


ГЛАВА 2. ФАКТОРЫ ПРОТИВОМИКРОБНОЙ |
внутрь организма, так как клетки эпителия тесно при- |
ЗАЩИТЫ ОРГАНИЗМА ЧЕЛОВЕКА |
легают друг к другу. |
Весь комплекс защитных факторов организма чело- |
Так как эпидермис состоит из нескольких слоев, из |
века от возбудителей инфекционных заболеваний |
которых поверхностный представлен мертвыми оро- |
можно условно поделить на три эшелона: физиологи- |
говевшими клетками (чешуйками), то он практически |
ческие барьеры, система врожденного иммунитета и |
непроходим для большинства микробов. Более того |
система адаптивного (приобретенного) иммунитета |
постоянное слущивание чешуек при обновлении эпи- |
(рисунок 2.1). Для того чтобы вызвать заболевание, |
дермиса обеспечивает дополнительное очищение его |
возбудитель сначала должен преодолеть барьеры. Ес- |
поверхности от микробов. Поэтому инфицированию |
|
|
ли это происходит (по причине дефекта барьеров или |
человека через кожу, как правило, предшествует трав- |
|
|
из-за наличия у микроба факторов патогенности, спо- |
ма (чаще микротравма, не заметная невооруженным |
|
|
собствующих разрушению или нейтрализации этих |
глазом), вследствие чего образуются дефекты в эпи- |
|
|
барьеров), то активируются системы врожденного, а |
дермисе, через которые и проникают микробы. |
|
|
затем и приобретенного иммунитета. Существует про- |
Слизистые оболочки представлены однослойным |
|
|
стая образная аналогия со средневековым замком, ко- |
или многослойным неороговевающим эпителием, что |
|
|
торая доступно объясняет функционирование защит- |
облегчает проникновение микробов. Однако имеются |
|
|
ных сил организма человека в ходе нейтрализации па- |
дополнительные защитные механизмы, обеспечиваю- |
|
|
тогенов. Физиологические барьеры при этом сравни- |
щие механическое очищение слизистых от микробов: в |
|
|
ваются со рвом и стенами, окружающими замок; сис- |
ЖКТ это перистальтика, а в воздухоносных путях – |
|
|
тема врожденного иммунитета – с жителями замка, |
слизь и биение ресничек. Например, турбулентные по- |
|
|
способными противостоять врагу; а система адаптив- |
токи воздуха в дыхательных путях обеспечивают при- |
|
|
ного иммунитета – с профессиональными воинами, ко- |
липание микробов к вырабатываемой бокаловидными |
|
|
торые руководят обороной и решают самые трудные |
клетками слизи, которая впоследствии эвакуируется за |
|
|
задачи. |
счет биения ресничек наружу, поэтому у здорового че- |
|
|
|
ловека воздухоносные пути стерильны, уже начиная с |
2.1. Физиологические барьеры |
бронхов 3-4 порядка. |
|
|
Физиологические барьеры – это комплекс защит- |
|
ных механизмов, не позволяющих патогену внедрить- |
Химические барьеры |
|
|
ся в восприимчивый макроорганизм. Принято выде- |
Экзокринные железы кожи, слизистой желудка, пе- |
|
|
лять три группы барьеров: механические, химические |
чень и поджелудочная железа вырабатывают химиче- |
|
|
и биологические. |
ские вещества, блокирующие размножение микробов. |
|
|
|
Сальные железы кожи вырабатывают жирные кисло- |
Механические барьеры |
ты, снижающие pH кожи, и таким образом создают не- |
|
|
Кожа (точнее эпидермис) и слизистые оболочки ме- |
благоприятные условия для размножения ряда микро- |
|
|
ханически препятствуют проникновению микробов |
организмов. Потовые железы секретируют большое |
|
|
|
Рисунок 2.1. Компоненты и систе- |
|
мы, обеспечивающие противоин- |
|
фекционную защиту человека. Для |
|
попадания в макроорганизм микробы |
|
должны преодолеть многочисленные |
|
физиологические барьеры. Неболь- |
|
шая часть возбудителей, которая ус- |
|
пешно преодолевает барьеры, распо- |
|
знается и нейтрализуется системами |
|
врожденного и адаптивного иммуни- |
|
тета. |
Зорников Д.Л., Литусов Н.В. Основы противоинфекционной иммунологии |
Страница 3 |

количество солей и других осмотически активных ве- |
опасного биоматериала (в том числе и микробов). При |
|
ществ, что приводит к потере воды бактериальными |
этом, исходя из особенностей функционирования, вы- |
|
клетками. Обкладочные клетки собственных желез |
деляют две системы иммунитета: врожденного и адап- |
|
желудка продуцируют соляную кислоту, за счет чего |
тивного (приобретенного) (подробно о данных систе- |
|
pH в желудке снижается до 1-2, что является губитель- |
мах смотри в главе 4). |
|
ным для большинства микроорганизмов. Секреты пе- |
Основные различия врожденного и адаптивного |
|
чени и поджелудочной железы содержат желчные ки- |
иммунитета представлены в таблице 2.1. Главное раз- |
|
слоты и ряд гидролитических ферментов, блокирую- |
личие заключается в том, что первый появляется и |
|
щих рост и размножение микроорганизмов. |
функционирует сразу после рождения человека (отсю- |
|
|
|
да название врожденный иммунитет), а другой – толь- |
Биологические барьеры |
ко после предварительного антигенного стимула, и та- |
|
Микроорганизмы, в норме колонизирующие биото- |
ким образом он появляется в течение жизни человека, |
|
пы человека, защищают его от патогенных микробов |
после того как он встретится с данным антигеном (от- |
|
посредством ряда механизмов: |
сюда название приобретенный или адаптивный имму- |
|
конкуренции за площадь адгезии; |
нитет). |
|
конкуренции за питательный субстрат; |
В функционировании систем врожденного и адап- |
|
модификации среды за счет выработки кис- |
тивного иммунитета принципиально можно выделить |
|
|
лых метаболитов; |
три отличия: специфичность действия, скорость и сте- |
|
продукции бактериоцинов (убивающих дру- |
пень реагирования на стимул. Система врожденного |
|
гие бактерии веществ); |
иммунитета действует не специфически, то есть реа- |
продукции активных форм кислорода; |
гирует на любой микробный стимул. Поэтому в неко- |
|
стимуляции иммунной системы человека. |
торых (особенно в отечественных) литературных ис- |
|
Совокупность этих механизмов позволяет эффек- |
точниках применяют термины «факторы неспецифи- |
|
|
||
тивно защищаться от многих патогенов. В этом плане |
ческой защиты» или «факторы неспецифической рези- |
|
|
||
показателен эксперимент с гнотобионтами – живот- |
стентности». Данная система реагирует на микробный |
|
|
||
ными, выращенными в максимально возможных асеп- |
стимул незамедлительно и с одинаковой интенсивно- |
|
|
||
тических |
условиях. При попадании гнотобионтов в |
стью вне зависимости от кратности стимулов. В то |
|
||
микробную среду фиксировали гипофункцию их им- |
время как система адаптивного иммунитета действует |
|
|
||
мунной системы, так как для ее полноценного разви- |
сугубо специфично, для ее активации и развития им- |
|
|
||
тия необходимы постоянные микробные стимулы. |
мунного ответа при первой встрече с антигеном требу- |
|
|
||
|
|
ется достаточно длительное время, зато при повтор- |
2.2. Врожденный и адаптивный иммунитет |
ной стимуляции данным антигеном адаптивный им- |
|
|
||
Человек обладает иммунной системой, обеспечи- |
мунный ответ развивается быстрее и интенсивнее. Эту |
|
|
||
вающей |
нейтрализацию чужеродного потенциально |
способность системы адаптивного иммунитета обычно |
|
||
называют иммунологической памятью.
Таблица 2.1. Отличия в функционировании систем врожденного и адаптивного иммунитета. Врожденный иммунитет
формируется еще до встречи с патогеном и является неспецифичным, для его развития при первом попадании микроба в организм не требуется много времени. Адаптивный иммунитет развивается напротив только после антигенной стимуляции и является высокоспецифичным, при первой встрече с патогеном происходит активация и развитие иммунного ответа, что требует значительного времени. При повторной встрече с тем же антигеном адаптивный иммунитет развивается быстрее и интенсивнее, чем при первой встрече, за счет иммунологической памяти.
Зорников Д.Л., Литусов Н.В. Основы противоинфекционной иммунологии |
Страница 4 |
ГЛАВА |
3. |
КЛЕТКИ И |
ОРГАНЫ |
ИММУННОЙ инкапсулированные скопления лимфоидной ткани, ас- |
||
СИСТЕМЫ |
|
|
|
|
социированные с кожей или слизистыми оболочками. |
|
|
|
|
|
Например, миндалины глотки, сгруппированные в |
||
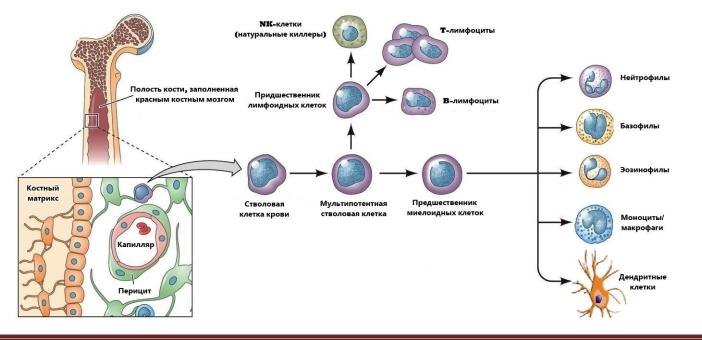
Все клетки иммунной системы человека происходят |
||||||
вальдейерово кольцо, пейеровы бляшки, расположен- |
||||||
из единого предшественника – стволовой клетки кро- |
||||||
ные в подслизистом слое подвздошной кишки или |
||||||
ви (рисунок 3.1). Соответственно, все предшественни- |
||||||
лимфоидная ткань аппендикса. Функция данных скоп- |
||||||
ки клеток иммунной системы образуются у человека в |
||||||
лений такая же, как у селезенки и лимфатических уз- |
||||||
красном костном мозге. Однако на различных стадиях |
||||||
лов. |
||||||
дифференцировки многие клетки мигрируют в другие |
||||||
В красном костном мозге происходит пролиферация |
||||||
органы и ткани. |
|
|
|
|||
|
|
|
и определенные этапы дифференцировки всех клеток |
|||
Часть клеток иммунной системы формирует опре- |
||||||
иммунной системы. Все миелоидные клетки и B- |
||||||
деленные органы (которые, однако, могут выполнять |
||||||
лимфоциты проходят в костном мозге полную диффе- |
||||||
не только иммунную функцию), тогда как другая часть |
||||||
ренцировку, а затем мигрируют в кровь и ткани. T- |
||||||
находится в периферической крови или распределена |
||||||
лимфоциты уже на ранних стадиях дифференцировки |
||||||
по всем тканям организма. |
|
|
||||
|
|
покидают костный мозг и мигрируют в тимус. |
||||
|
|
|
|
|
||
3.1. Функции органов иммунной системы |
В тимусе происходит дифференцировка T- |
|||||
лимфоцитов. Основная задача тимуса – провести се- |
||||||
Большая часть клеток иммунной системы сгруппи- |
||||||
лекцию («обучение») T-лимфоцитов (рисунок 3.3). |
||||||
рованы в лимфоидные органы и ткани (рисунок 3.2). В |
||||||
Сначала мигрировавшие из костного мозга предшест- |
||||||
зависимости от |
назначения |
выделяют |
первичные |
|||
венники T-лимфоцитов проходят позитивную селек- |
||||||
(центральные) и вторичные (периферические) органы |
||||||
цию: отбираются только лимфоциты, способные взаи- |
||||||
иммунной системы. В первичных органах происходит |
||||||
модействовать с антигенпрезентирующими рецепто- |
||||||
пролиферация и основная дифференцировка клеток |
||||||
рами индивидуума (смотри 5.4. Презентация антиге- |
||||||
иммунной системы. К данным органам относят крас- |
||||||
нов), в противном случае лимфоциты подвергаются |
||||||
ный костный мозг и тимус. В свою очередь, во вторич- |
||||||
программированной смерти – апоптозу. На втором |
||||||
ные органы мигрируют (из первичных), как правило, |
||||||
этапе происходит негативная селекция T-лимфоцитов: |
||||||
уже дифференцированные клетки. Вторичными орга- |
||||||
отбираются только те клетки, которые не способны |
||||||
нами являются селезенка и лимфатические узлы: здесь |
||||||
распознавать собственные антигены, тогда как ауто- |
||||||
происходит |
контакт иммунокомпетентных клеток с |
|||||
реактивные лимфоциты подвергаются апоптозу. В ко- |
||||||
различными антигенами. Кроме первичных и вторич- |
||||||
нечном итоге полную дифференцировку проходят |
||||||
ных органов в организме человека присутствуют не- |
||||||
около 1-5% мигрировавших из костного мозга предше- |
||||||
|
|
|
|
|
||
Рисунок 3.1. Происхождение клеток иммунной системы. Все типы клеток иммунной системы продуцируются в красном
костном мозге из единого предшественника – стволовой клетки крови. На ранних этапах дифференцировки часть стволовых клеток дифференцируется в предшественники лимфоидных клеток, из которых в дальнейшем происходят все лимфоциты. Другая часть стволовых клеток дифференцируется в предшественники миелоидных клеток, из которых впоследствии происходят гранулоциты (нейтрофилы, базофилы, эозинофилы), дендритные клетки и моноциты. Последние после миграции в ткани окончательно дифференцируются в макрофаги.
Зорников Д.Л., Литусов Н.В. Основы противоинфекционной иммунологии |
Страница 5 |
|
Полиморфноядерные лейкоциты (гранулоциты) |
|||
|
Данная группа клеток морфологически характери- |
|||
|
зуется наличием гранул в цитоплазме и разной формой |
|||
|
ядер, которая определяется степенью дифференци- |
|||
|
ровки. В зависимости от окраски цитоплазматических |
|||
Рисунок 3.2. Органы иммунной системы. Красным |
гранул выделяют три типа гранулоцитов: нейтрофи- |
|||
лы, базофилы, |
эозинофилы. Дифференцировка всех |
|||
шрифтом обозначены первичные органы, в которых про- |
||||
гранулоцитов проходит в костном мозге, после чего |
||||
исходит пролиферация и основная дифференцировка кле- |
||||
ток иммунной системы. Синим шрифтом – вторичные ор- |
они выходят в периферическую кровь. Гранулоциты |
|||
ганы, в которых мигрировавшие из первичных органов |
являются короткоживущими клетками – в крови они |
|||
клетки иммунной системы распознают антигены и могут |
||||
циркулируют от нескольких часов до нескольких дней. |
||||
проходить окончательную дифференцировку. Зеленым |
||||
|
|
|
||
шрифтом обозначен пример неинкапсулированной лим- |
В течение этого времени они могут мигрировать в |
|||
фоидной ткани – миндалины. |
ткани при наличии в последних воспалительного оча- |
|||
|
||||
ственников T-лимфоцитов. |
га. Если по прошествии нескольких дней не происхо- |
|||
Лимфатические узлы расположены по всему телу, |
дит миграции гранулоцитов в ткани – они разрушают- |
|||
что позволяет дренировать лимфу и тканевую жид- |
ся. |
|
|
|
кость из всех тканей организма. Лимфа попадает в |
Нейтрофилы составляют до 95% от всех гранулоци- |
|||
лимфатический узел через афферентные сосуды, про- |
тов крови. Основной функцией данных клеток являет- |
|||
ходит через паренхиму, представленную лимфоцитами |
ся фагоцитоз микробов. В гранулах нейтрофилов со- |
|||
и мигрировавшими дендритными клетками, и выходит |
держатся бактерицидные вещества (дефензины, лизо- |
|||
через единственный эфферентный сосуд. Таким обра- |
цим, лактоферрин и др.), которые способствуют обез- |
|||
зом происходит «фильтрация» лимфы от микробных |
вреживанию и эффективному перевариванию микро- |
|||
клеток и их дериватов. Очищенная лимфа по лимфати- |
бов после их поглощения. |
|
||
ческим сосудам попадает в более крупные лимфатиче- |
Эозинофилы и базофилы присутствуют в крови в |
|||
ские узлы, где снова происходит ее фильтрация. В ко- |
гораздо меньших количествах, чем нейтрофилы. Дан- |
|||
нечном итоге стерильная лимфа через грудной лимфа- |
ные клетки также обладают способностью к фагоцито- |
|||
тический проток попадает в кровь. |
зу, однако это не является их прямой функцией. |
|||
Селезенка функционирует по тому же принципу, |
Эозинофилы |
обеспечивают |
иммунитет против |
|
что и лимфатические узлы. Поступающая в селезенку |
крупных паразитов, таких как гельминты. Гранулы эо- |
|||
кровь проходит через белую пульпу, представленную |
зинофилов включают различные микробоцидные ве- |
|||
лимфоцитами, фолликулярными дендритными клет- |
щества. В отличие от нейтрофилов, эозинофилы не по- |
|||
ками и макрофагами, что обеспечивает ее очищение от |
глощают микробы, вместо этого они высвобождают |
|||
микробных антигенов. Таким образом, селезенка обес- |
содержимое своих гранул (дегранулируют) после кон- |
|||
печивает дополнительную защиту от микробов, пре- |
такта с микробом (смотри 6.1.3. Клеточная цитоток- |
|||
одолевших лимфатические узлы или попавших в |
сичность). Такая тактика позволяет противодейство- |
|||
кровь, минуя лимфатические узлы (например, при па- |
вать возбудителям, которые значительно превышают |
|||
рентеральном или трансмиссивном инфицировании). |
по размерам клетки организма, и соответственно не |
|||
Неинкапсулированная лимфоидная ткань локали- |
могут быть фагоцитированы. |
|
||
зована под слизистыми оболочками или под кожей – то |
Базофилы или схожие с ними тучные клетки (схо- |
|||
есть в местах наиболее интенсивного контакта с мик- |
жие по функциям, однако отличающиеся по происхож- |
|||
робами, что также обеспечивает защиту организма от |
дению) содержат в своей цитоплазме гранулы с раз- |
|||
инфекции. |
личными биологически активными веществами, наи- |
|||
|
|
|||
|
|
|||
Зорников Д.Л., Литусов Н.В. Основы противоинфекционной иммунологии |
Страница 6 |
|||

более важным и изученным из которых является гис- |
клетки являются основными антигенпрезентирующи- |
||
тамин. Базофилы присутствуют в крови в небольших |
ми клетками (antigen-presenting cells – APC) и презен- |
||
количествах, тогда как тучные клетки являются осед- |
тируют T-лимфоцитам большую часть антигенов. То- |
||
лыми и обитают в тканях организма. Оба типа клеток |
гда как другие APC: макрофаги, моноциты и B- |
||
высвобождают гистамин во внеклеточное пространст- |
лимфоциты менее эффективно презентируют антиге- |
||
во после контакта со специфическими лигандами |
ны T-лимфоцитам. Дендритные клетки способны по- |
||
(смотри в 4.1. Элементы врожденного иммунитета и |
глощать микробы и их дериваты не только фагоцито- |
||
5.2. Активация фагоцитов и эндотелия). Гистамин спо- |
зом, но и макропиноцитозом, что позволяет им «пере- |
||
собствует увеличению проницаемости эндотелия и тем |
рабатывать» большие объемы тканевой жидкости. |
||
самым ускоряет выход гуморальных и клеточных про- |
|
|
|
тивомикробных факторов в воспалительный очаг. |
Лимфоциты |
|
|
|
Лимфоцитами называют все лейкоциты лимфоид- |
||
Моноциты, макрофаги и дендритные клетки |
ного происхождения. Основные этапы дифференци- |
||
Моноциты образуются в костном мозге и выходят в |
ровки лимфоциты проходят в первичных органах им- |
||
кровоток. Они отличаются от других лейкоцитов круп- |
мунной системы, после чего мигрируют во вторичные |
||
ными размерами и характерным подковообразным |
органы, где происходит распознавание поступающих в |
||
ядром. Моноциты присутствуют в крови всего не- |
организм антигенов. С током крови лимфоциты могут |
||
сколько суток, после чего мигрируют в ткани, где |
рециркулировать по различным органам и тканям, что |
||
окончательно дифференцируются в оседлые макрофа- |
обеспечивает более эффективное распознавание анти- |
||
ги. В зависимости от клеточного окружения в различ- |
генов. |
|
|
ных тканях, макрофаги имеют различную форму и на- |
Именно лимфоциты обеспечивают развитие адап- |
||
звания (например, альвеолярные макрофаги – в легких |
тивного иммунного ответа. Лимфоциты – это единст- |
||
и клетки Купфера – в печени), однако выполняют оди- |
венные клетки в организме человека, способные рас- |
||
наковые функции. Функциями макрофагов является |
познавать антигены (смотри 5.5. Активация T и B- |
||
фагоцитоз микробов и других чужеродных соедине- |
лимфоцитов). В зависимости от места дифференци- |
||
ний, а также клеточного детрита. Кроме фагоцитоза |
ровки и выполняемых функций выделяют три популя- |
||
макрофаги (и в меньшей степени моноциты) выпол- |
ции лимфоцитов: T-лимфоциты, B-лимфоциты, NK- |
||
няют функции презентации антигена T-лимфоцитам и |
клетки (функции этих клеток смотри в 4.1. Элементы |
||
активации эндотелия с последующим развитием вос- |
врожденного иммунитета и 4.2. Элементы адаптивно- |
||
палительной реакции (смотри 5.2. Активация фагоци- |
го иммунитета). Среди T-лимфоцитов, в свою очередь, |
||
тов и эндотелия и 5.4. Презентация антигенов). |
выделяют субпопуляции: T-хелперы и T-киллеры. |
||
Дендритные клетки – это гетерогенная популяция |
Для дифференцирования популяций и субпопуля- |
||
клеток с характерной отростчатой формой, по всей |
ций лимфоцитов используют метод иммунофенотипи- |
||
видимости, миелоидного происхождения. Дендритные |
рования – выявляют наличие определенных мембран- |
||
|
Рисунок 3.3. Этапы селекции T- |
||
|
лимфоцитов в тимусе. Предше- |
||
|
ственники T-лимфоцитов мигри- |
||
|
руют из красного костного мозга |
||
|
в тимус. По мере миграции из |
||
|
кортикальной части тимуса к ме- |
||
|
дулярной происходит сначала по- |
||
|
зитивная, а затем негативная се- |
||
|
лекция. В результате позитивной |
||
|
селекции |
отбираются |
только |
|
клетки, |
способные эффективно |
|
|
взаимодействовать с антигенпре- |
||
|
зентирующими рецепторами. В |
||
|
ходе негативной селекции выжи- |
||
|
вают только лимфоциты, не спо- |
||
|
собные |
распознавать |
собствен- |
|
ные антигены индивидуума. На |
||
|
обоих этапах происходит апоптоз |
||
|
не прошедших селекцию клеток. В |
||
|
итоге селекцию проходят не бо- |
||
|
лее 5% T-лимфоцитов, которые с |
||
|
током крови мигрируют из тиму- |
||
|
са во вторичные органы иммун- |
||
|
ной системы. |
|
|
|
|
||
|
|
||
Зорников Д.Л., Литусов Н.В. Основы противоинфекционной иммунологии |
Страница 7 |
||

ных белков, называемых кластерами дифференциров-
ки (clusters of differentiation – CD). Для обозначения кластеров дифференцировки используют аббревиатуру CD и порядковый номер, например, CD4. Определенные CD появляются на мембране лимфоцитов в процессе их дифференцировки и обеспечивают различные биологические функции. Каждая популяция лимфоцитов имеет свой набор CD (таблица 3.1).
Таблица 3.1. Кластеры дифференцировки (CD) отдельных
популяций и субпопуляций лимфоцитов. Определение CD
молекул на мембранах позволяет дифференцировать популяции и субпопуляции лимфоцитов.
Зорников Д.Л., Литусов Н.В. Основы противоинфекционной иммунологии |
Страница 8 |