

Анатомический практикум. Черепные нервы специальной чувствительности Учебное пособие
.pdfсоответствуют оболочкам головного мозга. Так, наружное влагалище зрительного нерва, vagina externa nervi optici, состоит из двух листков: твердой оболочки головного мозга, dura mater, и паутинной оболочки, arachnoidea; внутреннее влагалище зрительного нерва, vagina interna nervi optici, соответствует мягкой оболочке головного мозга, pia mater encephali. У глазного яблока все оболочки нерва срастаются между собой, а также с фиброзной оболочкой глазного яблока. Межвлагалищные пространства соответствуют межоболочечным пространствам и представляют собой соответственно щелевидное субдуральное пространство, spatium subdurale, и подпаутинное пространство, spatium subarachnoidale, в котором содержится спинномозговая жидкость. Межвлагалищные пространства зрительного нерва сообщаются с одноименными пространствами оболочек головного мозга.
Рис. 5. Зрительный нерв, n. opticus, правый, снизу (слева удалены: глазное яблоко, зрительный нерв и часть обонятельного тракта) (по В.П. Воробьёву)
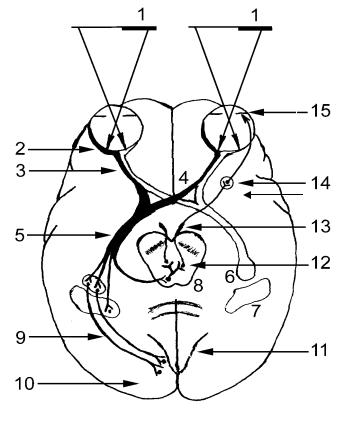
Зрительные нервы кпереди от турецкого седла образуют зрительный перекрест, chiasma opticum, (рис. 6, 4). Перекрёст является частичным, так как здесь перекрещиваются нервные волокна, идущие от внутренних (носовых, или медиальных) половин сетчаток. Волокна же из наружных (височных, или латеральных) половин сетчаток проходят через chiasma opticum неперекрещенными. После перекреста зрительные пути носят название зрительных трактов, tractus opticus, (рис 6, 5). Следует иметь в виду, что при
10
поражении (например, при сдавлении опухолью) всегда выпадают поля зрения, противоположные выпавшему полю сетчатки.
В результате особенностей перекрёста, в зрительном тракте проходят волокна не от одного глаза, как в зрительном нерве, а от одноименных половин сетчаток обоих глаз: так, в левом зрительном тракте содержатся волокна от обеих левых половин сетчаток, в правом зрительном тракте – от обеих правых половин сетчаток.
Рис. 6. Схема зрительных нервов, |
|
||
зрительных путей |
и зрачкового |
|
|
рефлекса: |
|
|
|
1 – поле зрения; 2 – зрительная часть |
|
||
16 |
|||
сетчатки глазного яблока, pars optica |
|||
|
|||
retinae bulbi oculi; 3 – зрительный |
|
||
нерв, n. opticus; 4 – зрительный |
|
||
прекрест, chiasma opticum; 5 – |
|
||
зрительный тракт, tractus opticus; 6 – |
15 |
||
подушка зрительного бугра, pulvinar |
|
||
thalami; 7 – латеральное коленчатое |
|
||
тело, corpus geniculatum laterale; 8 – |
|
||
верхние холмики |
крыши среднего |
|
|
мозга, colliculi superiores tecti mesencephalici; 9 – зрительная лучистость (пучок Грациоле), radiatio optica; 10 – затылочная доля мозга, lobus occipitalis; 11 – шпорная борозда, sulcus calcarinus; 12 –
добавочные ядра глазодвигательного нерва (ядра Эдингера-Вестфаля), nuclei accessorii nervi oculomotorii; 13 – преганглионарные волокна в составе глазодвигательного нерва, neurofibrae preganglionicae; 14 – ресничный узел, ganglion ciliare; 15 – постганглионарные волокна, neurofibrae postganglionicae; 16 – мышца, суживающая зрачок, m. sphincter pupillae
Таким образом, зрительный тракт содержит волокна от латеральных отделов сетчатки одноименной стороны и волокна от медиальных отделов сетчатки противоположной стороны.
Направляясь кзади и кнаружи, зрительный тракт огибает наружную поверхность ножки мозга и, предварительно разделившись на пучки, заканчивается в подкорковых зрительных центрах – подушке зрительного бугра, pulvinar thalami, (рис. 6, 6), латеральном коленчатом теле, corpus geniculatum laterale, (рис. 6, 7) и верхних холмиках пластинки крыши, colliculus superior tecti laminae mesencephalici, (рис. 6, 8), где лежат IV нейроны зрительного пути. Далее в корковые зрительные центры импульсы передаются только из латеральных коленчатых тел. Аксоны нейроцитов их ядер с каждой стороны проходят через
11
задний отдел задней ножки внутренней капсулы, сapsula interna, затем огибают наружную стенку заднего рога бокового желудочка и под названием зрительной лучистости, radiatio optica, (пучок Грациоле) (рис. 6, 9) заканчиваются в корковых центрах зрения. Проекционный центр зрения расположен на медиальной поверхности затылочной доли мозга, по бокам от шпорной борозды, suclus calcarinus, (рис. 6, 10). В глубине шпорной борозды заканчиваются нервные волокна, несущие импульсы от желтого пятна сетчатки, где располагаются колбочки.
Исследования последних лет показали, что верхняя половина сетчатки спроецирована на нейроны верхнего края шпорной борозды, нижняя половина сетчатки – на нейроны нижнего края шпорной борозды. Для коркового конца анализатора характерна группировка нейронов в колонки, идущие в радиальном направлении к поверхности коры. В каждой колонке осуществляется анализ и синтез одного конкретного параметра зрительной информации: размеров предмета, его формы, цвета и удаленности от глаз.
Аксоны нейронов серых слоев верхних холмиков пластинки крыши среднего мозга направляются к добавочным парасимпатическим ядрам (рис. 6, 12) глазодвигательного нерва своей и противоположной стороны и переключаются на его клетках. Таким образом, пучки волокон tractus opticus, направляющиеся к верхним холмикам крыши среднего мозга, входят в состав рефлекторной дуги зрачкового рефлекса. Чувствительной (афферентной) частью этой дуги является фрагмент зрительного пути, от палочек и колбочек сетчатки глаза до серых слоев верхних холмиков крыши среднего мозга (см. выше). Двигательной (эфферентной) частью дуги являются волокна, исходящие из нейроцитов добавочных парасимпатических ядер глазодвигательного нерва (рис. 6, 13), которые прерываются в ресничном узле, ganglion ciliare, (рис. 6, 14). Парасимпатические постганглионарные волокна от клеток ресничного узла подходят к мышце, суживающей зрачок, m. sphincter pupillae, (рис. 6, 16), и к ресничной мышце, m. ciliaris, обеспечивающей аккомодацию (при сокращении мышцы капсула хрусталика расслабляется и изменяется его кривизна; хрусталик становится выпуклым, и это определяет приспособление глаза к ясному видению вблизи).
От верхних холмиков среднего мозга часть волокон идет в нисходящем направлении, образуя крышеспинномозговой путь, tractus tectospinalis, обеспечивающий двигательные рефлекторные реакции на внезапные зрительные раздражения.
Поражение различных участков зрительного пути ведет к определенным нарушениям зрения. Так, при полном нарушении проводимости зрительного нерва наступает слепота (амавроз) на один глаз с одноименной стороны, частичное нарушение проводимости ведет к выпадению отдельных участков полей зрения (рис. 6, 1) (скотома).
Полная двусторонняя слепота наблюдается при разрушении зрительного перекреста. При сдавлении внутренней части зрительного перекреста (например,
12
при опухолях гипофиза) наступает выпадение наружных половин полей зрения
(билатеральная гемианопсия).
Полное поражение зрительного тракта, подкорковых зрительных центров, зрительной лучистости или коркового ядра зрительного анализатора приводит к развитию гемианопсии (половинной слепоты на оба глаза). Если очаг поражения слева, то выпадает правая половина поля зрения обоих глаз; при наличии поражения справа происходит выпадение левой половины поля зрения обоих глаз.
VIII пара черепных нервов – преддверно-улитковый нерв, nervus vestubulocochlearis, (рис. 7) является нервом специальной чувствительности, содержит чувствительные волокна, несущие информацию от преддверноулиткового органа (органа равновесия и слуха). Нерв состоит из двух различных по своим функциям нервов: преддверного нерва, nervus vestibularis, и улиткового нерва, nervus cochlearis.
Каждому нерву принадлежит чувствительный узел, отростки биполярных нейронов которого идут в двух направлениях. Периферические отростки образуют ветви нервов, идущие к рецепторам. Центральные отростки формируют корешки нервов, направляющиеся в головной мозг.
Nervus vestibularis является проводником импульсов от статокинетического аппарата (органа равновесия, органа гравитации или вестибулярного аппарата), заложенного в перепончатом лабиринте внутреннего уха в пирамиде височной кости. Статокинетический аппарат включает в себя эллиптический мешочек (маточка, utriculus), сферический мешочек (мешочек, sacculus) и три полукружных протока, ductus semicirculares membranacei,
расположенных в трех взаимно перпендикулярных плоскостях (рис. 8).
Рис. 7. Преддверно-улитковый нерв (по A. Leblanc)
13
Рецепторными органами эллиптического и сферического мешочков являются неподвижные пятна, maculae, (рис. 8), содержащие рецепторные волосковые клетки, cellulae pilosae, основания которых плотно охватываются терминалями афферентных волокон биполярных нейроцитов вестибулярного ганглия. Макула эллиптического мешочка, macula utriculi, занимает его основание и расположена параллельно основанию черепа; макула сферического мешочка, macula sacculi, расположена на его медиальной стенке в вертикальном положении.
Волоски рецепторных клеток соприкасаются со студенистой мембраной статоконий, membrana statoconiorum, содержащей статоконии (отолиты) – мельчайшие кристаллы карбоната кальция.
Плотность мембраны статоконий (отолитовой мембраны) существенно выше, чем у эндолимфы. Эллиптический мешочек, utriculus, и сферический мешочек, sacculus, – это отолитовые, или статолитовые органы, основная функция которых – регуляция статического равновесия, т. е. равновесия головы и тела, находящихся в покое. Адекватным стимулом для рецепторных клеток utriculus и sacculus является сила тяжести (именно с этим связано чувство земного тяготения). Гравитационное ускорение представляет собой особую форму линейных ускорений. При невесомости (во время космических полетов) устраняется действие силы тяжести на статоконии. Рецепторные волосковые клетки маточки и мешочка сигнализируют об изменении положения тела по отношению к центру земной тяжести (к гравитационному полю). Рецепторы маточки наиболее чувствительны к изменению положения головы в пространстве, рецепторы мешочка наиболее чувствительны к восприятию вибрационных колебаний.
Три полукружных протока связаны с эллиптическим мешочком - маточкой, utriculus, и каждый из них оканчивается расширением – ампулой, ampula.
В ампулах находятся производные соединительнотканной пластинки – ампулярные гребешки, cristae ampullares. Гребешки покрыты эпителием, содержащим рецепторные волосковые клетки. Волоски рецепторных клеток каждого ампулярного гребешка погружены в студенистое вещество, имеющее форму высокого узкого купола - купулы, cupula, (рис. 8, 2). Купулы ампул полукружных каналов обладают одинаковой плотностью с эндолимфой перепончатого лабиринта внутреннего уха, так как не содержит статоконий и поэтому при линейном ускорении не смещаются.
Полукружные протоки регулируют динамическое равновесие тела, то есть равновесие тела, движущегося в пространстве. Кинетические рецепторы ампул полукружных протоков чрезвычайно чувствительны к поворотам головы.
Они обеспечивают человеку способность определять положение тела в пространстве:
при повороте головы и тела вокруг вертикальной оси;
при наклоне головы и тела вперед и назад вокруг сагитальной оси;
14
при наклоне головы и тела влево и вправо вокруг фронтальной оси.
Кроме того, рецепторы ампул передают немедленную информацию об угловых ускорениях при всевозможных сложных двигательных комбинациях в любой плоскости. При вращательном (угловом) ускорении, т. е. при повороте головы, полукружные каналы, естественно, поворачиваются вместе с ней. Благодаря тому, что эндолимфа в полукружных протоках в силу своей инерции сохраняет прежнее положение (остается на месте), по обе стороны купулы возникает разность давлений и свободный конец купулы отклоняется в направлении, противоположном повороту. Инерционное смещение эндолимфы соответственно возникает в том перепончатом полукружном протоке, который находится в плоскости движения тела в пространстве. Угловое ускорение (смещение) вызывает сдвиг волосков рецепторных клеток, являясь адекватным стимулом, и изменяет активность терминалей афферентного нерва.
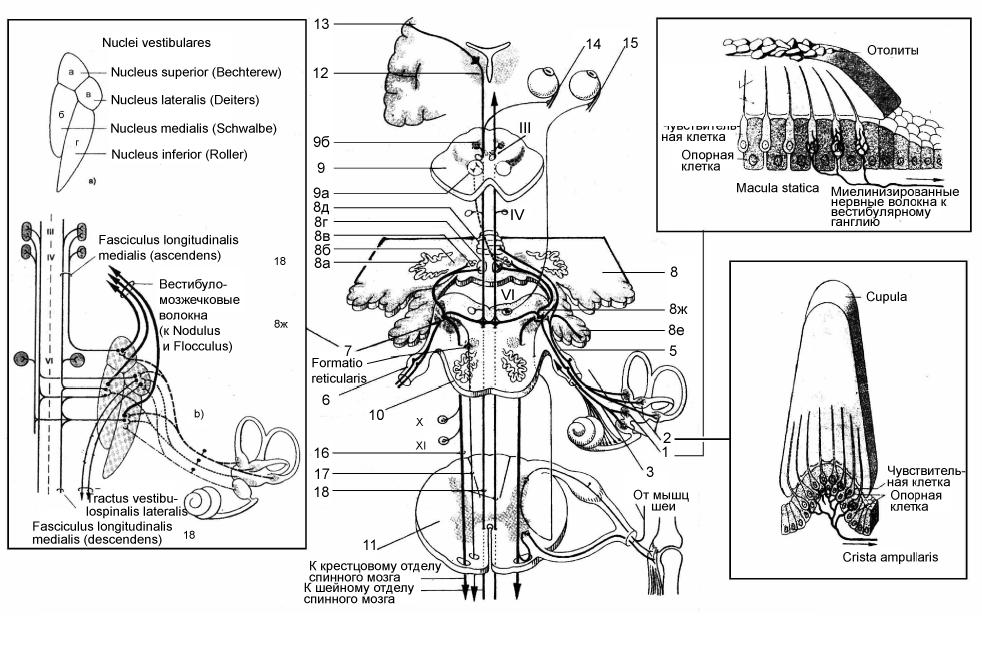
Чувствительный узел преддверного нерва, ganglion vestibulare, (рис. 8), лежит на дне внутреннего слухового прохода височной кости, meatus acusticus internus. Узел содержит биполярные нейроны (I нейроны вестибулярного пути). Периферические отростки (рис. 8, 3, 4) биполярных нейроцитов преддверного узла направляются к рецепторным органам перепончатого лабиринта внутреннего уха и группируются в афферентные нервы, носящие название тех образований, к которым они подходят. Различают следующие нервы:
n. utriculoampullaris – эллиптически-мешотчато-ампулярный нерв;
n. utricularis – эллиптически-мешотчатый нерв (нерв эллиптического мешочка (маточки) преддверия перепончатого лабиринта внутреннего уха);
n. saccularis – сферически-мешотчатый нерв (нерв сферического мешочка преддверия перепончатого лабиринта внутреннего уха);
n. ampullaris anterior – передний ампулярный нерв (нерв ампулы переднего полукружного протока внутреннего уха);
n. ampullaris lateralis – латеральный ампулярный нерв (нерв ампулы латерального полукружного протока внутреннего уха);
n. ampullaris posterior – задний ампулярный нерв (нерв ампулы заднего перепончатого полукружного протока внутреннего уха).
Раздражения волосковых чувствительных клеток пятен и гребешков воспринимаются чувствительными окончаниями соответствующих нервов, и импульс передается к телам биполярных нейронов преддверного узла, далее – на их центральные отростки. Совокупность центральных отростков представляет собой преддверный корешок, radix vestibulare.
Центральные отростки (рис. 8, 5) в составе преддверного корешка покидают внутреннее ухо, идут во внутреннем слуховом проходе и через внутреннее слуховое отверстие попадают в полость черепа; входят в ствол головного мозга в области мосто-мозжечкового угла и направляются к area
15
vestibularus ромбовидной ямки, где в покрышке моста лежат вестибулярные ядра, содержащие II нейроны вестибулярного пути (рис. 8, 7а, б, в, г):
медиальное ядро (Швальбе) – nucl. vestibularis medialis (Schwalbe’s);
латеральное ядро (Дейтерса) – nucl. vestibularis lateralis (Deiters’);
верхнее ядро (Бехтерева) – nucl. vestibularis superior (Bechterew);
нижнее ядро (ядро Роллера) – nucl. vestibularis inferior (Roller’s).
Аксоны биполярных нейроцитов ganglion vestibulare (I нейронов вестибулярного пути) образуют восходящие и нисходящие пучки. Восходящие заканчиваются в верхнем вестибулярном ядре, нисходящие – в трех остальных ядрах. От вестибулярных ядер волокна идут в нескольких направлениях:
к мозжечку (к узелку и клочку) – tractus vestibulocerebellaris (рис. 8,
8ж); от мозжечка к вестибулярным ядрам идут волокна в обратном направлении и, таким образом, между ними устанавливается тесная связь, а ядро шатра мозжечка становится важнейшим вестибулярным центром;
к спинному мозгу – tractus vestibulospinalis (рис. 8, 16);
к ядрам III, IV и VI пар черепных нервов – в составе медиального продольного пучка, fasciculus longitudinalis medialis, который начинается от ядра Даркшевича (рис. 8, 9б) и связывает ядра III, IV, VI пар; эти связи лежат в основе рефлексов, осуществляющих компенсирующие установки глаз, т. е. сохранение направления взгляда при перемене положения головы (рис. 8);
к ядрам IX и X пар черепных нервов волокна идут через ретикулярную формацию. Эти связи определяют вестибуловисцеральные реакции (кинетозы); сильное раздражение вестибулярного аппарата часто вызывает неприятные ощущения: головокружение, рвоту, усиленное потоотделение, тахикардию; в таких случаях говорят об укачивании, «морской болезни»;
к коре полушарий большого мозга от вестибулярных ядер идет перекрещенный путь через таламус (в центральных ядрах которого лежат III нейроны пути); от этих нейронов таламуса волокна идут в корковый конец вестибулярного анализатора, рассеянный, как считается, в средней и нижней височных извилинах, где обрабатывается вестибулярная информация – для осознания положения головы в пространстве.
Перечисленные здесь основные связи дают возможность вестибулярной системе играть центральную роль в генерировании двигательной эфферентации, обеспечивающей поддержание нужного положения тела и соответствующие глазодвигательные реакции; при этом вертикальная поза и походка определяются главным образом отолитовым аппаратом, тогда как полукружные каналы управляют в основном направлением взгляда.
16
Статоконии
( )
Мембрана статоконий
Пятно 
Купула
( )
17
Ампулярный гребешок
Рис. 8. Схема строения вестибулярной системы (по П. Дуусу с изменениями и дополнениями
Рис. 8. Продолжение:
1 – пятна сферического и эллиптического мешочков преддверия перепончатого лабиринта, maculae sacculi et utriculi; 2 – гребешки ампул полукружных протоков, cristae ampullares ductus semicirculares; 3 - n. utricularis et n. saccularis; 4 – nn. ampullares: 5 – вестибулярный нерв, n. vestibularis;6 –
преддверный узел, ganglion vestibulare( Scarpa’s); 7 – вестибулярные ядра: 7 а – верхнее; 7 б – медиальное; 7 в – латеральное; 7 г – нижнее; 8 – мозжечок, cerebellum; 8 a – nucleus dentatus; 8 б – nucleus emboliformis; 8 в – nucleus globosus; 8 г – nucleus fastigii; 8 д – червь, vermis; 8 е – клочок, flocculus; 8 ж – tractus vestibulocerebellaris; 9 – cредний мозг, mesencephalon; 9 a – красное ядро, nucleus ruber; 9 б – ядро Даркшевича; 10 – мост, pons (Varolus); 11 – спинной мозг, medulla spinalis; 12 – thalamus; 13 – кора полушарий большого мозга; 14 – m. rectus medialis; 15 – m. rectus lateralis; 16 – tractus vestibulospinalis; 17 – tractus reticulospinalis; 18 – медиальный продольный пучок, fasciculus longitudinalis medialis; III – глазодвигательный нерв, n. oculomotorius; IV –
блоковый нерв, n. trochlearis; VI – oтводящий нерв, n. abducens; IX – языкоглоточный нерв, n. glossopharyngeus; X – блуждающий нерв, n. vagus
Nervus cochlearis (улитковый нерв) является проводником слуховых импульсов от спирального органа улитки (Кортиев орган, organum spirale Corti), заложенного в перепончатом лабиринте внутреннего уха. Спиральный орган воспринимает звуки и трансформирует механические волновые колебания в электрические потенциалы действия нервного импульса. Рецепция звука осуществялется слуховой системой, состоящей из наружного, среднего и наиболее сложноустроенного внутреннего уха.
Ухо человека воспринимает звуковые волны с частотой колебаний от 16 до 20 000 Гц, человеческая речь имеет оптимальные границы от 1000 до
4000 Гц (рис. 9 в, 15).
Воздушные волны направляются в наружный слуховой проход, ударяются о барабанную перепонку, membrana tympani, и вызывают ее вибрацию. Барабанная перепонка является латеральной стенкой барабанной полости. Степень натяжения барабанной перепонки регулируется сокращением m. tensor tympani (иннервируется V парой черепных нервов).
Колебания барабанной перепонки приводят в движение цепь слуховых косточек (молоточек, наковальню, стремя) среднего уха. Основание стремени вдается в окно преддверия, fenestra vestibuli, расположеное на медиальной стенке барабанной полости, paries labyrinthicum. Амплитуда смещения стремени в окошке преддверия регулируется сокращением m. stapedius (иннервируется VII парой черепных нервов). В костном канале улитки, повторяя его 2,5 оборота, расположен улитковый лабиринт, включающий три пространства: лестницу преддверия, scala vestibuli, улитковый проток, ductus cochlearis, и барабанную лестницу, scala tympani, (рис. 9 б, в).
18
В пространствах scala vestibuli и scala tympani циркулирует перилимфа. Обе лестницы, находящиеся соответственно выше и ниже ductus cochlearis, сообщаются друг с другом в области верхушки улитки через ее отверстие - геликотрему, helicotrema. Улитковый проток представляет собой часть перепончатого лабиринта внутреннего уха, который начинается слепым выпячиванием в преддверии и слепо заканчивается на верхушке улитки; ductus cochlearis заполнен эндолимфой. Вдоль всего улиткового протока на базилярной пластинке располагается спиральный орган, содержащий рецепторы слухового анализатора. Благодаря тому, что основание стремени обращено в преддверную лестницу, его механические колебания смещают содержащуюся в ней перилимфу. Звуковые волны по перилимфе лестницы преддверия поднимаются вверх до верхушки улитки и передаются через helicotrema – на перилимфу барабанной лестницы, по которой спускаются вниз к круглому отверстию, fenestra rotundum, улитки, и гасятся о вторичную барабанную перепонку, membrana tympani secundaria, натянутую по его окружности.
По мере прохождения звуковых волн по перилимфе лестницы преддверия начинает колебаться преддверная стенка, paries vestibularis (membrana vestibularis Reissneri), и колебания передаются эндолимфе улиткого протока, ductus cochlearis. Перемещения эндолимфы вызывают смещение желеобразной покровной мембраны, membrana tectoria, соприкасающейся со стереоцилиями рецепторных волосковых клеток спирального органа.
Кроме того, колебания эндолимфы в его тоннелях вызывают вибрацию натянутых в виде струн соединительнотканных волокон базилярной пластинки,
расположенной на барабанной стенке, paries tympanicus (membrana spiralis)
улиткового протока.
Чувствительный узел улитковой части VIII пары – спиральный узел улитки, ganglion spirale cochleae, (рис. 9 в, 1), располагается в костной спиральной пластинке, образующей вокруг стержня улитки, modiolus cochlearis, завиток в два с половиной оборота. В спиральном узле улитки располагаются биполярные нейроны – первые нейроны слухового пути.
Периферические отростки (дендриты) (рис. 9 в, 2) биполярных клеток спирального узла улитки направляются к рецепторным вторичночувствующим (сенсорно-эпителиальным) клеткам кортиевого органа. Терминали дендритов образуют синапсы с основаниями этих клеток. В результате смещения рецепторных клеток возникают цитохимические процессы, вызывающие трансформацию энергии внешнего раздражения в нервный импульс, который по дендритам биполярных нейронов передается на их тела. Далее – по центральным отросткам (аксонам) этих нейронов импульс передается в головной мозг.
Центральные отростки (аксоны) биполярных нейронов, собираясь в улитковый корешок (нерв) VIII пары покидают внутреннее ухо, идут во внутреннем слуховом проходе и попадают в полость черепа через
19