

II. Окисление, сопряженное с фосфорилированием адф
Этот тип биологического окисления осуществляется двумя способами.
• Если макроэргическая связь возникает в момент непосредственного окисления субстрата, а затем тем или иным путем передается на фосфатный остаток, который, в свою очередь, используется для фосфорилирования АДФ, т. е. синтеза АТФ, то такой вид биологического окисления называют окислением, сопряженным с фосфорилированием АДФ на уровне субстрата (субстратным фосфорилированием).
Примеры реакций субстратного фосфорилирования:
– при окислении 3-фосфоглицеринового альдегида (3-ФГА) в 2-фосфоглицериновую кислоту (2-ФГК) – см. гликолиз;
– при превращении фосфоенолпировиноградной кислоты (ФЕП) в пировиноградную (пируват, ПВК) – см. там же;
– при превращении -кетоглутаровой кислоты в янтарную (см. реакции цикла Кребса).
• Если атомы водорода с коферментов дегидрогеназ, принимающих участие в окислении субстратов, передаются в оксидоредуктазную цепь, где сопряжено с переносом ионов Н+ и электронов на молекулярный кислород происходит активирование неорганического фосфата и при его посредстве – фосфорилирование АДФ с образованием АТФ, то такое сопряжение окисления с синтезом АТФ называют сопряжением на уровне электронотранспортной (окислительной, оксидоредуктазной, дыхательной) цепи (окислительным фосфорилированием). Понятно, что сам окисляемый субстрат в этом случае непосредственного участия в активировании неорганического фосфата не принимает.
Сопряжение окисления с фосфорилированием идет главным образом на внутренних мембранах митохондрий.
В переносе ионов Н+ и электронов от субстратов к молекулярному кислороду принимают участие следующие окислительно-восстановительные ферменты.
Пиридиновые дегидрогеназы
Коферментами данных ферментов являются НАД и НАДФ. Универсальным донором атомов Н для дыхательной цепи ферментов служит НАДН2. Если при окислении субстрата возникает НАДФН2, то осуществляется реакция:
НАДФН2 + НАД ⇄ НАДФ + НАДН2.
Следовательно, и в этом случае атомы водорода перед поступлением в дыхательную цепь передаются на НАД.
Реакция гидрирования НАД представлена в виде следующего уравнения (полную схему см. выше):

При переносе 2-х атомов водорода от субстрата на НАД+ один из них затем присутствует в восстановленном коферменте в виде атома Н, а другой – в виде электрона. Свободный ион Н+ остается в среде. (Для простоты восстановленные формы НАД и НАДФ изображают: НАДН2 и НАДФН2).
Особенности реакций с участием пиридиновых дегидрогеназ:
Легкая обратимость.
Коферменты легко отделяются от белковой части, обладают высокой подвижностью, что позволяет им переносить атомы Н, ионы Н+ и электроны из одной части клетки в другую.
НАД и НАДФ способны принимать атомы Н от большого числа субстратов, окислительно-восстановительные потенциалы которых ниже (
 ).
).
Флавиновые дегидрогеназы
Коферментами их являются ФМН и ФАД. Флавиновые ферменты являются акцепторами атомов водорода и осуществляют перенос их от НАДН2:
НАДН2 + ФАД ⇄ НАД + ФАДН2.
В некоторых случаях (при окислении янтарной кислоты в цикле Кребса или при окислении жирных кислот) флавиновые ферменты могут играть роль первичных дегидрогеназ, т. е. прямо, без участия НАД, принимать электроны и ионы водорода Н+ от субстратов.
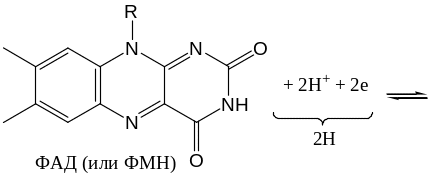
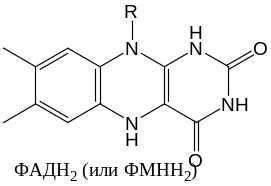
ФМН и ФАД, в отличие от НАД и НАДФ, очень прочно связаны с апоферментом и не отщепляются от него ни на одной стадии каталитического цикла. Активной частью молекул ФАД и ФМН является изоаллоксазиновое кольцо рибофлавина, к атомам азота которого могут присоединяться 2 атома водорода (полную схему см. выше):
Кофермент Q (убихинон)
КоQ – следующий компонент дыхательной цепи. Убихинон может играть роль промежуточного переносчика атомов водорода. Считают, что восстановленная форма флавиновых ферментов в дыхательной цепи передает атомы водорода коферменту Q (см. выше).
Цитохромы
Дальнейший перенос электронов от КоQ на кислород осуществляет система цитохромов, состоящая из ряда гемопротеидов, расположенных в порядке возрастания окислительно-восстановительных потенциалов, что обеспечивает упорядоченную передачу электронов. Цитохромы а и а3 содержат в своем составе еще и атомы меди. При транспорте электронов в направлении увеличения окислительно-восстановительных потенциалов происходят процессы:
Fe2+
![]() Fe3+
(в цит. b,
c1,
с)
Fe3+
(в цит. b,
c1,
с)
Cu+ Cu2+ (в цит. а, а3).
Схема дыхательной цепи
НАДН + Н+ Е = –0,32 В НАД+
![]()
Флавопротеин
ФМН Е = –0,30 В ФМНН2
![]()