

Мякушко С.А. Порівняльна анатомія. 2019
.pdfчеревних лусок змії прикріплюється потужний підшкірний м’яз, це полегшує рух тварини через чіпляння за ґрунт.
У ссавців підшкірна мускулатура є комбінованою. Значну її частину в передньому кінці тіла утворює підшкірний м’яз тулуба, який походить від грудного м’яза, що обслуговує кінцівки. Одночасно стискач шиї, утворюючи кільце поверхневих волокон навколо шиї, є елементом вісцеральної мускулатури. Цей підшкірний шар, даючи пучки від шиї на череп і щоки, разом з іншими бере участь у формуванні мускулатури обличчя.
Вісцеральна мускулатура
Вісцеральний скелет, як і мускулатура, що його обслуговує, є похідними передньої частини первинної кишки та розвивається не із міотомів, а із мезенхіми вісцерального листка бічної пластинки.
У сучасних хребетних вона представлена як гладенькими м’язовими волокнами (стінки кишечника, судин, сечовивідних шляхів та ін.), так і поперечносмугастими (мускулатура бранхіомерів або зябрового апарату нижчих хребетних, мускулатура під’язикового апарату, голови, гортані і частково деяких внутрішніх органів).
Особливістю мускулатури зябрової області є її поперечна посмугованість. Оскільки вся система вісцеральної мускулатури пов’язана з первинною кишкою, припускають, що у давніх форм хребетних вся мускулатура травного тракту була гладенькою. У сучасних хребетних вона поділена на дві частини: краніальну поперечносмугасту та каудальну, яка залишилася гладенькою. Для захоплення їжі та дихання (первинних функцій рота і глотки) потрібні були більш енергійні і координовані рухи, властиві поперечносмугастим м’язам; для повільних скорочень м’язового волокна, які задовольняють процеси травлення, достатньо гладенької мускулатури. Не виключено, що в процесі такої перебудови м’язів зіграло роль те, що м’язи краніальної частини кишки отримали зв’язок з елементами вісцерального скелета; гладенькі м’язи каудальної частини лежать виключно в м’яких тканинах кишки.
Зяброва мускулатура досить добре розвинута у круглоротих у вигляді м’язів, які стискують зяброві мішки і спеціалізованих м’язів язика.
Найбільш повно мускулатура зябрової області представлена в акул, її можна вважати вихідною для всіх інших щелепноротих.
130
Ускладі зябрового бранхіомеру знаходяться поверхневі констриктори (стискачі) — дорсальний і вентральний, аддуктори, що зменшують кути між члениками дуг, а також міждугові м’язи, що з’єднують фарінгоелементи сусідніх дуг. М’язи щелепної дуги включають три компоненти. По-перше, це її дорсальний констриктор, або підіймач піднебінноквадратного хряща, який фактично бере участь у його підвішуванні. Далі, гіпертрофований аддуктор у вигляді привідного м’яза нижньої щелепи зі складною внутрішньою організацією. Нарешті, це вентральний констриктор, який злитий з констриктором під’язикової дуги та з’єднує половинки нижньої щелепи. Спинний констриктор під’язикової дуги добре розвинений та займає велику поверхню, проте він не диференційований.
Укостистих риб дорсальний констриктор щелепової дуги диференційований для забезпечення рухів комплексу первинної верхньої щелепи, гіомандибуляра та симплектикума. При цьому підіймач піднебінної дуги відводить комплекс убік, а аддуктор дуги — наближує його до середньої лінії, звужуючи тим самим ротоглоткову порожнину. Спинний констриктор під’язикової дуги в зв’язку з появою зябрової кришки диференціювався на два м’язи: відвідний м’яз зябрової кришки та її підіймач. Крім того, наявний аддуктор, який приєднується до кришки зсередини, від черепної коробки.
Убагатьох костистих розвинутий трапецієподібний м’яз.
Убезхвостих амфібій в м’язах щелепової дуги підіймач очного яблука бере участь у вертикальних переміщеннях ока. Передній міжщелеповий м’яз впливає на внутрішні деформації нижньої щелепи у разі затискування ніздрів і викиданні язика. Дорослі жаби не здатні до активного розширення глотки у поперечному напрямку, під’язиковий апарат здійснює лише вертикальні руху.
Задня частина поверхневого констриктора риб у наземних тетрапод посилюється і вже у плазунів розростається у великий м’яз, який вкриває шию з боків і вентральної сторони, він отримав назву стискач шиї. У ссавців він поділяється на глибокий і поверхневий шари. М’язові волокна обох шарів розростаються на ділянку голови та дають початок складній системі підшкірної лицевої мускулатури. Від глибшого констриктора шиї походять м’язи рота, губ, щік.
131
Ряд м’язів зябрового апарату риб, з втратою зябрового дихання у наземних форм редукувався, за винятком дрібних пучків, які збереглися як м’язи під’язикового апарата, глотки та гортані. Останні мають величезне значення у людини, оскільки управляють голосовими зв’язками. З цієї групи м’язів у наземних хребетних найкраще збереглася трапецієподібна мускулатура, яка поширилася на плечовий пояс і зв’язує його з головою, шиєю, частиною грудного відділу хребта. Передні пучки трапецієподібних м’язів можуть відокремлюватися, утворюючи самостійні груднинно-соскоподібний і клю- чично-соскоподібний м’язи. Останній, у разі редукції ключиці, зростається з пучками дельтоподібного м’язу, утворюючи тонкий і довгий м’яз від голови до грудної кінцівки.
Гладенька мускулатура. Основна частина такої мускулатури розвивається за рахунок мезодерми. Лише деякі з них розвиваються з інших джерел. Так гладенькі м’язові волокна шкіри амфібій розвиваються за рахунок основного шару ектодерми; ектодермальне походження мають і гладенькі м’язи, які зовні вкривають потові залози ссавців.
Гладенька мускулатура шкіри розвивається з елементів шкірного листка соміта — дерматома. Гладенька мускулатура кишки — із вісцерального листка бічних пластинок соміта; м’язи сечостатевих органів і стінок судин — за рахунок мезенхіми бічних пластинок, а стінки судин шкіри — із мезенхіми самої шкіри.
Гладенька мускулатура шкіри найчастіше не організована в окремі м’язи, але до піхви пер птахів і волосяних сумок ссавців ідуть окремі м’язові пучки. Гладенька мускулатура стінки судин утворює суцільний кільцевий шар, на капілярах гладенькі волокна ідуть поздовжньо. В стінці кишки два шари гладеньких м’язових волокон: внутрішній кільцевий і зовнішній поздовжній; їх сумісна дія спричинює перистальтику (може бути й третій середній шар гладеньких волокон, які ідуть навскіс). Стінка сечовивідних шляхів також має кільцевий і поздовжній шари із гладеньких м’язових волокон.
Зараз існують два погляди на співвідношення між гладенькою і поперечносмугастою мускулатурою хребетних. Одні вчені (І.І. Шмальгаузен та ін.) вважають, що між цими двома типами мускулатури існують досить глибокі відмінності, про що свідчать різні походження та джерела іннервації. Інші (А. Ромер, Т. Парсонс) вважають,
132
що між цими типами мускулатури глибокі відмінності відсутні, а поперечносмугасті м’язи утворилися з гладеньких. Як докази наводять приклади трансформації гладеньких м’язів у поперечносмугасті. Ця думка мала би більше підґрунтя, якби були наявними приклади і зворотних перетворень. Цілком справедливо, що різні джерела походження гладенької (із мезенхіми вісцерального листка бічної пластинки) і поперечносмугастої (із міотомів) мускулатури, вважають важливими аргументами, але сам матеріал міотомів і мезенхіми вісцерального листка є похідними середнього зародкового листка — мезодерми. Ймовірно, глибинні процеси диференціації мезодерми є складнішими, ніж сучасні уявлення про них.
ЗАПИТАННЯ ДЛЯ САМОКОНТРОЛЮ
1.Назвіть, які функції виконує опорно-рухова система?
2.Які існують різновиди хрящової тканини, в яких ділянках тіла вини представлені?
3.У чому полягають відмінності у розвитку хондральних і шкірних кісток?
4.Охарактеризуйте основні складові елементи осьового скелета.
5.Охарактеризуйте сутність основних теорій щодо походження хребців.
6.Назвіть характерні риси будови хребців різних типів.
7.Які відділи виділяють у хребті представників різних груп тварин?
8.Порівняйте будову ребер і груднини серед представників різних груп хребетних.
9.Охарактеризуйте головні етапи розвитку мозкового черепа.
10.Порівняйте будову нейрокраніуму первинноводних хребетних.
11.Наведіть відомі Вам класифікації типів черепів амніот.
12.У чому полягають відмінності у модифікаціях діапсидного черепа рептилій і птахів?
13.Назвіть специфічні риси будови черепа ссавців.
14.Чим відрізняється походження елементів вісцерального черепа від мозкового?
133
15.Які основні еволюційні напрямки перетворення зябрових дуг?
16.Які існують типи приєднання щелепної дуги до мозкового черепа?
17.Охарактеризуйте основні типи кінетизму черепа серед птахів.
18.Охарактеризуйте основні тренди еволюційних перетворень ротового апарату тетрапод.
19.Назвіть основні риси будови різних типів хвостових плавців риб.
20.Які основні етапи розвитку парних кінцівок хребетних?
21.Порівняйте основні теорії походження парних плавців.
22.У чому полягає гомодинамність будови парних кінцівок тет-
рапод?
23.Назвіть специфіку будови интеркарпального та інтертарзального суглобів наземних хребетних.
24.Які риси будови пост краніального скелета птахів пов’язані
зпольотом?
25.Порівняйте будову плечового поясу тетрапод.
26.Порівняйте будову тазового поясу тетрапод.
27.У чому полягає специфіка будови різних типів м’язової тка-
нини?
28.Порівняйте будову осьової мускулатури анамній.
134
РОЗДІЛ 3 ТРАВНА СИСТЕМА
Будова органів травної системи у сучасних хребетних має широкий спектр відмінностей, які обумовлені, з одного боку, рівнем організації та систематичним положенням різних груп хребетних, з
іншого — трофічними спеціалізаціями в межах кожної із груп.
Розвиток органів травлення. В ембріогенезі хребетних
першою із органів травної системи утворюється первинна кишкова трубка — архентерон. Вона з’являється на стадії гаструли, порож-
нина якої називається гастроцелем. В цей же період утворюється й один із отворів кишкової трубки — гастропор, але він скоро закривається. Таким чином, в розвитку ембріональної кишки є такий період, коли вона має форму сліпозамкненої трубки. Після стадії гаструли розвиток кишки полягає, головним чином, в її видовженні, диференціації відділів, утворенні похідних органів, формуванні складної гістологічної будови її стінки (рис. 3.1).
В амфібій, яйця яких є мезолецитальними (містять середню кількість жовтка), розвиток травної системи відбувається за найпростішою схемою: на місці бластопора утворюється клоака. Перед проривом
вторинного рота на передньому кінці зародка утворюється вгинання — стомодеум. Після прориву стомодеум формує передню, екто-
дермальну частину кишкової трубки. У тварин з полілецитальними яйцями та у ссавців порожнина первинної кишки спочатку невіддільна від жовткового мішка, пізніше поступово відшнуровується від нього. Бластопор рано закривається, а клоака або анальний отвір
проривається потім заново — аналогічно вторинному роту, з утворенням ектодермального вгинання — проктодеума, який потім фор-
мує задню частину клоаки (або прямої кишки у більшості ссавців). Поступово формуються дивертикули (сліпі вирости) первинного кишечнику — закладки щитоподібної залози (на місці ендостиля), легенів, печінки, підшлункової залози та алантоїса. Вхід у жовтковий мішок пізніше заростає, хоча може залишатися його рудимент. Першим відділом, який диференціюється в головному кінці первинної кишки, є глотка. Глотковий відділ кишки дає початок досить важливим органам, які пізніше увійшли до респіраторної системи.
135
Порівняно на ранніх стадіях розвитку первинної кишки утворюється печінковий виріст, який відходить від її вентральної стінки. Невдовзі після печінкового, на дорсальній стінці кишки, утворюється основний виступ підшлункової залози.
3
4
15
|
|
2 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
13 |
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
12 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
14 |
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
11 |
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
10 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
15 |
||||
|
|
9 |
|
|
|
|
|
|
|
|
|
||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
20 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
21 |
|
|
|
|
|
|
|
|
|
||
8 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
19 |
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
16 |
|||||||||
|
|
22 |
|
|
|
|
|
|||
|
|
|
|
18 |
|
|
|
|||
|
|
|
|
|
|
|
||||
|
|
|||||||||
|
|
|
|
17 |
|
|||||
|
|
|
|
|
||||||
7 |
||||||||||
|
|
|
|
|
|
|
|
|
||
|
|
6 |
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|||
|
||||||||||
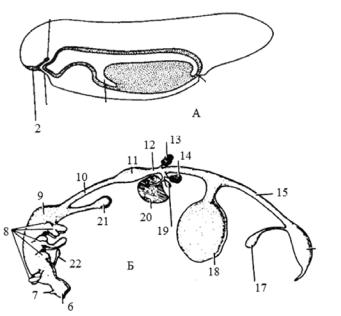
Рис. 3.1. Розвиток травної системи:
А — зародок личинки амфібії, Б — диференціація первинної кишки амніот, 1 — стомодеум, 2 — нюховий епітелій, 3 — гіпофізарний карман, 4 — проктодеум, 5 — печінковий виріст, 6 — ротова порожнина,
7 — карман Ратке, 8 — глоткові кармани, 9 — глотка, 10 — стравохід, 11 — шлунок, 12 — дванадцятипала кишка, 13 — дорсальна підшлункова залоза, 14 — вентральна підшлункова залоза, 15 — тонка кишка,
16 — клоака, 17 — сечовий міхур, 18 — жовтковий мішок, 19 — жовчний міхур, 20 — печінка, 21 — легені, 22 — щитоподібна залоза
(за Ковтун та ін., 2005)
Майже одночасно з цим формується шлунок. Він з’являється як розширення первинної кишки. З диференціацією шлунка кишкова трубка поділяється на передню (краніальніше шлунка) і задню (каудальніше) кишки. Передня частина кишкової трубки, між шлунком
136
і глоткою, стає стравоходом. Проте функцію стравоходу ця частина кишки набуває тільки після з’єднання глотки з ротовою порожниною.
Якщо первинна кишка починає формуватися на стадії гаструли, то ротовий отвір і ротова порожнина закладаються значно пізніше — на стадії, коли починають утворюватися зябра. Первинна ротова порожнина з’являється у вигляді кармана із ектодерми на вентральній стороні головного кінця тіла зародка.
Своїм дном цей карман прилягає до переднього кінця кишкової трубки. Між дном кармана та глотковим кінцем кишкової трубки довгий час зберігається роздільна перетинка, яка руйнується лише на стадії бруньок грудних кінцівок. Після цього глотка з’єднується
зротовою порожниною, а кишкова трубка набуває переднього отвору. Дещо пізніше проривається анальний отвір. Оскільки і передній, і задній отвори утворювалися шляхом вп’ячування ектодерми та змикалися
зентодермою кишкової трубки, то епітелій, який вистилає ці ділянки, має мішану, тобто ектодермальну та ентодермальну природу.
Існують різні підходи до поділу травного тракту на відділи. Згідно
знайпростішою схемою, яка ближче відповідає ситуації у первинноводних хребетних, розрізняють головну кишку (ротова порожнина та глотка) і тулубову кишку (всі інші частини). Складніша схема включає передню, середню та задню кишки. Передня кишка — це ротова порожнина, глотка, стравохід і шлунок (її задня межа — пілоричний сфінктер або отвір жовчної протоки). До складу середньої кишки входять відділи тонкої кишки — дванадцятипала, порожня та клубова (раніше сюди відносили також сліпу кишку). Задня кишка, за сучасними уявленнями, відмежована від середньої кишки клубово-сліпим (ілеоцекальним) клапаном, тобто включає сліпу,
товсту і пряму кишку.
Ротова порожнина та її органи. Функціонально і топографічно ротова порожнина пов’язана з двома системами органів — травною та респіраторною. В історичному аспекті розвиток, будова і функції ротової порожнини пов’язані з еволюційними перебудовами вісцерального скелета.
Ротова порожнина обмежена: спереду і з боків краями щелеп і зубами, якщо вони є; покрівля ротової порожнини у риб і земноводних утворена дном мозкової коробки, кістки якої з деякими покривними
137
кістками утворюють первинне піднебіння. Каудально ротова порожнина обмежена м’яким піднебінням з власною мускулатурою, під- небінно-глотковими дугами, у деяких мавп і людини ще й виступом м’якого піднебіння — язичком; вентрально — елементами під’язикового апарата та м’якими тканинами. Стінки ротової порожнини вистелені епітелієм ектодермального походження; епітелій багатошаровий, іноді частково зроговілий, у земноводних — миготливий.
Отвір у ротову порожнину в більшості хребетних обмежений губами — відносно пластичними утвореннями із епідермісу та сполучної тканини. У круглоротих ротовий отвір круглий, звідки чому походить назва цієї групи; у міксин по його краях є тактильні органи — щупальця, а у міног краї рота утворюють присосок, яким тварина прикріплюється до жертви. Тактильні органи, здебільшого у вигляді вусиків, наявні також у більшості костистих риб. Губи риб, земноводних і плазунів являють собою невеликі шкіряні складки назовні від зубів і щелеп.
Накопичення кератину в епітелії ротового отвору та щелеп можуть перетворювати губи на роговий дзьоб, як у птахів, черепах, деяких нижчих ссавців. У плацентарних ссавців губи добре розвинені та відмежовані від країв щелеп і зубів глибокими канавками, так що утворюється невеликий простір — присінок рота. З розвитком лицевої мускулатури, і, зокрема, кругового м’яза рота, характерного для ссавців, губи ссавців стають дуже рухливими, що дозволяє новонародженим ссавцям смоктати молоко. У ссавців також утворюється певний простір між боковими поверхнями щелеп і щоками, стінки останніх включають елементи лицевої мускулатури. В окремих видів (гризуни, мавпи) простір за щоками розширився, в результаті чого утворилися защічні мішки для накопичення їжі.
Як правило, у більшості хребетних ротовий отвір розміщений попереду голови або зміщений на її черевну поверхню. У селяхій та осетрових розвивається рило, у такому разі ротовий отвір знаходиться каудальніше переднього кінця голови. У наземних тетрапод з’явилася тенденція до розмежування травного та повітряного шляхів, яка завершилася утворенням вторинного кісткового піднебіння.
У ротовій порожнині зустрічаються тверді епідермальні структури. Зокрема, це рогові зубці передротової воронки та язика круглоро-
138
тих, покриви язика птахів і ссавців, нарешті, пластини вусатих китів (китовий вус) — результат зроговіння покривів на піднебінних валиках.
Язик (lingua) круглоротих докорінно відрізняється від такого у риб — це первинно м’язовий орган, тоді як у риб він утворений вкритими шкірою вентральними кінцями вісцеральних дуг (починаючи з під’язикової) і копули, але не містить м’язів. Язик міноги, втягуючись, сприяє присмоктуванню або ж скоблить роговими зубцями тканини жертви. У разі рухів язика вперед, коли ефект присмоктування зникає, мінога фіксується на жертві зубцями передротової лійки. У міксин озброєний загнутими назад зубцями язик захоплює тканини жертви та поздовжніми рухами втягує їх всередину, при зворотному русі язика корм утримується зубцями ротової порожнини.
У риб з’являється складка слизової оболонки, яка підтримується переднім непарним елементом вісцерального скелета — базігіале. Зв’язок з елементом під’язикового апарата свідчить, що ця структура гомологічна язику вищих хребетних. Язик бере участь у просуванні здобичі до стравоходу, хоча головним засобом управління кормовими об’єктами залишається рух води через зміни об’єму ротоглоткової та оперкулярної порожнин.
Язик амфібій важливий для маніпуляції з кормом в межах і, головне, поза ротової порожнини, оскільки його рухами у повітрі вже не можна управляти шляхом змін об’єму порожнин. Язик розвивається попереду від базігіале на дні ротової порожнини як ділянка покривів, на якій закінчуються пучки під’язичних м’язів і утворюються залозисті поля. У хвостатих язик пов’язаний з під’язиковим апаратом, а в безхвостих незалежний від нього, що повністю звільняє під’язиковий апарат для обслуговування гулярного (нагнітаючого) дихання. Язик жаб являє собою мішечок з м’язовими стінками, який прикріплений до нижньої щелепи в області симфізу та наповнюється лімфою. М’яз, який здійснює протракцію язика (висування вперед), є частиною підборідочно-язикового м’яза; ретрактор язика відокремлюється від груднинно-під’язикового. В амфібій, які живуть переважно або постійно у воді язик часто редукується.
Язик амніот розвивається із непарного зачатка, який розміщується в області другої і третьої зябрових дуг. Потім до цього зачатка приєднується пара валиків, що лежать з боків і спереду від під’язикової
139