных уровнях слуховой
системы. Как видно из рис. 68 и табл. 6,
длительность суммарных ответов
возрастает от нижележащих уровней
слуховой системы к вышележащим, что
может служить свидетельством
постепенной десинхронизации афферентного
потока импульсации. Уменьшение частоты
воспроизведения ритма стимуляции и
возрастание времени полного восстановления
ответа на второй стимул (при действии
парных звуковых сигналов с различными
интервалами между ними) может
рассматриваться как проявление понижения
разрешающей способности по времени от
нижележащих отделов слуховой системы
к вышележащим (табл. 7). Увеличение
способности к суммации (снижение порога
суммарного ответа при увеличении
длительности звукового сигнала) от
нижележащих отделов к вышележащим,
по-видимому, связано с возрастанием
длительности суммарных ответов, т. е.
с десинхронизацией афферентного потока
при его прохождении по ходу слухового
пути. Использование сигналов с разным
временем нарастания может явиться
примером исследования степени
устойчивости афферентного потока
импульсации на тех или иных уровнях
слухового пути к десинхронизирующему
воздействию, каким является увеличение
пологости фронта сигнала. Как показывают
данные табл. 7, по мере повышения уровня
слухового пути, требуется значительно
большее время нарастания стимула, чтобы
вызвать полную десинхронизацию
(исчезновение) суммарного ответа. Эти
факты могут служить доказательством
того, что несмотря на уменьшение
синхронизации начальной части
афферентного потока импульсации, в
вышележащих отделах слуховой системы
существуют механизмы, препятствующие
дальнейшим десинхронизирующим
влияниям (подробнее об этом см. стр.
297). Наконец, с помощью суммарных ответов
исследована специфическая сторона
деятельности слуховой системы —
тонотопическая локализация, т. е.
пространственное представительство
в центральных отделах слуховой
системы различных по частотной
настроенности областей улитки. Этот
материал подробнее рассмотрен в
последующих разделах изложения
(стр. 228, 240, 250). Функциональное значение
различных характеристик суммарных
ответов слуховой системы подробно
рассматривалось в ряде обзоров
(Rosenzweig, Rosen- blith, 1953; Гершуни, 1962; Альтман,
Марусева, 1965).
Однако
несмотря на изложенный выше достаточно
широкий круг вопросов, которые можно
исследовать с помощью суммарных ответов,
сам метод накладывает ряд ограничений
на его применение. Основными из них
являются: а) трудность трактовки
происхождения различных компонентов
суммарных ответов, а следовательно,
и трактовки обнаруживаемых изменений;
б) отражение в первичных суммарных
ответах лишь части афферентного потока
импульсации, возникающего главным
образом на включение (иногда выключение)
звукового сигнала; соотношение характе
216
ристик
более поздних (чем первичные) ответов
с параметрами звукового стимула до
настоящего времени остается
неисследованным. Микроэлектродный
метод исследования импульсной активности
одиночных нейронов слуховой системы
лишен части этих ограничений и делает
возможным более глубокий анализ
процессов, происходящих в слуховой
системе при действии звука. Однако
естественным ограничением этого метода
является невозможность одновременного
исследования достаточно большого числа
элементов. Таким образом, если суммарные
первичные вызванные ответы позволяют
судить о некоторых характеристиках
начальной части афферентного потока
одновременно в большой совокупности
нейронных каналов системы передачи
информации о звуке, то реакции одиночных
элементов дают возможность проследить
тонкую временную организацию работы
отдельных каналов этой системы. При
этом важно подчеркнуть, что в обоих
случаях исследователь имеет дело с
разными сторонами единого процесса,
поэтому регистрируемые явления
оказываются функционально взаимосвязанными.
ИМПУЛЬСНАЯ
АКТИВНОСТЬ НЕЙРОНОВ РАЗНЫХ ОТДЕЛОВ
СЛУХОВОЙ СИСТЕМЫ
Рассмотрение
характеристик импульсной активности
нейронов слуховой системы будет
проведено последовательно по основным
ее отделам от нижележащих к вышележащим
(слуховой нерв, кохлеарные ядра,
верхняя олива, задние холмы, внутреннее
коленчатое тело, слуховая область
коры.) При этом будет дано описание
следующих основных характеристик
импульсной активности нейронов
слуховой системы: спонтанной активности,
узора разряда в зависимости от
длительности и интенсивности
звукового сигнала, частотной
избирательности нейронов, тормозных
явлений, а также форм поведения нейронов,
либо обусловленных действием сигналов
с изменяющимися во времени параметрами
(амплитудно- и частотно-модулированные
сигналы), либо не связанных непосредственно
с параметрами стимула. Краткие сведения
об эфферентной регуляции импульсной
активности слуховой системы также
будут представлены ниже.
В
заключительной части главы будет
проведено сопоставление основных
характеристик электрической активности
нейронов разных уровней слуховой
системы — от слухового нерва до слуховой
зоны коры больший полушарий.
Слуховой
нерв
Как
указывалось, волокна слухового нерва
представляют собой проксимальные
отростки биполярных нейронов спирального
ганглия; они передают информацию от
улитки в первый центральный отдел
слуховой системы — кохлеарные ядра.
217
Как
показали микроэлектродные исследования,
практически
все волокна слухового
нерва у кошки обладают спонтанной
актив-
ностью, т. е. дают шмпульса-
п32
цию
в отсутствие f
звукового
раздражения. При этом
ча-
стота спонтанной активности,
как
правило, высока и в зна-
чительной
части случаев пре-
вышает 50, достигая
иногда
Рис.
71. Постстимульные гисто-
граммы
реакции волокон слухового
нерва
кошки на тон оптимальной
частоты
длительностью 250 мсек,
различной
интенсивности (по:
Kiang, 1965а).
Цифры
слева от гистограмм: —
дб зату-
хания относительно условного
исходного
уровня. Внизу
—
время в мсек., справа
—
количество
импульсов.
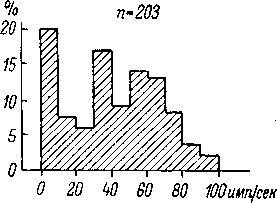
Рис.
70. Распределение волокон
слухового
нерва кошки (вверху,
по:
Kiang, 1965а) и обезьяны
(внизу,
по: Nomoto et al., 1964)
по уровню спонтанной
активности.
По
оси абсцисс —
частота спонтанной
импульсация в
имп./сек.; по
оси ор-
динат —
количество нейронов в про-
центах.
Наркоз: диал-уретан (кошки)
и
натрий-пентобарбитал (обезьяны).
п
—
число нейронов.
100
имп./сек. и более (Kiang, 1965а). У морской
свинки значительное число волокон,
реагирующих на звук, обнаруживают
218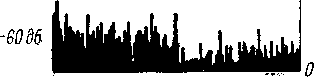
лишь
очень редкую, «случайную» спонтанную
импульсацию (Та- saki, 1954). У обезьяны
обнаружено довольно большое число
волокон, вообще не имеющих спонтанной
активности (Nomoto et al., 1964). Возможно, что
последнее обстоятельство связано с
характером применяемого в этом
случае барбитурового наркоза.
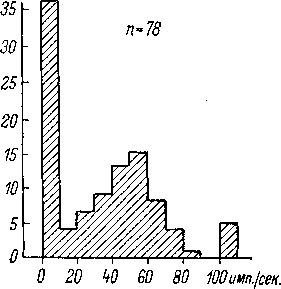
Распределение волокон слухового
нерва по уровню спонтанной активности
(рис. 70) показывает наличие двух основных
групп нейронов — с относительно
низким уровнем спонтанной активности
(до 10—20 имп. /сек.) и с более высоким
уровнем спонтанной активности. При
этом в общем сходный характер распределения
обнаруживается у разных представителей
млекопитающих (кошка, обезьяна, — рис.
70).
Временная
последовательность импульсов спонтанной
активности носит нерегулярный
характер, причем распределение
межимпульсных интервалов приближается
к пуассоновскому. Вопрос о происхождении
спонтанной активности в волокнах
слухового нерва представляет специальный
интерес. Уровень спонтанной активности
не связан ни с величиной порога реакции
на звук, ни с тем, какую область кортиева
органа (по его длине) данное волокно
иннервирует. Возможно, что спонтанная
активность волокон слухового нерва
обусловлена чувствительностью как
самих волокон, так и иннервируемых ими
волосковых клеток к различным воздействиям
случайной природы; однако этот вопрос
остается открытым.
Реакция
отдельного волокна на тональную или
шумовую посылку длительностью в
несколько десятков миллисекунд
или больше представляет пачку импульсов,
частота следования которых превышает
частоту спонтанной импульсации;
длительность такой реакции довольно
точно соответствует длительности
действующего стимула. Характерно,
что с течением времени действия стимула
частота импульсов в реакции на звук
уменьшается, достигая некоторого
стационарного уровня (процесс адаптации).
Этот процесс особенно отчетливо виден
при регистрации импульсной активности
в виде постстимульных гистограмм (рис.
71), которые статистически отражают
распределение импульсации во времени
от момента подачи звука. Длительность
процесса адаптации в волокнах
слухового нерва может быть различной
и в ряде случаев достигает нескольких
минут (Kiang, 1965а). Существенно отметить
также характерную реакцию волокон
слухового нерва на короткий (0.1 мсек.)
щелчок. В ответ на такой щелчок,
представляющий собой звуковой стимул
с широким спектром, волокно, отвечает
несколькими импульсами, интервал между
которыми равен периоду характеристичной
(оптимальной) частоты данного волокна
(рис. ЮЗ), если эта частота не превышает
4—5 кгц (Kiang, 1965а). Это означает, что
интервал между импульсами определяется
собственной частотой колебаний той
области базилярной мембраны, с которой
данное волокно связано.
219
При
исследовании реакции слуховых волокон
на звуковой стимул обычно предварительно
определяют оптимальную (или
характеристичную) частоту, т. е. такую
частоту, порог реакции на которую
является наиболее низким для данного
волокна. Характеристичная частота
позволяет судить о той области базилярной
мембраны, с которой связано исследуемое
волокно. Скрытый период реакции волокон
слухового нерва на звуковой стимул
(щелчок) зависит от характеристичной
частоты волокна: для волокон с
характеристичной частотой выше 2 кгц
величина скрытого периода может
составлять всего около 1 мсек.; при
понижении характеристичной частоты
величина скрытого периода закономерно
возрастает до 4—5 мсек, благодаря
увеличению времени распространения
бегущей волны от основания улитки к ее
вершине, т. е. к области кортиева
органа, дающей максимальную реакцию
на звуки низкой частоты (Bekesy, 1960).
Зависимость
порога реакции на тональный сигнал от
частоты тона характеризуется так
называемыми частотнопороговыми кривыми
(рис. 72). Область, ограниченная
частотнопороговой кривой, представляет
собой область звуковых частот, вызывающих
ответ данного волокна, если интенсивность
звука выше соответствующего порогового
значения. Частотнопороговые кривые
характеризуют ширину частотной области
базилярной мембраны улитки, с которой
связано данное волокно. Как показывает
рис. 72, частотная область ответа для
разных волокон является различной. В
целом, однако, эти кривые являются более
острыми,чем кривые, характеризующие
частотную избирательность базилярной
мембраны улитки и представленные на
рис. 52. Это означает, что частотная
избирательность волокон слухового
нерва выше, чем частотная избирательность
базилярной мембраны. Механизмы,
обеспечивающие обострение частотной
избирательности при переходе от
рецепторного аппарата улитки к
волокнам слухового нерва, остаются
неясными. Частотная избирательность
волокон является выражением
пространственного принципа передачи
информации о частоте звука, основы
которого заложены в свойствах самой
улитковой перегородки (см. главы 5 и
10).
Существенно
отметить, что при действии тонального
сигнала время появления импульса
соответствует определенной фазе
звуковой волны, по крайней мере в
случаях, когда частота звука не превышает
5 кгц (Kiang, 1965а; Rose et al., 1967). При этом,
однако, импульс, как правило, возникает
в ответ не на каждую звуковую волну, в
результате чего временная картина
последовательности импульсов при
обычной осциллографической регистрации
представляется непериодической,
случайной, причем неперио- дичность
наблюдается даже при частоте звука
ниже 100 гц. Лишь при специальном машинном
способе обработки данных удалось
показать связь между временем появления
импульса и фазой звуковой волны
(рис. 73): статистически интервал между
последо
220
вательными
импульсами в реакции на тон ниже 5 кгц
оказался кратным числу периодов звуковой
волны действующего стимула. Это явление
может иметь существенное значение для
передачи волокнами слухового нерва
сведений о частоте звука. Согласно
распространенной гипотезе, которая
получила название «принципа залпов»
(Wever, 1949), информация о частоте звука
может передаваться группой нервных
волокон, которые дают разряд в одну и
ту же фазу звуковой волны, хотя в
отдельности эти волокна от-
Рис.
72. Частотнопороговые кривые волокон
слухового нерва кошки
(по: Kiang, 1965а).
По
оси абсцисс —
частота звука в кгц; по
оси ординат —
пороговая интенсивность
звука, в дб
затухания относительно условного
исходного уровня.
вечают
не на каждую звуковую волну. То
обстоятельство, что возникновение
нервного импульса связано с определенной
фазой звуковой волны, обеспечивает
появление синхронного залпа одновременно
в нескольких волокнах, причем такой
суммарный залп будет возникать в ответ
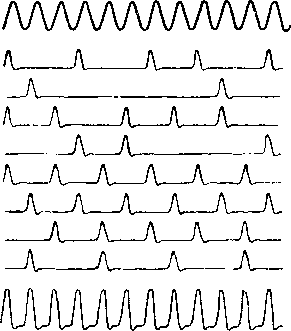
на каждую звуковую волну (рис. 74). В
результате группа волокон оказывается
способной передать информацию о частоте
звука на основе определенной временной
последовательности возникновения
разрядов в отдельных волокнах этой
группы (рис. 73).
Изменение
интенсивности звукового стимула находит
свое отражение в частоте импульсации
волокон слухового нерва. При увеличении
интенсивности звукового стимула число
импульсов в реакции на него возрастает
за счет увеличения частоты следования
импульсов в реакции на звук. При этом
максимальная частота импульсации
в волокнах слухового нерва при действии
221
5.0' 10.0
' 15.0 ' 200мсек.
Рис.
73. Гистограммы межимпульсных
интервалов, характеризующие реакцию
волокна слухового нерва обезьяны
на тон различной частоты (Rose et al., 1967).
Цифры
над гистограммами —
частота тона в гц. Точки
под гистограммами
— отметки
периода тонального сигнала.
тона
оптимальной частоты, по имеющимся в
литературе данным, в отдельных случаях
может достигать 800 имп./сек.; обычно эта
частота значительно ниже и приближается
к 100 — 200 имп./сек. Характер зависимости
частоты импульсации (F)
от
интенсивности (7) тонального сигнала
может быть различным: у подавляющего
большинства нейронов соответствующие
кривые
222


достигают
некоторого преде-
льного уровня
(«плато»); при
дальнейшем увеличении
интен-
сивности кривая в ряде слу-
чаев
идет вниз, т. е. часто-
та импульсации
уменьшается.
Лишь у небольшей части
ней-
ронов (примерно у 10% —по
данным
Nomoto et al., 1964)
кривые F (7) не достигают
уров-
ня насыщения вообще. Дина-
мический
диапазон, т. е. диа-
пазон интенсивности,
в пре-
Рис.
75. Постстимульные гисто-
граммы,
характеризующие по-
давление
спонтанной (б. зв.) и
вызванной
активности волокна
слухового нерва
кошки при вклю-
чении тональной
посылки 21 кгц
на фоне длящегося тона
оптималь-
ной частоты (18.5 кгц) (по:
Kiang
1965а).
Цифры
слева —
частоты действующих стимулов, справа
—
число импульсов.
Рис.
74. Схематическая иллюстра-
ция
принципа «залпов» (по:
Schwartzkopff, 1961).
Вверху
—
звуковой сигнал; следую-
щие
8 строк —
реакция отдельных
волокон; нижняя
кривая
— суммарная
реакция этих волокон.
делах
которого частота импульсации возрастает
с возрастанием интенсивности звука, у
разных волокон может быть различным.
Японские авторы (Nomoto et al., 1964) показали,
что две трети слуховых волокон
обнаруживают примерно одинаковый ход
кривых F
(Z) при воздействии сигналов различной
частоты, в то время как у 20% волокон
динамический диапазон на
223
оптимальной
частоте существенно больше, чем на
других. Связывая крутой ход кривых F
(/) на оптимальной частоте с локальностью
иннервации кортиева органа данным
нервным волокном, авторы приходят
к выводу, что волокна первой группы
являются радиальными, а волокна второй
— спиральными. Это согласуется с
электронномикроскопическими данными
о численном соотношении радиальных и
спиральных волокон (Spoendlin, 1966).
Скрытый
период реакции слуховых волокон на
звуковой сигнал сокращается с увеличением
его интенсивности (Kiang, 1965а), однако эта
зависимость не была исследована при
действии звуков различной частоты.
Интересно
отметить, что уже на уровне волокон
слухового нерва имеют место тормозные
явления, в частности полное подавление
импульсной активности (не только
вызванной, но и спонтанной) при
одновременном действии двух звуков
(рис. 75). При этом звуковые частоты,
вызывающие торможение активности,
вызванной тоном характеристичной
частоты, лежат по обе стороны от
частотнопороговой кривой (Sachs, Kiang,
1968). Таким образом, область тормозящих
частот непосредственно граничит с
областью ответа нейрона и даже частично
перекрывает ее. Скрытый период торможения
активности при взаимодействии двух
звуков составляет менее 2 мсек.
(Nomoto et al., 1964), а явление наблюдается и
после интракраниальной перерезки
слухового нерва (Kiang, 1965а).
Имеются
также отдельные указания на подавление
спонтанной активности волокон слухового
нерва при действии одного звука
определенной частоты (Rupert et al., 1963; Sachs,
Kiang, 1968) или щелчка (Kiang, 1965a); в последнем
случае подавление спонтанной
активности возникает в фазу повышения
давления на барабанную перепонку.
Возможные механизмы организации
тормозных процессов на уровне 1
афферентного нейрона слуховой системы
дискутируются в литературе (Nomoto et al.,
1964; Kiang, 1965a).
Импульсная
активность слухового нерва регулируется
через систему нисходящих (эфферентных)
волокон оливо-кохлеарного пучка
Расмуссена, идущих к улитке из ядер
верхней оливы. Эти волокна оканчиваются
на волосковых клетках улитки и, кроме
того, дают синаптические контакты с
окончаниями афферентных слуховых
волокон. При электрическом раздражении
оливокохлеарных пучков (как
перекрестного, так и неперекрестного)
происходит подавление импульсации,
вызванной звуком, в афферентных
волокнах слухового нерва и уменьшение
амплитуды суммарного нервного ответа
(Нх)
на звуковой щелчок. При этом эффект от
раздражения перекрестного пучка в
несколько раз сильнее, чем неперекрестного.
В опытах с предварительной перерезкой
оливо-кохлеарных волокон было
показано ухудшение различения сигнала
в шуме в этих условиях, а также ухудшение
частотного различения. По-видимому, в
естественных условиях слухового
восприятия
224
система
нисходящих связей, в которую входит
оливо-кохлеарный пучок, служит настройке
периферического аппарата слуховой
системы на оптимальный режим восприятия
звуковых сигналов.
Основные
сведения о характеристиках электрической
активности волокон слухового нерва и
сводку литературы по этим вопросам
можно найти в ряде работ (Katsuki et al., 1962;
Kiang et al., 1962; Nomoto et al., 1964; Kiang, 1965a, 1965b; Rose
et al., 1967, 1969; Fex, 1968; Радионова, 1971).
Кохлеарные
ядра
Все
афферентные волокна слухового нерва
передают импульсацию в комплекс
кохлеарных ядер, которые являются
первым центральным отделом слуховой
системы.
Спонтанная
активность нейронов кохлеарных ядер
ниже, чем волокон слухового нерва и по
числу спонтанно активных нейронов,
и по частоте импульсации. У кошки в
условиях барбитурового наркоза
спонтанной активностью обладает
примерно половина нейронов, причем
средняя частота спонтанной импульсации
в подавляющем большинстве случаев
не превышает 40—50 имп./сек. Было установлено
(Koerber et al., 1966), что спонтанная активность
нейронов вентральных отделов кохлеарного
комплекса целиком определяется
приходящей импульсацией от улитки:
после разрушения улитки нейроны этих
областей утрачивают спонтанную
активность. В то же время спонтанная
активность нейронов дорсального
кохлеарного ядра при этом сохраняется
неизменной и, следовательно, имеет
другое происхождение.
Узор
разряда на тональный сигнал оптимальной
(характеристичной) частоты является
значительно более многообразным для
нейронов кохлеарных ядер, чем для
волокон слухового нерва. Это, очевидно,
определяется многообразием межнейронных
связей на уровне кохлеарных ядер
(см. главу, посвященную структуре).
На рис. 76 показаны основные типы
постстимульных гистограмм для
нейронов кохлеарных ядер, выделенные
Киангом с соавторами (Kiang et al., 1965).
Одна часть нейронов отвечает на
тональный сигнал характеристичной
частоты разрядом, временной узор
которого весьма сходен с временным
узором ответа волокон слухового нерва
(рис. 76, Л); такой ответ представляет
собой относительно равномерный
тонический разряд с более или менее
выраженным процессом адаптации.
Другой тип узора разряда отличается
от первого наличием выраженной
периодичности в проявлении импульсов
(рис. 76, Б),
причем эта периодичность не связана с
частотой действующего звука. Третий
тип разряда представляет собой on-ответ
(рис. 76, Г).
Наконец, четвертая разновидность узора
импульсного разряда характеризуется
ответом на включение, за которым
после паузы следует тонический разряд
(рис. 76, В).
Нейронам с тоническим узором разряда,
как правило, свойствен про-
15
Сенсорные системы
225

цесс
адаптации; длительность процесса
адаптации, как и на уровне волокон
слухового нерва, может быть различной
и в ряде случаев достигает нескольких
минут (Rose et al., 1959). Даже перечисленные
основные типы узоров импульсного
разряда, не исчерпывающие многообразные
варианты, говорят о достаточно сложной
организации функции кохлеарных
ядер. При этом следует подчеркнуть, что
узор разряда зависит от параметров
звукового раздражения — частоты
звука, его интенсивности, длительности.
Кроме
перечисленных видов узора импульсного
разряда, нейроны кохлеарных ядер
нередко обнаруживают лишь подавление
спонтанной импульсации при действии
звука — при соответствующем подборе
его частоты и интенсивности (рис. 76, Д).
В ряде случаев узор разряда может иметь
сложную конфигурацию — с тормозными
паузами и последействием (рис. 76, Е).
Узор разряда в виде короткой пачки
импульсов (рис. 76, Ж), который характерен
для нейронов более высоких уровней
слуховой системы, на уровне кохлеарных
ядер встречается крайне редко.
При
действии короткого звукового раздражения,
например щелчка, нейроны кохлеарных
ядер, как правило, отвечают одним- двумя
импульсами с коротким скрытым периодом.
Однако нейроны, дающие в ответ на
тональный сигнал разряд типа,
представленного на рис. 76, Б,
В,
отвечают на щелчок со значительно
большим скрытым периодом, величина
которого обнаруживает значительные
флуктуации. Кроме того, при достаточно
низкой характеристичной частоте (/0)
(в литературе приведена единственная
иллюстрация при fQ=0.6
кгц— Kiang et al., 1965), в кохлеарных ядрах
может наблюдаться такое же явление,
как и в волокнах слухового нерва, т. е.
отражение периода /0
в постстимульной гистограмме нейрона,
отвечающего на щелчок.
Пространственная
упорядоченность хода слуховых волокон
(см. выше) определяет существование
тонотопической локализации во всех
основных отделах кохлеарных ядер. Это
проявляется в том, что при проходе
электрода через тот или иной отдел
кохлеарного комплекса последовательно
встречаемые нейроны обнаруживают
определенную закономерность расположения
по своим характеристичным частотам:
так, при ходе электрода в дорсо-вентраль-
ном направлении вначале встречаются
нейроны с наиболее высокой
характеристичной частотой, затем — с
более низкой и нако-
Рис.
76. Варианты узора разряда для различных
нейронов (А—
Ж)
в кохлеарном ядре кошки.
А—Г
—
по: Kiang et al., 1965; ответы на тон длительностью
25 мсек.* задержка сигнала относительно
начала гистограммы 2.5 мсек. Д—Ж
—
ориг.; ответы на тон длительностью 100
мсек. Д, Ж — реакция на сигнал постоянной
интенсивности. Е
—
реакция на сигнал различной интенсивности
(цифры
слева —
дб над уровнем 0.0002 Дин/см2.
На Е,
Ж
регистрация «методом точек»: первый
вертикальный ряд точек
— момент подачи сигнала, остальные
точки —
импульсация нейрона.
15*
227
нец
— с самой низкой (рис. 77). Таким образом,
в кохлеарных ядрах многократно
представлена частотная проекция улитки.
Частотнопороговые
кривые нейронов всех основных отделов
кохлеарных ядер в общем сходны и при
этом принципиально не отличаются
от частотнопороговых кривых волокон
слухового нерва (рис. 78). Следовательно,
на уровне кохлеарных ядер не происхо-
Рис.
77. Схема сагиттального среза через
левый комплекс кохлеар-
ных ядер
кошки, иллюстрирующая последовательность
оптималь-
ных частот нейронов по
ходу микроэлектрода (по: Rose et al., 1959).
А
—
переднее вентральное кохлеарное ядро;
С
—
мозжечок; D
—
дорсальное кохлеарное ядро; Р
—
заднее вентральное кохлеарное ядро.
Цифры
—
оптимальные частоты нейронов. Ход
микроэлектрода обозначен прямой
линией, пересекающей
D
и Р.
Штриховые линии —
условные границы между отдельными
областями кохлеарного комплекса.
дит
дальнейшего обострения частотной
избирательности нервных элементов по
сравнению с волокнами слухового нерва.
Напротив, в ряде случаев на частотнопороговых
кривых нейронов кохлеарных ядер
имеется дополнительный пик в области
частот более низких, чем характеристичная;
однако этот дополнительный пик, как
правило, выражен сравнительно слабо и
лежит в области значительно более
высоких уровней интенсивности, чем
основной пик, связанный с характеристичной
частотой нейрона.
Привязанность
импульсного разряда нейронов кохлеарных
ядер к фазе тонального сигнала остается
неисследованной. Косвенным
свидетельством в пользу существования
такой привязанности
228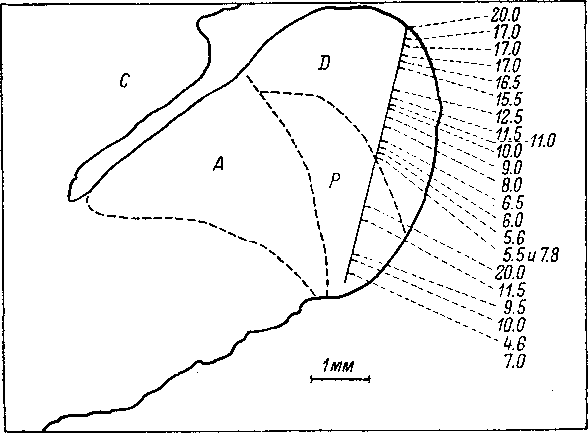
является
воспроизведение частоты тона в суммарном
ответе кохлеарных ядер до 4—5 кгц.
Этот предел воспроизведения частоты
раздражения в суммарном ответе не ниже
ритма раздражения, воспроизводимого
в слуховом нерве.
Реакция
нейронов кохлеарных ядер на увеличение
интенсив
ности
звука, как правило, проявляется в
увеличении числа и ча
стоты
импульсов в разряде и уменьшении
скрытого периода реакции
и
его дисперсии. Однако
быть выражены
в раз-
личной степени. Кроме
того,
изменение интен-
сивности звука
может
приводить к качествен-
ному
изменению узора
разряда*нейронов:
к по-
явлению или исчезнове-
нию
тормозных пауз, к
Рис.
78. Частотнопорого-
вые кривые для
нейронов
кохлеарных ядер
кошки
(Радионова, 1971).
По
оси абсцисс —
частота звука
в ьтц; по оси
ординат —
ин-
тенсивность звука (Jo)
в дб от-
носительно 0.0002 дин/см2.Пруж-
ки
со стрелкой —
уровень ин-
тенсивности ниже
порога
реакции.
у
разных нейронов эти явления могут
определенной
группировке импульсов в пачки и т. д.
Максимальная частота импульсации,
вызванной длящимся звуком достаточной
интенсивности, у нейронов кохлеарных
ядер нередко может достигать 200—300
имп. /сек. и в отдельных случаях — даже
600 имп./сек. Таким образом, если предельная
цифра, приведенная в литературе и
являющаяся уникальной (600 имп./сек., см.:
Moller, 1969), для кохлеарных ядер ниже
предельной цифры для волокон слухового
нерва (800 имп./сек.), то максимальная
частота импульсации, достигаемая
довольно часто (200—300 имп./сек.), даже
несколько превосходит обычный предел
частоты импульсации волокон слухового
нерва (100—200 имп./сек.). Нередко при
увеличении интенсивности звука нейроны
по достижении некоторой предельной
частоты импульсации в дальнейшем
обнаруживают ее уменьшение вплоть до
полного подавления реакции на звук.
Все описанные явления при изменении
интенсивности звука могут быть в
различной степени выражены на
оптимальной и неоптимальных частотах.
229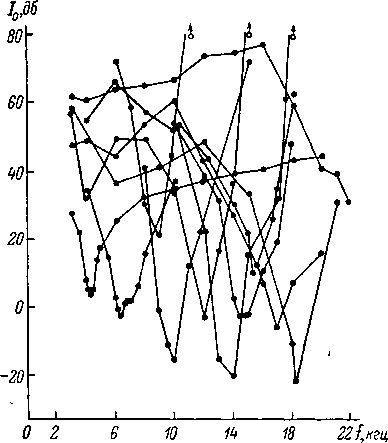
Скрытый
период реакции при действии звука
достаточно высокой интенсивности
для подавляющего большинства нейронов
составляет 2—4 мсек, и возрастает при
понижении интенсивности стимула, причем
для подавляющего большинства нейронов
его величина в околопороговой области
интенсивности звука опти-
Рис.
79. Характеристики нейронов коротколатентного
типа
в кохлеарном ядре кошки.
а
—
реакции на сигнал оптимальной частоты;
б
—
частотно пороговые кривые при длительности
звука 100 и 1 мсек. На а: сверху
вниз —
узор; импульсного разряда в ответ на
сигнал интенсивностью около 10 дб над
порогом реакции, длительностью 100 мсек.,
регистрация «методом точек» (см. рис.
76), вверху
отметка действия сигнала; зависимость
скрытого периода (СП, мсек.) от
интенсивности звука (Г, дб); зависимость
порога реакции (10,
Дб) от длительности звука (/, мсек.);
зависимость числа импульсов в ответе
(X) от уровня интенсивности звука (1, дб).
На б: по
оси абсцисс — частота
звука (/, кгц), по
оси ординат —
пороговая интенсивность звука
.
(То, Дб).
мальной
частоты и достаточной длительности
(порядка 100 мсек.) достигает нескольких
десятков миллисекунд. При переходе к
неоптимальной частоте эта величина
у части нейронов резко сокращается
и составляет всего несколько миллисекунд.
На
уровне кохлеарных ядер, как это показано
в исследованиях на кошке, может быть
выделена специальная группа нейронов,
свойства которых в значительной степени
обеспечивают быструю передачу
сведений о появлении звукового сигнала
в высшие
230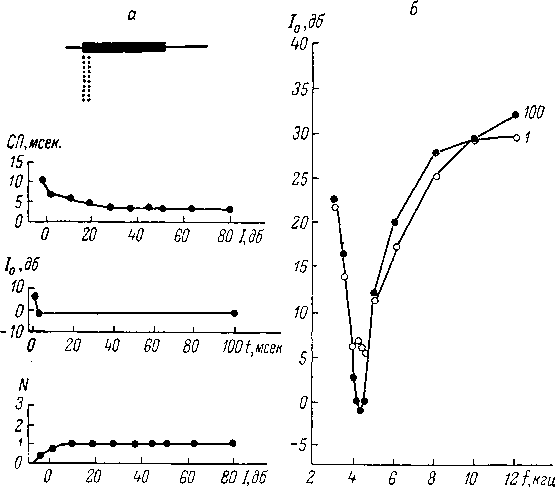
отделы
слуховой системы. Это группа нейронов,
реагирующих на звук по типу фазного
оп-ответа (рис. 76, Г)
или он—off-ответа, по крайней мере, в
области слабых, близких к порогу
интенсивностей звука (при больших
уровнях интенсивности узор разряда
этих нейронов может оказаться иным).
Такие нейроны имеют короткий скрытый
период реакции на звук (до 10 мсек.) даже
при низком уровне его интенсивности.
Дисперсия скрытого периода реакции
этих нейронов также мала и составляет
обычно доли миллисекунды. Их пороги
практически не зависят от длительности
стимула (отсутствие способности к
временной суммации на пороге реакции),
в результате чего характер частотнопороговых
кривых, свойственный нейронам этой
группы, оказывается сформированным
уже при очень малой длительности звука
(1—2 мсек.). Некоторые из основных свойств
этих нейронов представлены на рис. 79.
Эта группа нейронов может быть названа
группой коротколатентных, малосуммирующих
нейронов с фазным характером разряда
на пороге реакции. В кохлеарных ядрах
кошки эти нейроны составляют около 20%
всех нейронов (Радионова, 1967). Интересно
отметить, что среди волокон слухового
нерва кошки, по данным Кианга, элементов
с фазным разрядом не было обнаружено
несмотря на очень большое количество
обследованных волокон. В то же время у
обезьяны уже среди волокон слухового
нерва встречаются элементы, отвечающие
фазным разрядом на включение и выключение
звука (Katsuki et al., 1962). Однако если судить
по величине скрытого периода и его
дисперсии в области даже околопороговых
интенсивностей (Tasaki, 1954), то волокна
слухового нерва следует рассматривать
как коротколатентные.
Основная
масса нейронов кохлеарных ядер обладает
свойствами, существенно отличающими
их от нейронов коротко латентного
типа (рис. 80). Это нейроны с тоническим
узором разряда в широком диапазоне
интенсивности звука, вплоть до
околопороговых. Их скрытый период
резко зависит от интенсивности звука,
возрастая на десятки миллисекунд при
уменьшении интенсивности стимула;
при этом дисперсия скрытого периода
также резко возрастает. Порог реакции
этих нейронов существенно зависит
от длительности звука: при увеличении
длительности от 1 до 50—100 мсек, порог
реакции понижается на 20—30 дб, а в
отдельных случаях даже на 40—50 дб. Иными
словами, эти нейроны обладают хорошо
выраженной способностью к временной
суммации, илинакоплению сведений о
сигнале во времени. Характерной
особенностью многих нейронов этой
группы является то, что перечисленные
их свойства лучше всего выражены при
действии оптимальной частоты звука.
При переходе от оптимальной частоты к
неоптимальной временные характеристики
этих нейронов (в их количественном
выражении) нередко начинают приближаться
к временным характеристикам нейронов
первой, коротколатентной
231
группы
(ср. рис. 79, а
и 80, в). Иными словами, поведение нейронов
с длиннолатентным типом реакции на
оптимальной частоте звука может не
отличаться от поведения нейронов с
коротколатентным типом реакции,
если частота звука является неоптимальной.
В результате такого различия в свойствах
на оптимальной и неоптимальной
частотах частотнопороговые кривые
этих нейронов
Рис.
80. Характеристики нейронов длинно
латентного типа в кохлеарном
ядре
кошки.
На
а, б — то же, что и на рис. 79. На в
—
те же характеристики, что и на а, но при
действии звука неоптимальной частоты.
имеют
различный вид при действии коротких
(порядка 1 мсек.) и длительных (порядка
100 мсек.) звуков (рис. 80, б): в последнем
случае кривая является более острой
за счет более резко выраженной
временной суммации на оптимальной
частоте. Таким образом, в приведенном
случае имеет место своеобразный механизм
частотного обострения — временной
механизм, связанный с временными
характеристиками нейронов. К сожалению,
вопрос о существовании такого
механизма на уровне I нейрона (волокон
слухового нерва) исследован не был. Что
касается более высоких уровней
232
слуховой
системы, то на уровне задних холмов
этот временной механизм частотного
обострения хорошо выражен (см. ниже).
Тормозные
явления в кохлеарных ядрах представлены
весьма многообразно, о чем свидетельствует
уже многообразие узора импульсного
разряда нейронов. Сложный характер
узора разряда в значительной мере
обусловлен наличием тормозных пауз —
как на фоне действующего звука, так и
в его последействии. Вообще тормозные
явления на уровне кохлеарных ядер имеют
различное выражение — например, в виде
немонотонного характера зависимости
числа импульсов от интенсивности
сигнала, когда в диапазоне средних и
высоких уровней интенсивности звука
число импульсов в ответе на сигнал
меньше, чем в диапазоне малых уровней
интенсивности. Тормозные явления имеют
место также при одновременном действии
двух звуков — при определенном
соотношении их частоты и интенсивности
(явление маскировки). Кроме того, в
работе по исследованию реакции нейронов
кохлеарных ядер на частотно-модулированные
сигналы (Erulkar et al., 1968) было показано
существование определенной зависимости
реакции от направления модуляции звука
по шкале частот — явление, которое
также может быть объяснено лишь с учетом
тормозных явлений в межнейронном
взаимодействии.
Нейроны
кохлеарных ядер в целом характеризуются
достаточно высокой стабильностью
характеристик как спонтанной, так и
вызванной активности. Лишь очень
малая часть нейронов (примерно 0.5%)
обнаруживает «привыкание» к раздражителю
и перестает отвечать на него после
сравнительно небольшого числа
предъявлений. Как известно, такая
довольно сложная форма поведения в
большей степени свойственна нейронам
верхних уровней слуховой системы и
мультисенсорным нейронам подкорковых
и таламических ядер.
Регуляция
деятельности нейронов кохлеарных ядер
со стороны различных отделов центральной
нервной системы экспериментально
доказана и, по-видимому, хорошо развита.
История изучения и современное
состояние этого вопроса рассматривается
в работах Уитфильда (Whitfield, 1967) и Е. А.
Радионовой (1971). Известны несколько
нисходящих путей, оканчивающихся на
нейронах кохлеарных ядер. В дорсальном
кохлеарном ядре оканчиваются пути,
идущие из ипси- и контрлатеральных ядер
лемниска; третий путь идет от вентрального
ядра лемниска к вентральным отделам
кохлеарных ядер противоположной
стороны; четвертый путь идет от
S-образного ядра верхней оливы к
вентральному кохлеарному ядру своей
стороны. Описан также путь, идущий в
кохлеарные ядра из мозжечка. В
физиологических исследованиях было
показано, что ответ на звук в кохлеарных
ядрах подавляется при отвлечении
внимания животного, при многократном
предъявлении звука; спонтанная
активность подавляется при стимуляции
противоположного уха после деафферентации
кохлеарных ядер.
233
В
ряде более поздних исследований было
установлено также существование
положительных эффектов на электрическую
активность кохлеарных ядер: при
стимуляции оливо-кохлеарного пучка
можно наблюдать не только подавление,
но и усиление электрической активности
нейронов кохлеарного комплекса, а также
понижение порога их реакции на звук.
Таким образом, все эти данные
свидетельствуют о сложном характере
организации связей кохлеарных ядер
с другими отделами центральной нервной
системы; эти связи, по-видимому,
обеспечивают определенный уровень
настройки свойств нейронов кохлеарных
ядер в условиях естественного восприятия
звука.
Сводку
литературы и фактического материала
по функциональной характеристике
кохлеарных ядер можно найти в ряде
работ (Rose et al., 1959; Greenwood, Maruyama, 1965; Kiang et
al., 1965; Whitfield, 1967; Gerstein et al., 1968; Радионова,
1971).
Верхняя
олива
Как
уже указывалось выше (стр. 200), верхняяолива
является первым уровнем в слуховой
системе, где наблюдается билатеральная
конвергенция афферентации: основные
афферентные пути от правого и левого
кохлеарных ядер конвергируют на
нейронах, составляющих определенные
ядра верхнеоливарного комплекса.
При
исследовании спонтанной активности
нейронов верхней оливы было
установлено, что по количеству
спонтанноактивных нейронов отдельные
ядра верхнеоливарного комплекса
существенно различаются между собой.
Так, по данным Кларка и Данлопа (Clark,
Dunlop, 1969), наибольшее число нейронов,
обладающих спонтанной активностью,
зарегистрировано в ядре трапециевидного
тела (39% от общего количества обследованных
нейронов), несколько меньшее количество
— в медиальном (акцессорном) ядре
(21%) и единичные спонтанно-активные
нейроны были обнаружены в латеральном
(S-образном) ядре (7%). Спонтанная
активность нейронов перечисленных
выше ядер верхней оливы достаточно
высока (20—120 имп./сек.) и нерегулярна.
Значительно более редкой (от нескольких
единиц до 40—50 имп./сек.) и более регулярной
спонтанной активностью обладают
нейроны, дающие начало эфферентному
оливо-кохлеарному пучку Расмуссена
(стр. 209).
При
действии тональных сигналов длительностью
в 50—100 мсек, обнаружено большое
разнообразие узоров разряда верхней
оливы. Эти узоры разряда, схематически
представленные на рис. 81, могут быть
подразделены на те же группы, которые
были выделены уже в кохлеарных ядрах
(рис. 76): относительно равномерная
импульсация в течение всего времени
действия звука (I—7), импульсация с
выраженной временной структурой
разряда (S—10.
12)
и реакция только на включение (либо на
включение и выключение) тональной
посылки (11,
14—16).
При этом сле-
234
дует
указать, что количественное соотношение
нейронов с различ-
ным узором
разряда в основных ядрах оливарного
комплекса
существенно различается.
Так, в ядре трапециевидного тела ней-
роны
характеризуются различными вариантами
тонического узора
разряда,
в латеральном
(2%
от
общего числа
обследованных),
отвеча-
ющие только на включе-
ние
звукового сигнала,
в то время как
в меди-
альном ядре число ней-
ронов,
отвечающих на
включение сигнала,
до-
стигает одной трети от
общего
числа обсле-
дованных элементов
(рис.
81).
Исследование
реак-
ции нейронов верхней
оливы
при действии
Рис.
81. Схематическое изображение узоров
разряда (1—16)
нейронов основных ядер верхнеолй-
варного комплекса кошки при действии
тональной посылки (по: Clark, Dunlop,
1969).
ТТ
—
ядро трапециевидного тела; МЯ
—
медиальное ядро верхней оливы; ЛЯ
—
латеральное ядро верхней оливы.
Цифры
—
количество нейронов в процентах,
-у которых наблюдался тот или иной
тип разряда.
ядре
появляются отдельные нейроны |
ТТ |
МЯ |
ЛЯ |
|
ТТ |
МЯ |
ЛЯ |
|
°/o |
°/о |
°/о |
|
°!о |
% |
|
||
|
5 |
3 |
0 |
|Ы |
6 |
0 |
9 |
|
|
|
|
|
|
|
|
||
|
20 |
21 |
20 |
| ^0 |
1 |
к |
18 |
|
|
|
|
|
|
|
|
|
|
|
1 |
3 |
0 |
1 11 |
0 |
|
0 |
|
|
|
|
|
|
|
|
||
|
29 |
19 |
36 |
1 * 12 |
0 |
3 |
0 |
|
|
|
|
|
|
|
|
|
|
|
Ч |
0 |
0 |
13 |
0 |
ч |
0 |
|
|
|
|
|
|
|
|
||
_ЛИ |
|
5 |
|
13 |
| /4 |
0 |
18 |
2 |
|
|
|
|
|
|
|
|
|
|
29 |
5 |
7 |
II |
0 |
ч |
0 |
|
|
|
|
|
|
|
|
||
|
0 |
3 |
0 |
161 |
0 |
5 |
0 |
|
длительных
звуковых сигналов (секунды и десятки
секунд) позволило установить, что в
их импульсации наблюдается выраженная
адаптация. При этом время, в течение
которого после включения звука происходит
установление импульсации на постоянном
(более низком, чем исходный) уровне (от
3—4 до 20 сек.), зависит от интенсивности
воздействующего стимула: чем выше
уровень интенсивности, тем медленнее
наступает адаптация. Следует указать,
что в отдельных случаях уменьшение
импульсации до ее полного подавления
продолжалось в течение 30 мин. после
включения звука (Clark, Dunlop, 1969).
235
При
действии коротких звуковых щелчков
нейроны верхне-
оливарного
комплекса реагируют, как правило,
разрядом, состоящим из 1—3 импульсов,
и лишь у отдельных нейронов действие
одиночного щелчка вызывает непрерывный
разряд, длящийся десятки миллисекунд
(Watanabe et al., 1968). Следует отметить, что у
ряда нейронов верхней оливы, обладающих
низкой характеристичной частотой,
была обнаружена реакция, наблюдавшаяся
и при исследовании волокон слухового
нерва: при действии звукового щелчка
разряд характеризовался последовательностью
локализация
в S-
Tsu-
Рис.
82. Частотная
образном
ядре верхней оливы (по:
chitani, Boudreau,
1966).
Цифры
—
частота в кгц.
из
двух или нескольких им-
пульсов,
интервал между ко-
торыми соответствовал
пе-
риоду их характеристичной
частоты.
При
исследовании ней-
ронов верхнеоливарного
ком-
плекса в условиях действия
тональных
сигналов различ-
ной частоты было
установле-
но, что характеристичные
частоты
отдельных нейронов
у кошек и собак
перекрывают
диапазон от 0.2 до 40
кгц.
При этом наблюдались
ча-
стотнопороговые кривые раз-
личной
остроты, симметрич-
ные и несимметричные,
в от-
дельных
случаях с двумя
или тремя минимумами.
Си-
стематическое исследование
нейронов
S-образного ядра
позволило установить,
что
более широкие (при оценке
герцах)
частотнопороговые кривые
по
абсолютному значению в
наблюдаются
при более высоких характеристичных
частотах;
асимметрия
частотнопороговых кривых у нейронов
этого ядра
проявляется, как правило,
лишь при больших уровнях интенсив-
ности
стимула (Tsuchitani, Boudreau, 1966). Что касается
тоното-
пической локализации, то в
трапециевидном и акцессорном ядрах
кошки
к настоящему времени она не была
обнаружена, в то время
как в акцессорном
ядре собаки достаточно хорошо
выражена.
Отчетливая тонотопическая
локализация была установлена в
S-образном
ядре оливарного комплекса кошки (рис.
82).
Нейроны
верхнеоливарного комплекса, так же как
и нейроны
нижележащих отделов
слуховой системы, обнаруживают способ-
ность
воспроизводить импульсным разрядом
частоту тонального стимула. При этом
импульсный разряд возникает либо на
каждый
236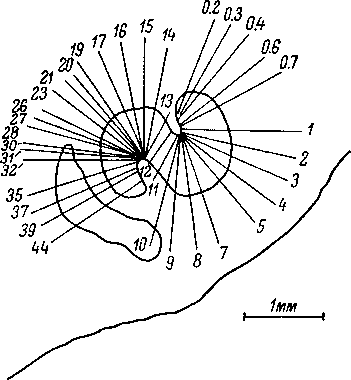
период
тонального сигнала, либо через интервалы,
кратные длительности периода (рис.
83). Это явление наблюдалось при стимуляции
тонами с частотой до 2 кгц.
При
увеличении интенсивности тонального
сигнала характеристичной частоты
количество импульсов у большинства
нейронов возрастает монотонно. Лишь в
отдельных случаях обнаружен немо
нотонный
ход кривой зависимости количества
импульсов от интенсивности стимула.
Особенности изменений величины реакции
при варьировании интенсивности
стимуляции в условиях действия
тональных сигналов неоптимальной
частоты к настоящему времени подробно
не исследованы.
Скрытый
период реакции нейронов верхнеоливарного
комплекса в основном иссле-
1000
20-
Рис.
83. Гистограммы межимпульсных
интервалов нейрона верхнеоливарного
комплекса (по: Moushegian et al., 1964b).
По
оси абсцисс —
интервал в мсек.;
по
оси ординат —
количество интер-
валов в процентах.
Цифры
над гисто-
граммами —
частота тонального сиг-
нала (гц),
использованного в качестве
раздражения.
О
0.8 1.6 2.9 3.2 9.0 9.8 5.6 6.9 7.2
1999
20-
10
- д
О
0.9 0.8 1.2 1.6 2.0 2.9 2.8 3.2 3.6
довался
при действии коротких звуковых щелчков.
Было установлено, что при средних
интенсивностях щелчка скрытый период
реакции у разных нейронов варьирует
от 3 до 12 мсек. Так же как и у нейронов
нижележащих отделов слуховой системы,
скрытый период реакции нейронов верхней
оливы закономерно изменяется при
изменении интенсивности стимула:
увеличение уровня раздражения
сопровождается укорочением скрытого
периода. Что касается исследования
импульсной активности нейронов
верхней оливы при изменении длительности
сигнала, то подробные данные по этому
вопросу в литературе отсутствуют.
Поэтому классификация нейронов верхней
оливы по критерию особенностей их
реакции на сигналы разной длительности
в настоящее время не представляется
возможной.
Так
же как и в нижележащих отделах слуховой
системы, тормозные влияния на уровне
верхней оливы достаточно хорошо
237
![]()
выражены.
Об этом свидетельствуют структурированные
по времени узоры разряда, схематическое
изображение которых представлено на
рис. 81. Кроме того, в исследовании Чанга
и By (Chang, Wu, 1964) показано, что характерный
для части нейронов ядра трапециевидного
тела длительный скрытый период связан
с развитием тормозных влияний и начальной
гиперполяризацией.
Относительно
специализированных реакций нейронов
верхней оливы имеются лишь отдельные
наблюдения. Так, имеются данные,
свидетельствующие о том, что нейроны
верхнеоливарного комплекса усиливают
свою активность при модуляции частоты
тонального сигнала в одном направлении,
в то время как при противоположном
направлении изменения частоты звука
импуль- сация подавляется (Watanabe et al.,
1968).
Основные
данные, характеризующие импульсную
активность нейронов верхней оливы,
кроме цитированных выше исследований,
представлены в ряде работ (Galambos et al.,
1959; Moushe- gian et al., 1964a, 1964b, 1967; Goldberg et al.,
1964; Goldberg, Brown, 1968, 1969; Rupert et al., 1968).
Задние
холмы
На
клетках задних холмов переключается
большая часть волокон латеральной
петли, состоящей из аксонов нейронов
II, III и IV порядков, клеточные тела которых
располагаются в кохлеарных ядрах,
верхней оливе и ядрах латеральной
петли (Stotler, 1953; см. стр. 201).
Уровень
спонтанной активности нейронов
центрального ядра задних холмов
снижается по сравнению с кохлеарными
ядрами: большинство нейронов разряжается
с частотой 0.5—8 имп./сек., количество
спонтаныоактивных нейронов уменьшается
до 12%.
Нейроны
задних холмов различаются по временному
узору импульсного разряда на сигнал
(рис. 84). Значительная часть нейронов
отвечает начальным разрядом из 1—2
импульсов на включение тона или шума
(рис. 84, Б).
В некоторых случаях ответ возникает
на включение и выключение сигнала (рис.
84, А).
В
отличие от кохлеарных ядер, где фазный
ответ, возникающий на включение, как
правило, наблюдается только при малых
интенсивностях звукового сигнала
(Радионова, 1967), 50% нейронов задних холмов
реагируют фазным разрядом при раздражении
любой интенсивности (Rose et al., 1963). Другая
многочисленная группа реагирует
длительным разрядом, продолжающимся
в течение всего времени действия
раздражителя (рис. 84, Г,
Д). Процесс
адаптации импульсной активности
продолжается 30— 200 мсек, после включения
звука длительностью около 1 сек. Часть
нейронов имеет более или менее выраженный
период отсутствия импульсации после
начального разряда, за которым следует
длительная нерегулярная импульсация.
Кроме описан-
238
Рис.
84. Типы ответов
Ответы
нейронов
(А—Д)
задних холмов кошки.
ных
наиболее часто4
встречающихся узоров ответа, для
нейронов задних холмов характерна
импульсная реакция в виде пачки
импульсов, возникающих при включении
звука (рис. 84, В).
Реакция
такого типа относительно редко
встречается в кохлеарных ядрах (см.
стр. 226). Некоторые нейроны обнаруживают
по
давление
исходного
уровня спонтанной
активности
при дей-
ствии звука. На ко-
роткие
звуковые сиг-
налы нейроны реаги-
руют
разрядом из
1—2 или пачки им-
пульсов,
в последнем
случае длительность
импульсации
превы-
шает длительность
звука.
Пространст
в е н н о
упорядоченное-' пред-
ставительство
звуко-
вых частот в цент-
ральном
ядре задних
холмов было показано
при
исследовании ре-
Рис.
85. Частотная лока-
лизация в задних
хол-
мах кошки (по: Rose
et al., 1963).
Схема
фронтального среза мозга на уровне
заднего холма, ех
—
латеральная кора; с — центральное ядро;
сплошная
прямая линия —
проекция электрода на срезе. Ход
электрода начинается от поверхности
латеральной коры, расположенной на
срезе дорсально, кау- дально и латерально
по отношению к точке наиболее глубокого
погружения, и заканчивается у
медиовентральной границы центрального
ядра. Черточками
на линии хода электрода
указано расположение точек, в которых
определялась характеристичная частота
нейронов. Над
схемой
вынесена проекция электрода с указанием
характеристичных частот (цифры
сверху)
в соответствующих точках. В латеральной
коре (ех)
порядок частот от высоких к низким, в
центральном ядре (с) — от низких к
высоким.
акций
одиночных клеток (Katsuki et al., 1958; Rose et al.,
1963; Гершуни и др., 1969) и вызванных
потенциалов этого отдела (Kemp et al., 1937;
Вартанян и Ратникова, 1967). На рис. 85
представлено схематическое изображение
частотной локализации в Латеральной
коре и центральном ядре задних хол мов.
При проходе микроэлектрода через разные
отделы заднего холма последовательно
встречаются нейроны, связанные с
определенными звуковыми частотами.
В латеральной коре последовательно
встречаются нейроны, оптимальные
частоты которых 240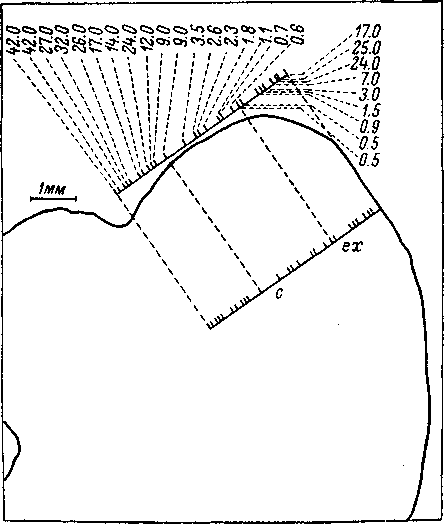
изменяются
от высоких к низким по мере погружения
электрода, в центральном ядре — от
низких к высоким.
Частотнопороговые
кривые (ЧПК) нейронов задних холмов
различны по форме и остроте настройки.
Эрулькар (Erulkar, 1959) различает симметричные
и несимметричные ЧПК. Первые, как
правило, более широкие и пологие, чем
вторые, которые
характеризуются
узкой полосой эффективных частот. На
рис. 86 показаны различные типы
частотнопороговых кривых нейронов
задних холмов: острые (А), пологие (Б),
мультипиковые (Б). В отдельных случаях
частотнопороговые кривые, зарегистрированные
в задних холмах, острее, чем в кохлеарных
ядрах (Katsuki et al., 1959; Suga, 1964). Однако в целом
обострения частотных характеристик
нейронов задних холмов, по сравнению
с предыдущими отделами отметить не
удается. Некоторые нейроны могут
активироваться раздельными полосами
звуковых частот (Erulkar, 1959; Гершуни и
др., 1969). Это явление в слабой степени
уже проявляется в кохлеарных ядрах
(см. стр. 229), в задних холмах оно более
четко выражено; некоторые частотнопороговые
кривые
16
Сенсорные системы
241
имеют
хорошо выраженные 2—3 максимума
чувствительности, находящиеся в
приблизительно октавных соотношениях
(рис. 86, В).
По
характеру изменения количества импульсов
при увеличении интенсивности
раздражения нейроны могут быть разделены
на две группы: нейроны с монотонным
увеличением импульсации при увеличении
интенсивности звукового сигнала и
нейроны с максимумом импульсации при
определенных, немаксимальных уровнях
интенсивности (Rose et al., 1963; Марусева,
1969). Функция зависимости количества
импульсов от интенсивности в ряде
случаев зависят от воздействующей
звуковой частоты. Предельная частота
импульсации в разряде, состоящем из
короткой пачки импульсов на включение
сигнала, составляет около 800 имп./сек.
Обычно средняя частота импульсов в
реакции, продолжающейся в течение
действия звукового сигнала длительностью
200— 1000 мсек., значительно ниже и составляет
100—150 имп./сек.
Скрытый
период нейронов нижних холмов при
больших интенсивностях звука
составляет 5—15 мсек. Самый короткий
скрытый период регистрируется на
характеристичной (оптимальной) для
нейрона частоте (Hind et al., 1963). Однако при
уменьшении интенсивности сигнала
выступают различия нейронов по характеру
изменений скрытого периода. Одна группа
нейронов обнаруживает резкое
увеличение скрытого периода и его
дисперсии при околопороговых
интенсивностях звука. Для нейронов
другой группы характерны незначительные
изменения скрытого периода и его
дисперсии при уменьшении интенсивности
сигнала до около- пороговой величины.
Нейроны с большим скрытым периодом при
околопороговых интенсивностях звука
характеризуются выраженной глубиной
временной суммации (более 10 дб) и
длительным тоническим разрядом.
Нейроны с коротким скрытым периодом
при околопороговой интенсивности
сигнала имеют незначительную
временную суммацию (менее 5 дб) и начальный
фазный разряд при всех интенсивностях
звука. Указанные признаки (узор разряда,
величина скрытого периода на пороге
реакции, глубина временной суммации)
были положены в основу функциональной
классификации нейронов этого отдела
слуховой системы (Гершуни и др., 1969)
подобно функциональной классификации
нейронов кохлеарных ядер (см. выше).
Нейроны
с фазным разрядом, практически не
обладающие способностью к временной
суммации, имеют выраженную частотную
избирательность независимо от
длительности звука (рис. 87, А,
2). В то же время в группе длиннолатентных
сильно суммирующих нейронов с длительным
разрядом на сигнал увеличение
длительности раздражителя приводит
к обострению их частотных характеристик
(рис. 87, Л, 1). Увеличение частотной
избирательности нейронов этой группы
отмечено также при увеличении постоянной
времени звукового сигнала (рис. 87, Б,
1). Оба эти явления, по-видимому, отражают
общий принцип временно-частот-
242
ного
обострения в слуховой системе (Гершуни,
1967), описанный
при
исследовании реакций отдельных клеточных
элементов и
суммарных
электрических ответов разных отделов
слуховой сис-
темы (Вартанян, 19666;
Радионова, 1966, 1967, 1971; Забоева,
1967;
Марусева, 1967;
В артанян, Снетков,
1970).
Таким образом,
частотный анализ
осу-
ществляется простран-
ственно
упорядоченной
многоканальной
систе-
мой, каждый канал ко-
торой
настроен на оп-
ределенную
частоту.
Временные свойства сиг-
нала
могут анализиро-
ваться благодаря
раз-
делению
нейронов по
их
временном характе-
ристикам на две
край-
Рис.
87. Частотнопорого-
вые кривые (ЧПК)
двух
типов нейронов задних
холмов
крысы при действии
коротких, длительных
и на-
растающих по амплитуде
.1
I I 1—1
4 8
12 16 20кгц
сигналов.
А
—
ЧПК при действии звуков с прямоугольным
фронтом включения длительностью
2
мсек, (штриховая
кривая)
и 200 мсек, (сплошная
кривая); 1 —
нейрон с длительным разрядом; 2
—
нейрон с начальным разрядом. Б
—
ЧПК при действии прямоугольных сигна-.
лов длительностью 200 мсек, (сплошная
кривая)
и нарастающих по амплитуде сигналов
(штриховая
кривая).
Постоянная времени сигналов ( т) для 1
—
70 мсек., для 2
—
8 мсек. 1, 2
—
то же, что и для А.
На Ап
Б по оси абсцисс —
частота сигнала в кгц, по
оси ординат —
интенсивность в дб относительно уровня
0.0002 дин/см*
2.
ние
группы, а также наличию ряда промежуточных
групп.
Тормозные
процессы проявляются в виде тормозных
пауз в узоре вызванного ответа на звук,
а также в виде полного или частичного
подавления спонтанной активности в
период действия сигнала или после его
выключения. Увеличение интенсивности
и длительности раздражителя в ряде
случаев приводит к уменьшению
количества импульсов в ответе и к
увеличению скрытого периода реакции.
Особенно отчетливо торможение проявляется
при действии двух звуков: реакция на
тон “оптимальной частоты может быть
полностью заторможена на фоне тонов
неоптимальной частоты. При этом
«тормозные» частоты располагаются по
обеим
16*
243
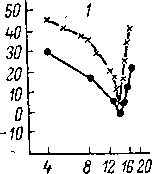

сторонам
от оптимальной частоты, частично вне
зоны ответа нейрона, частично перекрывая
ее. Сложный характер возбуждающих
и тормозных процессов был показан при
внутриклеточном отведении от нейронов
задних холмов (Nelson, Erulkar, 1963). Взаимодействием
возбуждения и торможения, по-видимому,
определяется высокая специфичность
реакции на сложные звуковые сигналы
(Suga 1965а), модулированные по частоте и
амплитуде (Nelson et al., 1966; Вартанян, 1969).
Например, такой признак частотно-модулированного
сигнала, как направление изменения
частоты в сигнале, имеет определяющее
значение для возникновения реакции
многих нейронов задних холмов.
Нейроны
задних холмов характеризуются
постоянством характеристик импульсной
активности. Лишь для небольшой части
нейронов (около 3%) обнаружено угасание
реакции при повторном применении
раздражителя (явление «привыкания»).
В отдельных случаях при многократном
применении сигнала отмечалось небольшое
понижение порога ответа нейрона. Однако
некоторые формы сложного поведения
нейронов обнаружены уже на этом уровне
слуховой системы. Это — «альтернирующий»
тип реакции (Альтман, 1966), отражение в
разряде последействия некоторых свойств
раздражителя (Гершуни и Вартанян, 1969),
которые могут быть отнесены к проявлениям
кратковременной памяти на уровне
одиночных нейронов. К сложным формам
поведения нейронов может быть отнесена
высокая специфичность реакции лишь к
определенным признакам звука:
направлению изменения частоты в сигнале
при частотной модуляции, частоте
амплитудной модуляции, направлению
движения источника звука в горизонтальной
плоскости (Suga, 1965а, 19656; Вартанян, 1968;
Альтман, 1968, 1969).
Внутреннее
коленчатое тело
Как
уже указывалось выше (стр. 202), основным
коллектором афферентных путей, идущих
от среднемозгового центра слуховой
системы (задних холмов) к ее таламическому
уровню (внутреннему коленчатому телу),
является ручка заднего холма.
По
сравнению с задними холмами на уровне
внутреннего коленчатого тела
наблюдается значительное увеличение
количества спонтанно-активных нейронов,
число которых достигает 75% от общего
количества обследованных элементов.
Спонтанная активность достаточно
редкая (как правило, 0.5—10 имп./сек.) и
нерегулярная. Еще одной особенностью
спонтанной активности ряда нейронов
внутреннего коленчатого тела является
ее группирование в пачки (2—5 импульсов
в пачке), которые повторяются 5—10
раз в течение 2—3 сек. Интервалы между
этими сериями пачек нерегулярны и
длятся от нескольких до 10—15 сек.
Вызванная
активность нейронов внутреннего
коленчатого
244
тела,
возникающая при действии тональных
сигналов, длящихся 50—250 мсек.,
характеризуется теми же типами узора
разряда, которые были описаны при
рассмотрении нижележащих отделов
слуховой системы: одиночный импульс,
возникающий при включении (либо при
включении и выключении) сигнала;
«пачковый» разряд (3—6 импульсов в
пачке), возникающий при включении
сигнала; начальный разряд, состоящий
из одного или нескольких импульсов, за
которым после паузы (10—50 мсек.) следует
разряд, длящийся все время действия
звука; тонический разряд, протекающий
в течение всего времени действия
звукового сигнала.
На
уровне внутреннего коленчатого тела
по сравнению с задними холмами
наблюдается увеличение количества
нейронов, реагирующих только на
включение звукового сигнала и уменьшение
количества элементов, обладающих
тоническими узорами разряда. Так, по
данным Маруяма с соавторами (Maruyama et
al., 1966), количество нейронов, отвечающих
только на включение сигнала одиночным
импульсом или пачкой импульсов, достигает
80% от общего числа обследованных
элементов.
Относительно
изменений импульсной активности
нейронов внутреннего коленчатого тела
при действии длительных (секунды)
звуковых сигналов имеются лишь отдельные
наблюдения,- свидетельствующие о
том, что уменьшение частоты импульсации
продолжается в течение 6—8 сек. после
включения звука, после чего импульсация
устанавливается на постоянном, более
низком, чем исходный, уровне (Gross, Thurlow,
1951).
При
действии коротких звуковых щелчков
реакция нейронов внутреннего коленчатого
тела, как правило, характеризуется
одним или несколькими импульсами,
возникающими непосредственно после
действия стимула. Однако у ряда нейронов
при этом возникает еще один своеобразный
тип реакции, не наблюдавшийся в
нижележащих отделах слуховой системы
и получивший название «реверберирующего»
разряда. У этих нейронов в ответ на
щелчок возникает уже описанный выше
начальный разряд, состоящий из одного
или нескольких импульсов, вслед за
которым после большой паузы (0.2—1.0 сек.)
следует несколько пачек импульсов,
длящихся 1—5 сек. Реверберирующий тип
разряда наблюдается у ряда нейронов и
при отсутствии начального разряда,
возникающего непосредственно после
действия щелчка (рис. 88, /, В).
Следует указать, что реверберирующий
тип разряда возникает не только при
звуковом раздражении, но и при
электрическом раздражении ножки заднего
холма (Aitkin, Dunlop, 1968). По всей вероятности,
реверберирующий разряд имеет отношение
к организации краткосрочной слуховой
памяти.
При
исследовании нейронов внутреннего
коленчатого тела в условиях действия
тональных сигналов различной частоты
получены частотнопороговые кривые
самого различного вида — узкие, широкие,
замкнутые, с несколькими максимумами
чув-
245
ствительности
(рис. 88, III).
Оптимальные частоты различных нейронов
внутреннего коленчатого тела отличаются
между собой и покрывают примерно тот
же диапазон частот, что и в нижележащих
отделах слуховой системы. Следует
указать, что тоно-
1сек. 1сек. 1сек.
100мкв\
_
^20
мсек. 50 мсек.
Рис.
88. Типы разрядов (I, II)
и частотнопороговые кривые (III)
нейронов
внутреннего коленчатого тела.
I
—
постстимульные гистограммы ответов
трех нейронов (А, Б, В) на щелчок (по:
Альтман и др., 1970). А
—
нейрон с начальным разрядом; Б — нейрон
с начальным разрядом и послеразрядом;
В — нейрон с одним послеразрядом. N
—
число импульсов. II
—
«пачковый» тип ответа двух нейронов
(А, Б) на тональный сигнал (по: Adrian
Lifschitz et aL, 1966)* A — ответ на один и тот же
сигнал (показан внизу);
Б —
ответы на сигнал различной интенсивности
(цифры
слева —
затухание относительно максимального
уровня дб, снятые методом «точек» (см.
пояснения к рис. 84)* III
—
частотнопороговые кривые для двух
нейронов (А, Б) (по: Maruya- ma et al., 1966).
Обозначения те же, что и на рис. 78.
топическая
локализация к настоящему времени во
внутреннем коленчатом теле не обнаружена.
Подробные
данные о зависимости числа импульсов
в реакции нейронов внутреннего
коленчатого тела от интенсивности
звуковых сигналов оптимальных и
неоптимальных частот в лите
246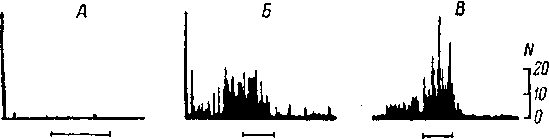
![]()
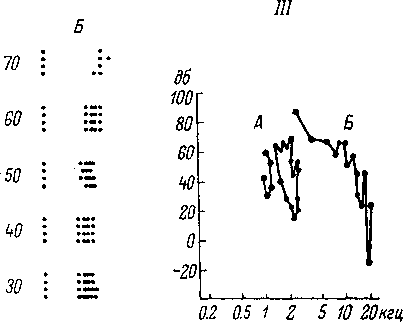
ратуре
не представлены. Имеющиеся в этом
отношении данные, полученные, как
правило, при использовании звуков
оптимальной частоты, свидетельствуют
о том,что кривые указанной зависимости
у нейронов внутреннего коленчатого
тела достаточно пологи (Katsuki et al., 1959).
Что
касается скрытого периода реакции (при
стимуляции тональными посылками
оптимальной частоты или короткими
щелчками), то при средних надпороговых
интенсивностях стимула скрытый период
колеблется в достаточно широких пределах
— от 5 мсек, до нескольких секунд; однако
у основной массы обследованных
нейронов скрытый период группируется
вокруг значений 5—30 мсек.; величины
порядка 30—100 мсек, встречаются значительно
реже; нейроны, обладающие скрытым
периодом больше 100 мсек., единичны.
Отсутствие данных, характеризующих
импульсную активность нейронов
внутреннего коленчатого при действии
сигналов разной длительности, не
позволяет проводить классификацию
этих нейронов по критериям, которые
были использованы при рассмотрении
нейронных популяций ряда нижележащих
отделов слуховой системы.
Тормозные
влияния на уровне внутреннего коленчатого
тела достаточно хорошо выражены, что
проявляется во временном структурировании
разряда, наличии немонотонных кривых
зависимости числа импульсов от
интенсивности стимула, а также в явлении
маскировки тона шумом или щелчка тоном.
Кроме того, тормозные влияния на нейроны
внутреннего коленчатого тела обнаружены
при раздражении зоны AI слуховой области
коры (Watanabe et al., 1966; Aitkin, Dunlop, 1969).
Данные,
характеризующие импульсную активность
нейронов внутреннего коленчатого тела,
представлены и в других исследованиях
(Galambos, 1952; Galambos et al., 1952; Vernier, Galam- bos, 1957;
Adrian, Lifschitz et al., 1966; Aitkin et al., 1966).
Слуховая
кора
Основная
часть работ, посвященных исследованию
нейрональной активности слуховой
области коры, проведена на первичной
зоне (AI) слуховой коры, которая получает
прямые восходящие связи от всех слуховых
ядер таламического уровня (Rose,
Woolsey, 1958). Большинство авторов, работавших
как на анестезированных препаратах,
так и на неанастезирован- ных
необездвиженных животных, отмечают
непостоянство ответов от раздражения
к раздражению, быструю их адаптацию и
значительный процент нейронов, не
реагирующих на звук (20—34% по данным
разных авторов) (Erulkar et al., 1956; Katsuki et al.,
1959; Evans, Whitfield, 1964; Вардапетян, 1967a, 19676).
Только в одной работе на обездвиженных
флакседилом кошках отмечается всего
3% нейронов, нечувствительных к звуковому
247
раздражителю,
и подчеркивается относительная
стабильность разряда нейронов по
сравнению с данными других авторов
(Goldstein et al., 1968). Основные отличия в
результатах исследований вряд ли
определяются особенностями условий
острого или хронического экспериментов,
различиями методик отведения активности
(металлические или стеклянные
микроэлектроды) или сдавливанием
исследуемой зоны в результате
экспериментальной процедуры. Скорее
это различие является результатом
отсутствия единых критериев оценки
«вызванного» звуком ответа. В большинстве
работ ответ нейрона рассматривается
как импульсная реакция, возникающая
на включение, выключение или продолжающаяся
в период действия звукового сигнала.
Именно в этом случае обнаруживается
значительный процент нейронов,
нечувствительных к звуковому раздражителю.
В тех случаях, когда ответ рассматривается
как любое изменение импульсации по
сравнению с исходным уровнем спонтанной
активности (Goldstein et al., 1968; Вартанян и
Шмигидина, 1971) даже через значительные
промежутки времени после нанесения
раздражителя, большинство нейронов
области AI могут быть отнесены к
звукочувствительным элементам.
Более того, именно такой подход к
вызванной активности позволил
выделить в коре группу нейронов с
длительными послеразрядами, в которых
даже через секунды после нанесения
короткого раздражителя (длительностью
несколько миллисекунд) отражаются
некоторые свойства действовавшего
сигнала (Вардапетян, 19676; Альтман, 1971).
Поскольку кора является конечным звеном
слуховой системы и высшим интегративным
центром, естественно возникает
предположение, что деятельность
корковых нейронов обеспечивает более
сложные формы слухового поведения, чем
распознавание таких простых свойств
звука, как интенсивность и частота, и
связана с выделением и анализом более
сложных признаков сигнала, в особенности
его временной структуры (см. стр. 268).
Поэтому классификация по тем же
свойствам, которые принимаются для
нейронов нижележащих отделов, вряд
ли применима для нейронов слуховой
коры.
Исследования
функциональной организации слуховой
корковой зоны включают следующие
вопросы: 1) пространственное
представительство звуковых частот по
поверхности слуховой коры (тонотопическая
локализация); 2) организация клеток с
одинаковыми функциональными
свойствами в виде вертикальных столбов
(колумнарное строение); 3) функциональная
организация клеток, соответствующая
анатомически определенным слоям коры
(ламинарная организация); 4) общие
характеристики импульсной активности
нейронов. Рассматриваются как частотно
избирательные свойства нейронов
(оптимальная частота, ширина и форма
частотнопороговых кривых), так и общие
характеристики разрядов нейронов
на различные звуковые сигналы — щелчки,
тоны, шум, а также делаются попытки
классифицировать кор
248
ковые
нейроны по критериям специфичности
реакций на звуки, модулированные по
частоте (Suga, 1965а,b; Evans, 1968) и по стабильности
ответов на повторяющиеся раздражители
(Варда- петян, 1967а), т. е. по критериям,
отличным от применяемых для нижележащих
отделов.
Частотно-пространственная
организация. Представление о
пространственном представительстве
периферического отдела слуховой
системы — улитки в слуховой коре было
положено работами Вулси и Уолцла
(Woolsey, Walzl, 1942). Они показали, что при
электрической стимуляции волокон
кохлеарного нерва, идущих от апикального
конца улитки, ответы возникают в задней
части средней эктосильвиевой извилины
(обозначенной как зона AI); раздражение
волокон, связанных с основанием
улитки, вызывает ответы в передней
части зоны AI. В области АП, располагающейся
параллельно и вентрально по отношению
к средней эктосильвиевой извилине
(рис. 66), имеется сходное, но противоположно
направленное пространственное
представительство улитки: ответы в
передней части этой зоны возникали
при раздражении верхушки, в задней
части — при раздражении основания
улитки. В соответствии с данными Вулси
(Woolsey, 1961) в слуховой области коры имеется
по крайней мере четыре полных
представительства улитки: в AI, АП, Ер
и супрасильвиевой извилине.
С
помощью методики регистрации и оценки
порогов вызванных стрихнинных
спайков Тунтури (Tunturi, 1944, 1952, 1960) показал
наличие пространственного распределения
ответов на различные звуковые частоты
вдоль слуховой зоны AI собаки (рис. 89,
А).
Реакция на тоны низкой частоты
обнаруживается в задней части средней
эктосильвиевой извилины, реакция на
тоны высокой частоты — в передней
части. Вся кора может быть разделена
на полосы шириной 2 мм, каждая из них
соответствует частотам, различающимся
на одну октаву. Хинд (Hind, 1953) подтвердил
данные Тунтури на кошках, используя
сходную методику локальной
стрихнинизации коры. Однако столь
четкого распределения ответов, как у
Тунтури, им не было получено (рис. 89, Б).
Хиндом были обнаружены две низкочастотные
зоны: одна в задней части AI, другая в
Ер, тогда как частоты выше 30 кгц
представлены в передней части AL Зоны
представительства промежуточных
частот (10—30 кгц) перекрываются. Результаты
оценки частотной локализации в коре
кошки по вызванным потенциалам
(Качуро, 1963) показали еще большую
сглаженность этого явления по сравнению
с исследованиями, проведёнными с
использованием локальной стрихнинизации
коры (рис. 89, В).
В
этом случае относительно острая
локализация характерна только для
высоких частот (разница в порогах около
30 дб) в передней части области AI.
Сомнение
в существовании точного пространственного
рас-
249
пределения
нейронов, чувствительных к различным
звуковым частотам, было впервые высказано
Эрулькаром и соавт. (Erulkar et al., 1956) при
исследовании отдельных элементов зоны
AI. Эванс и др. (Evans et al., 1965) показали, что
вероятность обнаружения единиц,
характеристичная частота которых ниже
10 кгц, одинакова вдоль всей зоны AL
Элементы с высокими характеристичными
частотами (выше 10 кгц) сосредоточены в
узкой области в передней части AI. В
задней части AI было зарегистрировано
Рис.
89. Частотная локализация в слуховой
коре (X — по: Tunturi, 1950;
Б
—
по: Hind, 1953; В
— по: Качуро, 1963; Г —по: Goldstein et al., 1970).
На
А
—
данные получены путем оценки порогов
вызванного звуком стрихниного спайка
после локальной стрихнинизации слуховой
коры собаки; цифры
—
частота тонов в гц; Левое полушарие. На
Б — распределение оптимальных частот
</0.
кгц) по поверхности стрихнинизированной
слуховой коры кошки: Величина /0
указана под каждойдсхемой. На В
—
частотная локализация в слуховой коре
кошки, полу- ченная^на основании оценки
порогов вызванных потенциалов на тоны
в состоянии наркозами бодрствования;
цифры
—
оптимальные частоты (кгц) в указанных
точках. На Г
—распределение
одиночных нейронов в слуховой коре
кошки, максимально чувствительных к
частотам (/0,
кгц), указанным под каждой схемой.
скопление
нейронов, чувствительных к низким
частотам (Goldstein et al., 1970; рис. 89).
Совершенно
очевидно, что тонотопическая организация
слуховой корковой зоны, полученная
с помощью стрихнинной методики,
значительно более выражена, чем при
оценке вызванных потенциалов и ответов
одиночных нейронов. По-видимому, при
действии стрихнина возникают особые
условия для проявления закономерной
частотной локализации. Тонотопическая
организация
250


слуховой
коры рассматривается как функциональная
основа частотного различения (см.,
например: Granit, 1956). Поскольку, однако,
при исследовании одиночных нейронов
она проявляется очень слабо, многие
авторы приходят к заключению, что
тоното
дб
110
701
110
70
110
80
110
90
100
70
200мкм
Рис.
90. Частотнопороговые кривые слуховых
корковых нейронов летучей мыши (по:
Suga, 1965).
Все
кривые получены при одном погружении
микроэлектрода от поверхности коры.
Цифры
справа —
глубина погружения от поверхности
коры в мкм. По
оси абсцисс —
частота сигнала в кгц; по
оси ординат — интенсивность
в дб.
мм
О-
1-
^100^
Рис.
91. Частотные зоны ответов нейронов
слуховой зоны коры кошки, полученные
при четырех погружениях микроэлектрода
(по Goldstein et al., 1970).
По
оси абсцисс —
частота сигналов в кгц; по
оси ординат —
глубина погружения электрода от
поверхности в мм. Горизонтальные
линии —
ширина частотной зоны ответа нейронов,
расположенных на различной глубине.
пическая
локализация нейронов в коре не проявляется
в той степени, которая может иметь
существенное функциональное значение
для частотного анализа, наличие же
некоторой локализации определяется
проекционными связями от тонотопически
организованных нижележащих центров.
Карты частотной локализации,
251![]()
![]()
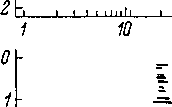
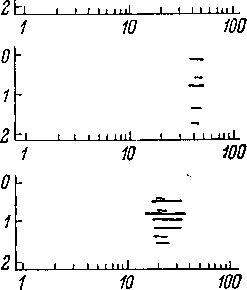
полученные
при оценке стрихнинного спайка и
вызванного ответа, могут рассматриваться
как явление, характеризующее лишь
ограниченный промежуток времени
действия сигнала и отражающее реакцию
слуховой коры на включение сигнала.
Так, например, время, предшествующее
развитию указанных электрических
явлений, составляет величину порядка
20—40 мсек. Действительно, удаление
слуховых корковых зон не ухудшает
различения интенсивности и частоты
длительно действующего тона (Roab, Ades,
1946; Neff, 1961; Бару, 1964). В то же время удаление
слуховых корковых зон вызывает нарушение
различения временной последовательности
звуков (см. стр. 277, 410), дефицит способности
локализовать источник звука в
пространстве (см. стр. 278), а также
заметное ухудшение различения и
обнаружения коротких сигналов (см. стр.
274).
Колумнарная
организация. Возможность
структурно-функциональной организации
корковых клеток в виде вертикальных
столбов вытекает из данных Лоренте де
Но (Lo- rente de No, 1933b, 1949), который показал,
что синаптические соединения корковых
нейронов объединяют клетки разных
слоев в вертикальные цепочки без
широкого горизонтального ветвления.
Определенная функциональная организация
вертикальных столбов клеток была
установлена в сенсо-моторной (Mountcastle,
1957; Powell, Mountcastle, 1959) и в зрительной коре
(Hubei, Wiesel, 1959, 1963). Эти клетки имеют общие
свойства: одинаковую сенсорную
модельность (в сенсо-моторной коре)
или одинаковые рецептивные поля (в
зрительной коре).
Предположение
о возможности колумнарного строения
слуховой коры было впервые выдвинуто
Хиндом и др.. (Hind et al., 1960) на основании
данных о том, что в одном вертикальном
проколе микроэлектродом через толщу
коры клетки имеют одинаковые или
близкие между собой характеристичные
частоты. Впоследствии появились
работы, показывающие, что слуховая кора
может быть разбита на множество
вертикальных столбов, организация
которых определяется характеристичной
частотой (рис. 90). По данным Суга (Suga,
1956b), характеристичные частоты нейронов
слуховой коры летучих мышей при
вертикальном погружении микроэлектрода
отличаются не более, чем на 0.1 октавы,
а пороги этих нейронов близки
(среднеквадратическое отклонение
составляет 8.5 дб). В наиболее подробной
работе (Abeles, Goldstein, 1970) было убедительно
показано, что нейроны, расположенные
вдоль вертикальной цепочки, во многих
случаях имеют не только одинаковые
оптимальные частоты, но и очень близкие
частотные зоны ответов (рис. 91). Временой
узор разряда и количество импульсов в
ответе нейронов, входящих в состав
вертикальной цепочки, резко различны
при максимальной чувствительности к
одинаковой характеристичной частоте
(Gerstein, Kiang, 1964; Abeles, Goldstein, 1970).
252
Ламинарная
организация. Попытки уста-
новить
принадлежность к определенному слою
слуховой коры
клеток, реагирующих
только на определенные звуковые
сигналы
(щелчок, тоны, шум) или
обладающих одинаковыми свойствами,
пока
не увенчались успехом (Abeles, Goldstein, 1970).
Известно,
что в зрительной системе клетки с разной
степенью
сложности рецептивных
полей относятся к различным слоям зри-
тельной
коры: IV слой клеток
содержит большее
количество
элементов с простыми
рецептив-
ными полями, тогда как
более
поверхностные слои включают
клетки
со сложными рецептив-
ными полями
(Hubei, Wiesel,
1962). По данным Ониши и
Кацуки
(Oonishi, Katsuki, 1965),
форма частотнопороговой
кри-
вой различна для нейронов,
расположенных
в различных сло-
ях слуховой коры.
В
соответствии с частотной
избирательностью
нейроны слу-
ховой коры отличаются
по харак-
теру частотнопороговых
кривых
(рис. 92). Частотнопороговые
кри-
вые, как и в других отделах
слуховой
системы, разделяются
по форме и
остроте на острые
Рис.
92. Частотнопороговые кривые
(ЧПК)
нейронов слуховой зоны коры
кошки
(по: Abeles, Goldstein, 1970).
А
—
острые ЧПК, В
—
широкие, В
—
муль-
типиковые. По
оси абсцисс —
частота сигнала
в кгц; по
оси ординат —
интенсивность в дб
над уровнем 0.0002
дин/см2.
(рис.
92, А)
пологие, или широкие (рис. 92, Б),
и мультипиковые (рис. 92, В).
Отношения по частотной шкале между
максимумами чувствительности в
мультипиковых частотнопороговых кривых
чаще всего близки к октавным (1 или 2
октавы). Максимальная чувствительность
регистрируется, как правило, для наиболее
высокой из характеристичных частот.
Для нейронов, расположенных в
поверхностных слоях, характерна плоская
частотнопороговая кривая, в более
глубоких слоях чаще встречаются нейроны
с узкой или мультипиковой частотнопороговой
кривой. Катцуки предполагает, что
мультипиковая частотнопороговая кривая
ха-
253
рактерна
для нейронов, на которых происходит
конвергенция клеток с одиночной
ответной зоной, нейроны с мультипиковыми
частотнопороговыми кривыми в свою
очередь конвергируют на нейронах более
поверхностных слоев коры с широкими и
плоскими частотнопороговыми кривыми.
Действительно, клетки с мультипиковыми
частотнопороговыми кривыми обнаружены
на большей глубине, чем клетки с широкими
ответными зонами (Oonishi, Katsuki, 1965).
Общие
характеристики импульсной активности
нейронов. Более 90% нейронов зоны AI
обладают спонтанной активностью,
частота которой варьирует от 1 до 35
имп./сек. В редких случаях частота
спонтанной активности превышает
35 имп./сек. Спонтанная активность
нерегулярна и в некоторых случаях
группируется в пачки.
При
действии шумовых и тональных сигналов
возникают ответы на включение
(on-эффект), выключение (off-эффект),
включение—выключение (on—off-эффект)
и разряд, продолжающийся в течение
всего времени действия звука. На рис.
93 представлены основные варианты
разрядов корковых нейронов на звук. У
некоторых нейронов отмечается
торможение спонтанной активности на
звук.
При
действии коротких звуковых сигналов
— щелчков описаны следующие импульсные
реакции: 1) ответ на включение раздражителя,
состоящий из 1—3 импульсов; такие ответы
могут быть зарегистрированы у различных
нейронов, независимо от уровня их
спонтанной активности; 2) торможение
спонтанной активности; такие ответы
могут состоять из начального разряда
с последующим торможением спонтанной
активности или из начального торможения
спонтанной активности с последующим
ее восстановлением до исходного уровня.
Интересно отметить, что во всех
нижележащих отделах слуховой системы
торможение спонтанной активности
нейрона при действии звука можно вызвать
при определенном подборе частоты и
интенсивности сигнала, тогда как в
слуховой коре нейроны с тормозными
реакциями составляют отдельную группу
в отличие от нейронов с возбуждающими
реакциями (Evans, Whitfield, 1964). Все указании
е выше варианты ответов были описаны
также для нейронов всех нижележащих
отделов слуховой системы. Характерным
свойством определенной части корковых
нейронов является длительный послеразряд,
который возникает через секунды после
начального ответа на включение сигнала
(Вардапетян, 19676; Альтман, 1970). Такие
реакции нейронов впервые появляются
лишь в медиальном коленчатом теле (рис.
88 Б,
В),
Количество импульсов в разряде
корковых нейронов при увеличении
интенсивности возрастает до определенной
величины, после чего количество
импульсов в реакции сохраняется на
постоянном уровне, либо наблюдается
уменьшение импульсации. В некоторых
случаях
254
максимальное
количество импульсов в ответе корковых
нейронов возникает в очень узком
диапазоне интенсивностей звукового
раздражителя (Brugge et al., 1969).
Скрытые
периоды ответов нейронов зоны AI
колеблются от 7 до 250 мсек, для разных
элементов. Нейроны могут быть разделены
на три группы в соответствии с величиной
скрытого периода ответа на звук:
7—15; 15—40; 84—250 мсек. (Erulkar et al., 1956; Серков,
Сторожук, 1969).
Рис.
93. Типы ответов нейронов слуховой зоны
коры кошки на звуковые сигналы (по:
Evans, Whitfield, 1964).
А
—
длительный разряд; Б
—
торможение при действии звука;
В
—
ответ на включение; Г
—
ответ на выключение; Д
—
ответ на
включение и выключение; Е
—
ответ, возникающий только при
действии
частотно-модулированного сигнала. На
нижнем луче —
форма
звуковых сигналов. Масштаб времени 500
мсек.
Тормозные
процессы в корковых нейронах проявляются
в виде пауз в импульсных реакциях
нейронов на звук, полного подавления
спонтанной активности, резкого
ограничения диапазона частот и
интенсивностей, которые вызывают
реакции нейронов. При стимуляции двумя
тонами наблюдается как тормозные, так
и облегчающие эффекты, выражающиеся в
уменьшении или увеличении импульсации.
Тормозные эффекты чаще отмечались у
нейронов с широкими и плоскими
частотнопороговыми кривыми, эффекты
облегчения — у нейронов с одно- или
мультипиковой частотнопороговой кривой
и не только тогда, когда частоты стимулов
были близки между собой и давали биения,
но и отдален
255
ные
по частотной шкале. Возможно, эффект
облегчения, обнаруженный при
стимуляции двумя тонами, имеет
существенное значение для различения
сложных звуков.
В
значительной части случаев реакция
нейронов на стационарные сигналы
отсутствует, а появляется лишь в условиях
действия звуков с изменяющимися во
времени параметрами, например, на
модулированные по частоте (ЧМ) звуки
(Suga, 1965b; Whitfield, Evans, 1965; Evans, 1968). На основании
оценки ответов на ЧМ — сигналы эти
авторы предлагают следующее деление
нейронов на группы:
Нейроны,
реагирующие на ЧМ-сигналы независимо
от направления изменения частоты
сигнала.
Нейроны,
реагирующие на ЧМ-сигналы только при
определенном направлении изменения
частоты сигнала (восходящем или
нисходящем). В некоторых случаях
достаточно очень малого диапазона
изменения частоты, порядка полутона
музыкальной шкалы, для того чтобы
вызвать реакцию нейронов, не реагирующих
на чистые тоны. По-видимому, эта группа
специализированных нейронов,
регистрирующих наличие такого признака
звука, как направление изменения
частоты.
Нейроны,
реагирующие только на ЧМ-сигналы, но
не реагирующие на чистые тоны. По
Эвансу (Evans, 1968) в слуховой коре кошки
таких нейронов около 20%.
Нейроны,
не реагирующие на ЧМ-сигналы. Обычно
это нейроны, которые возбуждаются
только очень узким диапазоном чистых
тонов.
В
отличие от стволовых отделов слухового
пути для нейронов зоны AI характерна
нерегулярность и значительная
изменчивость ответов при повторных
раздражениях. В ряде случаев наблюдалось
быстрое «привыкание» к раздражителю
после 2— 3-го применений. В других случаях
ответ, который при первых применениях
раздражителя был нерегулярным и
изменчивым, стабилизировался при
многократном применении сигнала.
Нестабильность реакций значительной
части корковых нейронов отмечалась
как на анестезированных препаратах
(Erulkar et al., 1956; Вардапетян, 1967а, 19676), так
и на неанестезированных и необездвиженных
животных (Katsuki et al., 1959; Bogdanski, Galambos, 1960;
Evans, Whitfield, 1964, и др.). Во многих случаях
после звукового раздражения наблюдаются
изменения спонтанной активности в
виде перераспределения импульсов в
пачках или изменения плотности
импульсации (Goldstein et al., 1968). Для многих
нейронов характерен своеобразный
поздний разряд возникающий через
секунды после нанесения раздражителя
(Альтман, 1971), а также значительная
длительность реакции нейрона, во много
раз превышающая длительность сигнала
(Вардапетян, 1967а). Описаны
«мультимодальные» нейроны, отвечающие
не только на щелчки, но и на движение
предмета в зри
256
тельном
поле Evans, Whitfield, 1964). Интересно отметить
своеобразие «нейронов внимания»
(Hubei et al., 1959), реакция которых возникает
только во время рассматривания животным
звучащего объекта.
Как
уже отмечалось, некоторые формы сложного
поведения характерны уже для нейронов
нижележащих отделов слуховой системы.
Возможно, своеобразие и специфика
коркового уровня определяется не
качественным, а количественным
соотношением нейронов с различными
функциональными свойствами и сложными
формами поведения.
О
ПРЕОБРАЗОВАНИИ ХАРАКТЕРИСТИК АФФЕРЕНТНОГО
ПОТОКА НА РАЗНЫХ УРОВНЯХ СЛУХОВОЙ
СИСТЕМЫ
Сопоставление
характеристик электрической активности
нейронов разных отделов слуховой
системы, рассмотренных выше, позволяет
проследить некоторые тенденции
преобразования афферентного потока
импульсации при прохождении его через
последовательные уровни синаптических
переключений в слуховой системе.
Как
следует из представленного материала,
число спонтанноактивных нейронов
постепенно понижается при переходе от
волокон слухового нерва через
кохлеарные ядра, вернеоливарный комплекс
к задним холмам. При этом происходит
также уменьшение частоты спонтанной
импульсации нейронов. Однако на уровне
медиального коленчатого тела и слуховой
области коры процент спонтанноактивных
нейронов вновь возрастает. Высокий
уровень спонтанной активности
значительной части нейронов нижних
уровней слуховой системы (волокон
слухового нерва, кохлеарных ядер и
верхней оливы) обеспечивает возможность
временного контрастирования ответа
нейронов на звуковой сигнал, т. е.
отделения его во времени от фоновой
активности благодаря выраженному
подавлению спонтанной импульсации
непосредственно после импульсной
реакции нейрона на звук (см., например,
рис. 71). Тем самым создаются благоприятные
условия для обнаружения слабых сигналов.
Характерным для спонтанной активности
вышележащих отделов слуховой системы
является ее нерегулярность и способность
группироваться в пачки, а также выраженные
изменения ее временной структуры,
которые могут наблюдаться через
длительные (порядка нескольких сек.)
промежутки времени после выключения
звука. Все это позволяет предполагать,
что изменения спонтанной активности
в верхних отделах слуховой системы
могут быть проявлением длительных
следов воздействия звуков в цепях
нейронов, являясь одним из факторов,
способствующих организации краткосрочной
слуховой памяти.
17
Сенсорные системы
257
Узор
импульсного разряда нейронов становится
значительно более многообразным и
сложным по мере повышения уровня (ср.
рис. 71, 76, 81, 84, 88, 93) слуховой системы.
Простой тонический узор разряда,
характерный для волокон слухового
нерва, уже не является универсальным
для нейронов кохлеарных ядер, где
встречаются также нейроны с тоническим
узором разряда, временная структура
которого осложнена тормозными паузами
различной продолжительности и различного
положения во времени относительно
момента начала сигнала. Кроме того, для
центральных отделов слуховой системы
типично наличие нейронов с фазным
и «пачковым» узором разряда. Количество
нейронов с фазным узором разряда
при действии тона оптимальной частоты
и достаточно малого уровня интенсивности
в кохлеарных ядрах составляет менее
20%, в ядрах верхнеоливарного комплекса
их процент также невелик, однако в
нижних холмах нейроны с фазным узором
разряда составляют около 50%, причем
фазный узор разряда наблюдается в
значительно большем диапазоне
варьирования параметров стимула, чем
на уровне кохлеарных ядер.
Насколько
можно судить по имеющимся данным, на
уровне медиального коленчатого тела
и слуховой коры, процент нейронов с
фазным разрядом вновь несколько
понижается за счет возрастания
процента нейронов с «пачковым» и
тоническим узором разряда. «Пачковый»
узор разряда, который крайне редко
наблюдается на уровне кохлеарных ядер
и несколько чаще — в нижних холмах,
является весьма характерным для высших
отделов слуховой системы и особенно
для нейронов медиального коленчатого
тела (рис. 88, II).
Таким образом, с повышением уровня
слуховой системы в целом уменьшается
число импульсов в реакции нейронов па
звук, а также соответствие характеристик
импульсной реакции нейронов параметрам
звукового стимула: если на нижних
уровнях слуховой системы частота
разряда большей части нейронов в
некоторых пределах соответствует
интенсивности звука, длительность
разряда соответствует длительности
звука, а характер постстимульных
гистограмм отражает форму огибающей
звукового стимула, то на верхних уровнях
слуховой системы такое соответствие
выражено в слабой степени или вообще
отсутствует (ср. рис. 71, 76, 81, 84, 88, 93);
уменьшается также способность
воспроизведения ритма стимуляции
в суммарной активности (табл. 7).
Кроме
того, уже на уровне кохлеарных ядер
появляются и затем все более возрастают
в числе нейроны, обнаруживающие
относительно сложное поведение. К таким
сложным формам поведения могут быть
отнесены нестабильность возникновения
ответа на звук, привыкание к действию
раздражителя, некоторые виды памяти о
свойствах сигнала, а также избирательность
реакции нейронов к определенным
параметрам стимула (в частности, к
определенному диапазону интенсивности
и длительности звука,
258
к
направлению модуляции частоты в
частотно-модулированиых сигналах).
Существенным,
по-видимому, является также резкое
возрастание вариабельности различных
характеристик разряда нейронов на звук
с постоянными параметрами: как уже
отмечалось, по мере повышения уровня
слуховой системы увеличивается дисперсия
величин скрытого периода импульсов и
их числа в ответе на звуковой сигнал;
возрастает также диапазон флуктуации
порога реакции нейронов. Эти изменения,
по-видимому, лежат в основе десинхронизации
афферентного потока импульсов по мере
его прохождения от нижних к верхним
отделам слуховой системы. При этом,
однако, возрастает устойчивость
суммарных ответов и импульсной активности
нейронов к таким десинхронизирующим
факторам, как пологий фронт нарастания
сигнала и маскировка.
Частотнопороговые
кривые (ЧПК), характеризующие частотную
избирательность нейронов, обнаруживают
тенденцию к усложнению своей формы
по мере перехода от нижних к верхним
уровням слуховой системы (ср. рис.
72, 78, 86, 87, 88, 92). Это усложнение
проявляется в виде дополнительных
пиков на ЧПК, в замкнутости частотной
области ответа, в распределении области
ответа на несколько не связанных между
собой зон. По-видимому, усложнение формы
ЧПК отражает усложнение конвергенции
импульсации на нейронах верхних
отделов слуховой системы по сравнению
с нижними. Что касается остроты ЧПК в
области максимальной чувствительности
нейронов, то каких-либо надежных данных
относительно ее изменения при переходе
от нижних уровней слуховой системы к
верхним в литературе нет. Более подробно
вопрос о частотном анализе в центральных
отделах слуховой системы рассмотрен
в специальном разделе (см. стр. 286).
Суммационные
свойства, оцениваемые по суммарным
ответам, постепенно повышаются от
уровня слухового нерва до уровня коры
(табл. 7), что может быть связано с
десинхронизацией потока афферентации
и увеличением длительности суммарных
ответов. Однако данные, полученные на
одиночных нейронах, свидетельствуют
о том, что суммационные свойства
нейронов, оцениваемые в более широком
временном диапазоне, наиболее хорошо
выражены па уровне первого центрального
отдела слуховой системы — кохлеарных
ядер. В нижних холмах способность к
временной суммации в целом выражена
значительно слабее, на уровне коры она
оказывается еще меньше. По-видимому,
накопление во времени снедений о
звуковом сигнале для их последующего
анализа является в основном функцией
нейронов нижних центральных уровней
слуховой системы. Различие результатов
при оценке суммационных свойств по
первичным суммарным ответам и по
активности одиночных нейронов,
очевидно, связано с различием
17*
259
отрезка
времени, в пределах которого производится
такая оценка. Вопрос о значении различных
временных каналов обработки и передачи
информации о звуке рассматривается
ниже (стр. 294).
Все
изложенное позволяет предполагать,
что непосредственное описание
свойств сигнала в характеристиках
импульсной активности нейронов
возможно, как правило, лишь на нижних
уровнях слуховой системы. Активность
нейронов более высоких уровней,
по-видимому, является отражением не
столько параметров звуковых сигналов,
сколько тех операций, которые осуществляет
слуховая система над поступающей
афферентацией. Однако детальное изучение
этих операций и лежащих в основе их
механизмов является задачей будущих
исследований.