

Биология Ярыгин 2003
.pdfфакторов среды. Примером служит интенсивность красной окраски цветков ночной красавицы, убывающая в ряду генотипов АА, Аа, аа, или интенсивность пигментации кожи у человека, увеличивающаяся при возрастании числа доминантных аллелей в системе полигенов от 0 до 8 (см. рис. 3.80). Влияние средовых факторов на экспрессивность признака демонстрируется усилением степени пигментации кожи у человека при ультрафиолетовом облучении, когда появляется загар, или увеличением густоты шерсти у некоторых животных в зависимости от изменения температурного режима в разные сезоны года.
6.1.2.Роль наследственных и средовых факторов
вопределении половой принадлежности организма
Соотносительная роль наследственной программы и факторов среды в формировании фенотипа особи может быть прослежена на примере развития признаков половой принадлежности организма.
Пол организма представляет собой важную фенотипическую характеристику, которая проявляется в совокупности свойств, обеспечивающих воспроизведение потомства и передачу ему наследственной информации. В зависимости от значимости этих свойств различают первичные и вторичные половые признаки.
Под первичными половыми признаками понимают морфофизиологические особенности организма, обеспечивающие образование половых клеток — гамет, сближение и соединение их в процессе оплодотворения. Это наружные и внутренние органы размножения. Вторичными половыми признаками называют отличительные особенности того или другого пола, не связанные непосредственно с гаметогенезом, спариванием и оплодотворением, но играющие важную роль в половом размножении (обнаружение, и привлечение партнера). Их развитие контролируется гормонами, синтезируемыми первичными половыми органами.
6.1.2.1. Доказательства генетического определения признаков пола
Важным доказательством в пользу наследственной детерминированности половой принадлежности организмов является наблюдаемое у большинства видов соотношение по полу 1:1 (табл. 6.1).
Таблица 6.1. Соотношение по полу у разных видов
231
medwedi.ru

Такое соотношение может быть обусловлено образованием двух видов гамет представителями одного пола (гетерогаметный пол) и одного вида гамет — особями другого пола (гомогаметный пол). Это соответствует различиям в кариотипах организмов разных полов одного и того же вида, проявляющимся в половых хромосомах. У гомогаметного пола, имеющего одинаковые половые хромосомы XX, все гаметы несут гаплоидный набор аутосом плюс Х-хромосому. У гетерогаметного пола в кариотипе кроме аутосом содержатся две разные или только одна половая хромосома (XY или ХО). Его представители образуют два вида гамет, различающиеся по гетерохромосомам: Х и Y или Х и 0.
У разных видов организмов хромосомный механизм определения пола реализуется по-разному. У человека и других млекопитающих, а также у дрозофилы гомогаметным является женский пол (XX), а гетерогаметным — мужской (XY). У некоторых насекомых (клопы рода Protenor) гетерогаметный мужской пол имеет лишь одну Х-хромосому (ХО). У птиц и некоторых насекомых женский пол является гетерогаметным (XY), а мужской — гомогаметным (XX). У некоторых бабочек гетерогаметный женский пол имеет одну Х-хромосому (ХО). В большинстве выше описанных случаев пол вновь образующегося организма определяется сочетанием половых хромосом, возникающим в зиготе при оплодотворении (табл. 6.2).
Таблица 6.2. Хромосомный механизм определения полов
Таким образом, хромосомный механизм определения половой принадлежности организмов обеспечивает равновероятность встречаемости представителей обоих полов. Это имеет большой биологический смысл, так как обусловливает максимальную вероятность встречи самки и самца, потомки получают более разнообразную наследственную информацию, поддерживается
232
оптимальная численность особей в популяции.
Исследования, проведенные на дрозофиле, обнаружили незначительную роль Y-хромосомы в определении пола. Особи с кариотипом ХО внешне являются типичными самцами, но стерильными, а особи с кариотипом XXY — плодовитые самки. Y-хромосома у дрозофилы по размеру близка к Х-хромосоме, однако она генетически инертна, так как состоит в основном из гетерохроматина. В связи с этим первичное определение пола у дрозофилы обусловлено не столько сочетанием в зиготе Х- и Y-хромосом, сколько соотношением числа Х-хромосом и наборов аутосом (половой индекс). У нормальных самок половой индекс равен 1 (2Х:2А), у нормальных самцов — 0,5 (XY:2A). Увеличение полового индекса более 1 при увеличении дозы Х-хромосомы (ЗХ:2А) приводит к развитию сверхсамок, а величина этого показателя ниже 0,5 (XY:3A) обеспечивает появление самцов. При значениях полового индекса более 0,5, но менее 1 (2Х:ЗА) развиваются интерсексы.
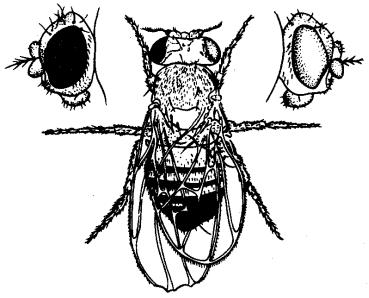
О значении дозы Х-хромосом в определении пола у дрозофилы свидетельствует развитие латерального гинандроморфизма у мух при утрате Х- хромосомы одной из клеток на стадии первого деления зиготы. В результате половина тела, развивающаяся из клетки с нормальным кариотипом (2АХХ), несет признаки самки, а другая половина, клетки которой лишены одной Х-хромосомы (2АХО), имеет признаки самца (рис. 6.2).
Рис. 6.2. Латеральный гинадроморфизм у Drosophila melanogaster, гетерозиготной по гену окраски глаз, находящемуся в Х-хромосоме
Левая половина мухи, клетки которой несут 2Х, имеет признаки самки и красный глаз; правая половина мухи, клетки которой несут одну Х-хромосому, имеет признаки самца и белый глаз
Обнаружение зависимости половой принадлежности развивающегося
233
medwedi.ru

организма от дозы Х-хромосом у дрозофилы и некоторых других насекомых привело американского генетика С. Бриджеса (1922) к формулировке гипотезы генного баланса, в соответствии с которой организм изначально бисексуален, т.е. несет в себе задатки и мужского, и женского полов. Развитие признаков одного из них в ходе онтогенеза определяется балансом женских и мужских генов — детерминаторов пола. У дрозофилы эти гены сосредоточены не только в половой Х- хромосоме, но и в аутосомах. Поэтому пол организма у них зависит от соотношения этих хромосом. У плодовой мухи Y-хромосома, будучи генетически инертной, в определении признаков пола не участвует.
У человека Y-хромосома играет важную роль в детерминации пола. Она содержит определенное количество генов, часть из которых гомологична генам Х- хромосомы, а часть не имеет в ней гомологов и наследуется только по мужской линии. Некоторые из этих генов непосредственно связаны с детерминацией мужского пола. Поэтому у человека присутствие Y-хромосомы в кариотипе независимо от количества Х-хромосом (2AXXY, 2AXXXY) обеспечивает развитие мужского пола. Особи с кариотипом 2АХО являются женщинами, несмотря на уменьшение дозы Х-хромосомы. Однако значение баланса генов в определении половой принадлежности организма подтверждается тем, что особи с кариотипами 2АХО, 2АХХХ, 2AXXY, 2AXXXY и т.д. отличаются наличием пороков развития и часто стерильны.
Как было отмечено, у большинства видов животных пол организма определяется сочетанием или количеством половых хромосом в диплоидном наборе аутосом. Однако у некоторых насекомых (пчелы, муравьи, осы) представители разных полов отличаются числом наборов хромосом. Самки у них развиваются из оплодотворенных яиц и поэтому диплоидны, самцы — из неоплодотворенных яиц и первично гаплоидны. В такой ситуации соотношение по полу зависит от того, происходит оплодотворение или нет. Половой состав популяции может быть различным в разные периоды ее жизни.
Зависимость половой принадлежности организмов от сочетания или количества половых хромосом в его кариотипе свидетельствует о том, что гены, определяющие пол, располагаются главным образом в этих хромосомах. Однако известен целый ряд примеров, указывающих на то, что в развитии признаков пола принимают участие также гены, локализованные в аутосомах. В частности, в геноме дрозофилы присутствует ген, называемый трансформатором. Рецессивный аллель этого гена в гомозиготном состоянии tt превращает женские зиготы 2Х в фенотипических самцов, которые оказываются стерильными. Самки1 XXt+t+ или XXt+t плодовиты, особи с кариотипом XYtt — нормальные самцы, организмы с кариотипом XXtt — превращенные стерильные самцы. Другим примером влияния аутосомных генов на развитие признаков пола может служить существование у кукурузы генов, мутантные аллели которых превращают ее из однодомного растения в двудомное, препятствуя развитию органов либо мужского, либо
1 t+ — доминантный аллель гена.
234
женского пола. Рецессивный аллель гена sk(silkless) в гомозиготном состоянии вызывает недоразвитие семяпочек и превращает растение в мужское. Мутантный аллель другого гена ts (tassel seed) в гомозиготном состоянии вызывает развитие семяпочек и семян в метелке, а пыльники не развиваются.
У человека развитие организма по мужскому типу обеспечивается не только геном, расположенным в Y-хромосоме и определяющим способность к синтезу мужского полового гормона — тестостерона, но и Х-сцепленным геном, контролирующим синтез белка-рецептора этого гормона. Мутация упомянутого Х- сцепленного гена приводит к развитию синдрома тестикулярной феминизации (см. разд. 3.6.5.2).
Приведенные примеры доказывают, что пол организмов как признак развивается на основе полученной от родителей наследственной информации и находится под контролем группы взаимодействующих генов, расположенных как в половых хромосомах, так и в аутосомах.
6.1.2.2.Доказательства роли факторов среды
вразвитии признаков пола
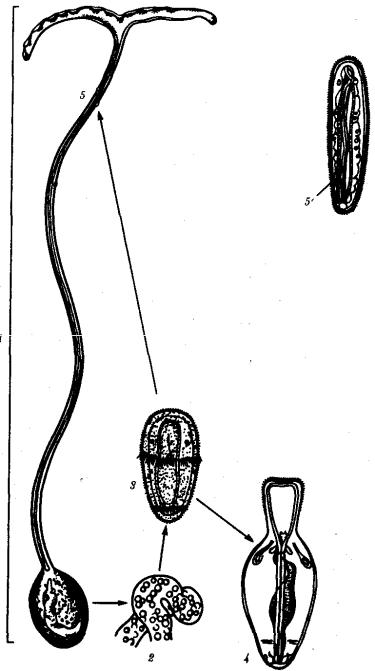
Убольшинства видов развитие признаков пола осуществляется на основе наследственной программы, заключенной в генотипе. Однако известны примеры, когда половая принадлежность организма целиком зависит от условий, в которых он развивается. Так, у морского червя Bonellia viridis пол зависит от того, будет зигота развиваться в непосредственном контакте с материнским организмом или самостоятельно. В первом случае образуется самец, во втором —самка (рис. 6.3).
Увысших организмов значение среды в определении признаков пола, как правило, невелико. Вместе с тем даже для них известны примеры, когда в определенных условиях развития происходило переопределение пола на
противоположный, несмотря на имеющуюся комбинацию хромосом в зиготе. Возможность переопределения пола обусловлена тем, что первичные закладки гонад у эмбрионов всех животных изначально бисексуальны. В процессе онтогенеза происходит выбор направления развития закладки в сторону признаков одного пола, включая дифференцировку половых желез, формирование половых путей и вторичных половых признаков. Первостепенная роль в развитии мужского или женского фенотипа принадлежит гормонам, образуемым гонадами.
Ведущим началом в дифференцировке пола являются гены, контролирующие уровень мужских и женских половых гормонов. Соотношение этих гормонов в организме является особенно важным для формирования и поддержания соответствующих признаков. Так как у млекопитающих и человека дифференцировка пола начинается очень рано, то полное переопределение пола в эмбриогенезе у них невозможно. Известны, однако, случаи, в частности у крупного рогатого скота, когда в разнополых двойнях рождались бесплодная телочка — интерсекс (фри-мартин) — и бычок. Это происходит потому, что клетки семенников мужского эмбриона начинают синтез соответствующего гормона раньше, чем клетки яичников эмбриона женского пола, оказывая на организм последнего
235
medwedi.ru
маскулинизирующее действие, вплоть до формирования половых органов мужского типа.
Рис. 6.3. Роль условий среды в развитии признаков пола у морского червя Bonellia viridis:
1 — самка, 2 — яйца, 3 — личинка, 4 — молодая самка, 5 — самец, 5' — самец в увеличенном виде
У некоторых видов в ходе обычного онтогенеза при определенных условиях происходит естественное переопределение пола. Так, в Тихом океане обитают рыбки Labroides dimidiatus, живущие стайками из самок и одного самца. Каждый из
236
членов такой семьи, кроме самца, постоянно находится в состоянии стресса, источником которого является самец. При этом уровень напряженности различается от самки к самке, так что можно выделить α-, β-, γ- самок и т.д. Гибель самца вызывает превращение α-самки (главной самки, сбрасывающей напряжение) в самца с полноценными семенниками. Описанное переопределение зависит от уровня в организме гормонов, выделяемых клетками надпочечников.
У человека угнетение эндокринной функции яичников, наступающее после прекращения гаметогенеза (в определенном возрасте у женщин), приводит к появлению некоторых вторичных признаков мужского пола, например росту волос на лице.
Искусственное изменение условий развития организмов в эксперименте приводит у некоторых видов к полному переопределению пола, вплоть до появления способности образовывать гаметы противоположного пола (некоторые виды амфибий, рыб, птиц). Например, кормление мальков самцов аквариумных рыбок до 8 мес. пищей с добавкой эстрогенов превращало их в самок, способных к скрещиванию с натуральными самцами и образованию полноценного потомства. В экспериментах на курах удаление половых желез до окончания периода полового созревания или подсадка гонад противоположного пола вели к появлению вторичных признаков другого пола: у курицы развивались петушиное оперение и инстинкты петуха, а петух приобретал куриное оперение. Исходная генетическая конституция, в том числе и по половым хромосомам, при этом оставалась прежней.
Из примеров, рассмотренных выше, следует, что генотип особи заключает в себе информацию о возможности формирования признаков того или иного пола, которая реализуется лишь при определенных условиях индивидуального развития. Изменение этих условий может стать причиной переопределения признаков пола. При этом у организмов разных видов относительное значение генетических и средовых факторов не одинаково: у одних видов определяющим фактором является среда, у других — наследственная программа.
6.2. РЕАЛИЗАЦИЯ НАСЛЕДСТВЕННОЙ ИНФОРМАЦИИ В ИНДИВИДУАЛЬНОМ РАЗВИТИИ. МУЛЬТИГЕННЫЕ СЕМЕЙСТВА
В процесса индивидуального развития организм закономерно меняет свои характеристики. Особенно интенсивные изменения происходят в эмбриональном периоде онтогенеза, когда из зиготы формируются структуры многоклеточного организма. При этом все многообразие клеток, выполняющих в организме различные функции, происходит из одной клетки путем митотического деления. Так как в результате митоза дочерние клетки получают полноценную наследственную информацию, заключенную в кариотипе, все клетки организма в генотипическом отношении равноценны. Некоторые различия, однако, наблюдаются за счет цитоплазматических генов, например митохондриальных, которые распределяются при делении не строго равномерно.
237
medwedi.ru
Чем определяются морфологические, физиологические и биохимические различия, появляющиеся между клетками в ходе развития? В процессе овогенеза в цитоплазме яйцеклетки накапливаются не только богатые энергией вещества, обеспечивающие развитие зародыша, но и мРНК для синтеза белков, необходимых на самых ранних стадиях эмбрионального развития. Распределение этих веществ в цитоплазме яйцеклетки оказывается неравномерным. Проникновение сперматозоида
вяйцеклетку вызывает перераспределение отдельных компонентов в объеме клетки,
всвязи с чем уже при первых делениях зиготы в дочерних клетках оказывается цитоплазма с разным составом веществ.
Взаимодействие между компонентами цитоплазмы и ядром приводит к дерепрессии определенных генов. Их продукты определяют дальнейшее углубление различий между разными частями зародыша, т.е. дифференцировку. Возникающие различия порождают новые взаимодействия между соседними клеточными группами, которые вызывают дерепрессию новых генов, вследствие чего меняется спектр активных генов и, следовательно, генетическая программа на последующий отрезок процесса развития. Таким образом, в ходе индивидуального развития первоначально репрессированный геном зиготы подвергается постепенной дерепрессии, причем в разных частях зародыша дерепрессируются разные группы генов. Набор активно функционирующих генов определяет своеобразие спектра белков, которые синтезируются клетками, выполняющими различные функции.
Как отмечалось выше, в процессе онтогенеза в клетках организма происходит смена активно функционирующих генов. Гены, транскрибировавшиеся в эмбриональном периоде, к моменту рождения или непосредственно после него репрессируются, в то же время активируются гены, определяющие специфические функции клеток во взрослом организме. Нередко вещества, продуцируемые определенным типом клеток в разные периоды онтогенеза, несколько различаются по своим свойствам. Изменение свойств диктуется изменением условий существования организма, например в эмбриональном и постэмбриональном периодах развития. Эти различия объясняются сменой функционирования близких, но не идентичных по заключенной в них информации генов. Такие гены в ряде случаев образуют группы, получившие название мультигенных семейств. Примером тому служат гены гемоглобина.
Мультигенное семейство — это группа генов, очень близких по
нуклеотидным последовательностям, со сходными фенотипическими функциями. Число генов в разных семействах у представителей разных видов варьирует от единиц до нескольких сотен. К примеру, число генов гистонов у разных видов в отдельных семействах колеблется от 10 до 1200, генов тРНК — от 6 до 400, генов 5SPHK — от 200 до 24000, генов α-глобинов — от 1 до 5, (β-глобинов — от 2 до 7. К числу белков, кодируемых мультигенными семействами, кроме указанных выше, относятся актины и тубулины, играющие важную роль в подвижности клеток, коллагены соединительной ткани, некоторые белки клеточных мембран и сыворотки крови.
Мультигенные семейства могут быть организованы в геноме по-разному (рис.
238

6.4). Так, члены семейства идентичных генов РНК у человека располагаются в виде тандемных повторов, в которых структурные гены разделены некодирующими — спейсерными — участками. Однако гены семейства не всегда идентичны. Например, в семействах глобиновых генов тандемно сцеплены близкие, но не одинаковые гены. В случае гистоновых генов у некоторых видов тандемно повторяются целые пакеты (кластеры) неидентичных генов, определяющих синтез разных видов гистонов, что может быть описано формулой Н2А — НЗ —Н2В — Н4 — H1 —. Возможно также диффузное распределение генов семейства по нескольким хромосомам, что показано для генов актинов и тубулинов.
Рис. 6.4. Типы организации мультигенных семейств:
I — идентичные гены, сцепленные тандемно (гены рРНК); II — близкие, но не идентичные гены, сцепленные тандемно (гены глобинов); III — скопления неидентичных генов, сцепленных тандемно (гистоновые гены); IV — близкие гены, рассеянные по нескольким хромосомам (гены актинов или тубулинов)
Описанные варианты организации мультигенных семейств создают необходимые условия для эффективной регуляции экспрессии соответствующих генов. Так, если продукт определенного гена необходим лишь на небольшом отрезке времени в онтогенезе, но в значительных количествах, мультигенное семейство образовано большим числом идентичных генных копий, обычно соединенных тандемно. Примером могут служить гены рРНК, которые в геноме соматических клеток взрослой шпорцевой лягушки представлены 450 копиями. Вместе с тем в овогенезе для быстрого образования необходимого количества рибосом, которых в яйце Xenopus содержится около 1012, гены рРНК амплифицируются и число их копий возрастает в 4000 раз. В других мультигенных семействах, состоящих, как правило, из неидентичных генов, в ходе онтогенеза происходит переключение с
239
medwedi.ru

одного гена на другой. Белки, контролируемые определенными генами такого семейства, наилучшим образом соответствуют либо условиям на разных стадиях онтогенеза, либо клеткам различных типов.
Рис. 6.5. Распределение генов α- и β-глобинов человека
Глобиновые гены изображены в виде прямоугольников; вертикальные полосы внутри соответствуют экзонам, остальные участки — интронам
Наиболее изучены в этом отношении мультигенные семейства α- и β- глобиновых генов (рис. 6.5). У человека они представлены кластером из 7 β- глобиновых генов, расположенных в 11-й хромосоме, и кластером из 5 α- глобиновых генов, локализующихся в 16-й хромосоме. У эмбрионов человека активно функционируют ζ (дзета)-глобиновый ген из α- семейства и ε (эпсилон)- глобиновый ген из β-семейства, обеспечивающие образование эмбрионального ζ2ε2- гемоглобина.
На более поздних стадиях онтогенеза у плода эти гены репрессируются, но дерепрессируются другие гены семейств, определяющие синтез фетального α2γ2- гемоглобина. После рождения начинают экспрессироваться δ (дельта)- β и (бета)- глобиновые гены, обеспечивающие образование преобладающего α2β2- и минорного α2β2- видов гемоглобина взрослого человека. В обоих кластерах имеются также неэкспрессирующиеся псевдогены ψα1, ψξ1, ψβ1, ψβ2.
Переключение генов в мультигенных семействах происходит не только в соответствии со стадией индивидуального развития, но и с типом и местом локализации клеток в организме. Так, эмбриональный гемоглобин С,г б2 образуется имеющими ядра мегалобластами в стенке желточного мешка. На 6-й неделе развития происходит смена экспрес-сируемых генов и переход к синтезу гемоглобина безъядерными эритроцитами печени и селезенки плода. Позднее главным местом образования гемоглобина становится костный мозг, где вскоре после рождения начинается синтез взрослых гемоглобинов α2β2 и α2δ2. Смена типов синтезируемого гемоглобина у эмбриона, плода и после рождения связана с конкретными условиями существования организма на разных стадиях онтогенеза. Так, у человека гемоглобин плода имеет более высокое сродство к кислороду, чем гемоглобин взрослого, что облегчает перенос кислорода через плаценту.
Таким образом, изменение характеристик фенотипа организма на разных стадиях онтогенеза является результатом регуляции экспрессии генов, которая имеет целью в одних случаях наращивание продукции определенных белков, а в других — переход от синтеза одного белка к синтезу белка более соответствующего изменяющимся условиям существования.
240