

Биология Ярыгин 2003
.pdfвзаимодействия между одинаковыми клетками основываются на антигенных свойствах их мембран.
Избирательная адгезия клеток определенного зародышевого листка друг с другом является необходимым условием нормального развития. Это показано путем сопоставления двух процессов: течения гаструляции в естественных условиях и способности клеток определенного зародышевого листка агрегировать в комплекс с себе подобными. Показано, что если у межвидовых гибридов зародышей амфибий нарушена гаструляция, то диссоциированные в опыте клетки зародышевых листков таких гибридов не способны к агрегации с себе подобными, если же в некоторых случаях гибридам удавалось образовать гаструлу, то и в эксперименте диссоциированные клетки зародышевых листков были способны к реагрегации. Повидимому, в обеспечении сортировки клеток важное место принадлежит генетическим механизмам.
Одним из примером потери клетками способности к избирательной сортировке и слипанию является их беспорядочное поведение в злокачественной опухоли.
Таким образом, сортировка клеток и их избирательная адгезия наряду с другими клеточными процессами играет важную роль в морфогенезе развивающегося зародыша и одновременно подвержена многоуровневым регуляционным воздействиям (генетическим, межклеточным, онтогенетическим), отражая целостность организма как системы.
8.2.4.Гибель клеток
Вразвитии зародышей наряду с размножением клеток важную роль играют
процессы гибели клеток.
Внастоящее время различают два принципиально различных типа клеточной гибели: апоптоз (в переводе с греческого «отпадающий») и некроз.
Апоптоз широко распространен и типичен для физиологических условий. Наряду с описанными выше делением, сортировкой и миграцией клеток, он способствует достижению характерных для определенного биологического вида черт его морфофункциональной организации. Следовательно, апоптоз является естественным, эволюционно обусловленным и генетически контролируемым механизмом морфогенеза. Некроз клеток возникает в нефизиологических условиях, например, в связи с действием неблагоприятных факторов, таких, как стойкое кислородное голодание, разного рода токсины и другие. Некроз обычно сопровождается воспалением и является патологическим процессом. Указанные воззрения в отношении апоптоза и некроза сформировались за последние три десятилетия и поэтому еще нередко некоторые физиологические процессы клеточной гибели описываются термином «некроз» вместо термина «апоптоз».
Наиболее яркие примеры разрушения клеток и органов относятся к постэмбриональным стадиям метаморфоза земноводных и насекомых. У головастиков резорбируются (рассасываются) хвост, кишечник и жаберные крышки,
361
medwedi.ru

у личинок насекомых разрушается большинство внутренних органов. В ходе эмбрионального развития высших позвоночных и человека также имеют место процессы дегенерации органов, которые вначале закладываются, а затем исчезают. У особей женского пола дегенерируют вольфовы протоки, у особей мужского пола—мюллеровы протоки, что является, по-видимому, результатом влияния половых гормонов. У эмбриона человека вначале закладываются ребра у 7-го шейного позвонка и 9—10 хвостовых позвонков, затем они обычно исчезают, так что шейные позвонки, как правило, ребер не несут и в копчике остается 4—5 позвонков (рис. 8.4).
Рис. 8.4. Область клоаки и хвоста у зародышей человека.
362
А —в конце 4-й недели; Б—на 8-й неделе: 1—ректальная область клоаки, 2— уроректальная складка, 3—область мочеполового синуса в клоаке, 4—хорда, 5— нервная трубка, 6—половой бугорок, 7—аллантоис, 8—желточный стебелек, 9— прямая кишка, 10—уроректальная перегородка, 11—мочеполовой синус, 12— мочевой пузырь
Немалое значение принадлежит процессам гибели клеток при образовании полостей тела или сосудов (так называемая кавитация), имеющих вначале вид тяжей без просвета.
В центральной нервной системе сначала образуется больше нервных клеток, чем затем сохраняется, так как часть нейронов, не установивших связи со своими мишенями, погибает.
Наиболее изучены процессы гибели клеток при образовании дефинитивной формы конечностей птиц и млекопитающих. У цыпленка почки конечностей закладываются в виде боковых утолщений соматоплевры примерно на 55-м часу развития. Они вырастают из тела зародыша в виде выступов, покрытых эктодермой
изаполненных мезодермальной тканью. По мере их роста начинают проступать контуры конечностей. Процесс формирования контуров сопровождается отмиранием клеток в ряде мезодермальных участков почки конечности. Участки будущей гибели клеток выявлены с помощью прижизненных красителей.
Так называемая задняя некротическая зона (ЗНЗ) обеспечивает формирование контуров проксимальных областей конечностей. Максимальной протяженности ЗНЗ достигает к 96-му часу развития. К этому времени гибнет 1500—2000 клеток, поглощаемых примерно 150 макрофагами. Другие клетки мезодермы, примыкающие к зоне некроза, не погибают. Группа клеток будущей ЗНЗ, пересаженная со своего местонахождения на боковую поверхность тела зародыша за 40 ч до начала ожидаемого некроза, все равно погибает в те же сроки, что и на своем обычном месте. В данном примере проявляется генетическая запрограммированность гибели определенных клеток, которую можно образно назвать «внутренними часами смерти».
Если клетки ЗНЗ пересадить на дорсальную поверхность почки крыла, то ее клетки не погибнут. Следовательно, некоторые другие клетки способны предотвратить гибель клеток ЗНЗ. Такой способностью обладают клетки мезодермы из дорсальной ненекротической области крыла. Показано, что контакт клеток ЗНЗ с «защищающими» их от гибели клетками дорсальной мезодермы крыла должен продолжаться не менее 6 сут., после чего они могут уже продолжать существование
ибез контакта с этими клетками. Таким образом, гибель клеток контролируется не только генетически, но и на уровне клеточных взаимодействий. Очевидно, что в описанных событиях мы наблюдаем апоптоз, несмотря на сохранение в названиях гибнущих зонах — «некротические».
Интересно сравнение влияния условий на процессы гибели клеток в дистальной части лапок у цыпленка и утенка. Как известно, у утят гибель клеток в дистальной части лапки невелика, вследствие чего у них между вторым, третьим и четвертым пальцами образуются перепонки. Когда создавали химеры, соединяя
363
medwedi.ru

мезодерму из почки ноги утки с эктодермой цыпленка и наоборот, подсаживая затем химерные почки конечностей на боковую поверхность куриного зародыша, в обоих случаях получали развитие по типу утиной конечности, т.е. с перепонкой между пальцами (рис. 8.5).
364
Рис. 8.5. Развитие задней конечности цыпленка (А) и утенка (Б); мутация talpid (В) и опыты по пересадке мезодермы и эктодермы в почках конечностей {Г):
1—мезодерма почки конечности, 2—апикальный эктодермальный гребень, 3— участки некроза между пальцами у зародыша цыпленка, 4—химерные почки конечностей, 5—мезодерма утки, 6—эктодерма цыпленка, 7—мезодерма цыпленка, 8—эктодерма утки
Таким образом, мезодерма цыпленка, обладая способностью к гибели при нормальном развитии, в присутствии эктодермы утки не погибает, т.е. изменяет свои свойства. Приведенный опыт подтверждает значение межклеточных регуляторных механизмов в развитии.
Генетический контроль клеточной гибели прослеживается также на примере мутаций. Анализ мутации под названием ta (talpid) у цыпленка показывает, что в конечности отсутствуют передняя и задняя некротические зоны, в результате чего не происходит формирования контуров, сужающих конечность, и лапка получается очень широкой. Кроме того, у таких мутантов отсутствует область апоптоза под названием темного пятна, в результате чего не происходит разделения большой и малой берцовых костей. Неразделение костей голени и отсутствие их сужения сочетаются с костной полидактилией (многопалостью) и синдактилией (сращением) мягких тканей в ноге. Конечность имеет широкий и веслообразный вид с 6—7 пальцами. Это аутосомно-рецессивная мутация. Аналогичные пороки описаны у человека.
Существуют также мутации, расширяющие зоны апоптоза. У дрозофилы мутация Bar сильно уменьшает глаза, а мутация vestigial уменьшает крылья за счет увеличения участков гибели клеток. Описаны подобные мутации и у кур.
Новейшим примером генетического контроля апоптоза является открытие гена р53. Белок, контролируемый этим геном, обладает способностью при определенных условиях блокировать клеточное деление и запускать механизм апоптоза. Мутационные изменения и дефектность функции этого гена или регулирующих его активность нуклеотидных последовательностей ДИК встречаются в опухолевых клетках и обнаружены, по данным разных исследователей, у 55—70% раковых больных.
Таким образом очевидно, что избирательная гибель клеток, не менее важна для морфогенеза, чем другие клеточные процессы. 'Различные тонкие по замыслу опыты позволяют установить, что гибель клеток имеет три уровня регуляции: генетический контроль, межклеточные взаимодействия и организменный уровень.
8.2.5. Дифференцировка клеток
Дифференцировка — это процесс, в результате которого клетка становится специализированной, т.е. приобретает химические, морфологические и функциональные особенности. В самом узком смысле это изменения, происходящие в клетке на протяжении одного, нередко терминального, клеточного цикла, когда
365
medwedi.ru
начинается синтез главных, специфических для данного клеточного типа, функциональных белков. Примером может служить Дифференцировка клеток эпидермиса кожи человека, при которой в клетках, перемещающихся из базального в шиповатый и затем последовательно в другие, более поверхностные слои, происходит накопление кератогиалина, превращающегося в клетках блестящего слоя в элеидин, а затем в роговом слое — в кератин. При этом изменяются форма клеток, строение клеточных мембран и набор органоидов. На самом деле дифференцируется не одна клетка, а группа сходных клеток. Примеров можно привести множество, так как в организме человека насчитывают порядка 220 различных типов клеток. Фибробласты синтезируют коллаген, миобласты — миозин, клетки эпителия пищеварительного тракта — пепсин и трипсин. 338
В более широком смысле под дифференцировкой понимают постепенное (на протяжении нескольких клеточных циклов) возникновение все больших различий и направлений специализации между клетками, происшедшими из более или менее однородных клеток одного исходного зачатка. Этот процесс непременно сопровождают морфогенетические преобразования, т.е. возникновение и дальнейшее развитие зачатков определенных органов в дефинитивные органы. Первые химические и морфогенетические различия между клетками, обусловливаемые самим ходом эмбриогенеза, обнаруживаются в период гаструляции.
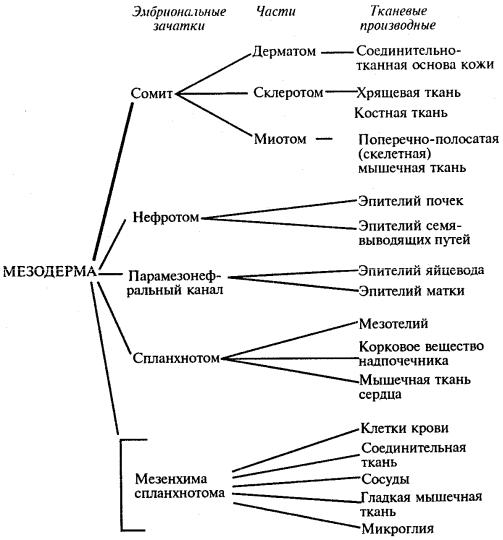
Зародышевые листки и их производные являются примером ранней дифференцировки, приводящей к ограничению потенций клеток зародыша. На схеме 8.1 представлен пример дифференцировки мезодермы (по В. В. Яглову, в упрощенном виде).
Схема 8.1. Дифференцировка мезодермы
366
Можно выделить целый ряд признаков, которые характеризуют степень дифференцированности клеток. Так, для недифференцированного состояния характерны относительно крупное ядро и высокое ядерно-цитоплазматическое
отношение Vядра/Vцитоплазмы (V—объем), диспергированный хроматин и хорошо выраженное ядрышко, многочисленные рибосомы и интенсивный синтез РНК,
высокая митотическая активность и неспецифический метаболизм. Все эти признаки изменяются в процессе дифференцировки, характеризуя приобретение клеткой специализации.
Процесс, в результате которого отдельные ткани в ходе дифференцировки приобретают характерный для них вид, называют гистогенезом. Дифференцировка клеток, гистогенез и органогенез совершаются в совокупности, причем в определенных участках зародыша и в определенное время. Это очень важно, потому что указывает на координированность и интегрированность эмбрионального развития.
В то же время удивительно, что, в сущности, с момента одноклеточной стадии (зиготы) развитие из нее организма определенного вида уже жестко предопределено. Всем известно, что из яйца птицы развивается птица, а из яйца лягушки —лягушка. Правда, фенотипы организмов всегда различаются и могут
367
medwedi.ru
быть нарушены до степени гибели или возникновения порока развития, а нередко могут быть даже как бы искусственно сконструированы, например у химерных животных.
Требуется понять, каким образом клетки, обладающие чаще всего одинаковыми кариотипом и генотипом, дифференцируются и участвуют в гисто- и органогенезе в необходимых местах и в определенные сроки соответственно целостному «образу» данного вида организмов. Осторожность при выдвижении положения о том, что наследственный материал всех соматических клеток абсолютно идентичен, отражает объективную реальность и историческую неоднозначность в трактовке причин клеточной дифференцировки.
В.Вейсман выдвинул гипотезу о том, что только линия половых клеток несет
всебе и передает потомкам всю информацию своего генома, а соматические клетки могут отличаться от зиготы и друг от друга количеством наследственного материала и поэтому дифференцироваться в разных направлениях. Ниже приведены факты, подтверждающие возможность изменения наследственного материала в соматических клетках, но их надо трактовать как исключения из правил.
Вейсман опирался на данные о том, что в ходе первых делений дробления яиц лошадиной аскариды происходит отбрасывание (элиминация) части хромосом в соматических клетках эмбриона. В дальнейшем было показано, что отбрасываемая ДНК содержит главным образом часто повторяющиеся последовательности, т.е. фактически не несущие информации.
Развитие представлений о механизмах цитодифференцировки изображено на схеме 8.2.
368

Позже были обнаружены и другие примеры изменения количества наследственного материала в соматических клетках как на геномном, так и на хромосомном и генном уровнях. Описаны случаи элиминации целых хромосом у циклопа, комара и у одного из представителей сумчатых. У последних из соматических клеток самки элиминируется Х-хромосома, а из клеток самца — Y- хромосома. В результате соматические клетки у них содержат только по одной Х- хромосоме, а в линии половых клеток сохраняются нормальные кариотипы: XX или XY.
В политенных хромосомах слюнных желез двукрылых ДНК может синтезироваться несинхронно, например при политенизации гетерохроматиновые участки реплицируются меньшее число раз, чем эухроматиновые. Сам процесс политенизации, напротив, приводит к значительному увеличению количества ДНК в дифференцированных клетках по сравнению с родоначальными клетками.
Такой механизм репликации ДНК, как амплификация, также приводит к многократному увеличению количества некоторых генов в одних клетках по сравнению с другими. В овогенезе многократно увеличивается число
369
medwedi.ru
рибосомальных генов, могут амплифицироваться и некоторые другие гены. Имеются данные о том, что в некоторых клетках в процессе дифференцировки происходит перестройка генов, например иммуноглобулиновых генов в лимфоцитах.
Однако в настоящее время общепризнанной является точка зрения, ведущая начало от Т. Моргана, который, опираясь на хромосомную теорию наследственности, предположил, что дифференцировка клеток в процессе онтогенеза является результатом последовательных реципрокных (взаимных) влияний цитоплазмы и меняющихся продуктов активности ядерных генов. Таким образом, впервые прозвучала идея о дифферециальной экспрессии генов как основном механизме цитодифференцировки. В настоящее время собрано много доказательств того, что в большинстве случаев соматические клетки организмов несут полный диплоидный набор хромосом, а генетические потенции ядер соматических клеток могут сохраняться, т.е. гены не утрачивают потенциальной функциональной активности.
Сохранение полного хромосомного набора развивающегося организма обеспечивается прежде всего механизмом митоза (возможные случаи соматических мутаций, возникающих, как исключение, во внимание не принимаем). Проведенные цитогенетическим методом исследования кариотипов различных соматических клеток показали почти полную их идентичность. Цитофотометрическим способом установлено, что количество ДНК в них не уменьшается, а методом молекулярной гибридизации показано, что клетки разных тканей идентичны по нуклеотидным последовательностям. На этом основании цитогенетический метод применяют для диагностики хромосомных и геномных болезней человека (хотя ошибки методов достигают 5— 10%), а метод гибридизации ДНК —для идентификации личности и установления степени родства.
Помимо установленной количественной полноценности ДНК большинства соматических клеток большой интерес представляет вопрос о сохранении функциональных свойств содержащегося в них наследственного материала. Все ли гены сохраняют способность к реализации своей информации? О сохранении генетических потенций ядер можно судить по результатам опытов, проведенных над растениями и животными. Прошедшая длительный путь дифференцировки соматическая клетка моркови способна развиваться в полноценный организм (рис. 8.6). У животных отдельные соматические клетки после стадии бластулы, как правило, не способны развиваться в целый нормальный организм, но их ядра, будучи пересажены в цитоплазму овоцита или яйцеклетки, начинают вести себя соответственно той цитоплазме, в которой они оказались.
Опыты по пересадке ядер соматических клеток в яйцеклетку впервые были успешно осуществлены в 50-х гг. в США, а в 60—70-х гг. получили широкую известность опыты английского ученого Дж. Гердона. Используя африканскую шпорцевую лягушку Xenopus laevis, он в небольшом проценте случаев получил развитие взрослой лягушки из энуклеированной яйцеклетки, в которую пересаживал ядро из эпителиальной клетки кожи лягушки или кишечника головастика, т.е. из
370