

2.Регуляція рухів;
3. Організація й реалізація рухових програм та інших поведіикових реакцій, зі значною питомою вагою гальмівних впливів (бя вважаються «гальмівним ситом» для кори мозку).
4. Організація контролю комплексних патерніа рухів, здебільшого поведінкових, із впливом на інтенсивність, напрямок, послідовність різноманітних наступних і паралельних рухів для досягнення
специфічних моторних наслідків, тобто певної мети.
5.забезпечують загальний руховий фон, збагачують головний рух допоміжними, співдружніми (розмахування руками під час ходи, бігу, стрибків; жестикуляція при розмові). Механізм та призначення полягає в підвищенні рівня активності робочих центрів реципрокним шляхом, що забезпечує високу ефективність головного рухового акту.
БЯ регулюють скорочення дрібних м'язових груп і рухи в дрібних суглобах, забезпечуючи цілеспрямованість, влучність рухових актів. Саме втручанням БЯ забезпечується емоційне забарвлення обличчя (тобто міміки) та інтонації голосу (мови).
Завдяки регуляції м'язового тоїгусу за пластичним типом (на його подолання потрібне безперервне зусилля) забезпечується злитність, плавність та економічність рухів.
Участь БЯ у здійсненні рухових програм стосується впливів на їх формування, а також здійснення; на ініціацію та завершення, особливо для повільних рухових компонентів; нарешті-на запам'ятовування. Заостанніми даними БЯ належить важлива роль у переході від задуму (фази підготовки) до обраної програми дії (фази виконання).
Синдром Паркінсона пов'зують головним чином з дефіцитом у постачанні дофаміну компактною частиною чорної речовини й отже порушенням у зв'язку з цим нігро-стріатних зв'язків. Саме недостатність дофаміну спричиняє надмірне збудження (розгальмовування) нейронів хвостатого ядра, отже й підсилений його гальмівний вплив на палеостріарні структури. Та використаїгаям для замінної терапії а-ДОФА (препарату дофаміну, здатного проникати через гемато-енцефалічний бар'єр) повністю проявів паркінсонізму не ліквідується. Мабуть крім дофаміну мають значення й інші медіатори - глутамат, ацетилхолін, пептиди тощо.
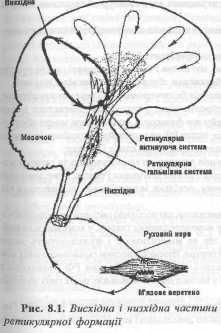
РЕТИКУЛЯРНА ФОРМАЦІЯ
Ретикулярна формація (РФ) (сітчастий утвір) являє собою комплекс структур мозку, що має велику протяжність. Починається від желатинної субстанції спинного мозку і закінчується неспецифічними ядрами таламусу.
Функціонально РФ поділяють на дві частини -низхідну (еферентну) і ввсхідну (аферентну) . Функції низхідного відділу ретикулярної формації
Низхідний відділ РФ представлений системою еферентних нейронів, аксони яких закінчуються си-напсами на нейронах
105
структур мозку, що розташовані ниж-че. В її складі є ядра, що забезпечують регуляцію рухів і м'язового тонусу.
Вперше вплив РФ на здійснення сгашальних рефлексів було доведено І.М.Сєченовим (1863) у досліді на «таламічній» жабі. Зануренням задньої кінцівки в розчин кислоти викликається захисний згинальний рефлекс і визначається його ЛП. Потім на таламус накладається кристал кухонної солі. Це призводить до значного збільшення ЛП рефлексу, або до його відсутності. Результатом досліду стало відкриття явища гальмування в ЦНС. У даному випадку виявлено гальмівний вплив РФ на рухові центри спинного мозку.
У середині XX століття групою американських вчених під керівництвом Мегуна було встановлено, що ретикулярні ядра гальмують спинальні рухові центри. Зокрема, було визначено пригнічення децеребраційної ригідності при подразненні ретикулярних ядер стовбура. Механізми можуть бути різними:
• безпосередній гальмівний вплив на а-мотонейрони спинного мозку;
• збудження гальмівних інтернейронів Реншоу.
Пізніше було встановлено, що вплив РФ на розташовані нижче центри може бути І збуджуючим. Механізми збуджуючих впливів слідуючі:
• збудження сс-мотонейронів безпосередньо або через інтернейрони;
• збудження сс-мотонейронів через у-петлю; РФ довгастого мозку збуджує здебільшого мотонейрони флексорів, тоді як ядра моста збуджують мотонейрони екстензорів; вплив РФ на мотонейрони шийних І грудних сегментів має суттєве значення для регуляції вдиху і видиху;
• збудження мотонейронів завдяки загальмовуванню зв'заних з мо-тонейронами гальмівних клітин Уілсона клітинами Реншоу; отже, кінцевий ефект виявляється збуджуючим, оскільки мотонейрони звільняються від гальмівних впливів.
В.МБєхтєрєвим було встановлено, що особливо сильний низхідний збуджуючий вплив виявляється у новонароджених тварин, у яких волокна нейронів пірамідного тракту ще не міелінізовані і функціонують лише філогенетично старіші рухові центри. Взаємовідносини гальмівного типу між центрами ще не сформувались. Ефект подразнення РФ виявляється генералізованими реакціями у вигляді досить повільних рухів типу тонічніх судом.
Окрім регуляції рухів, РФ впливає на ряд вегетативних функцій. Так, відомо, що до складу бульбарного дихального і кардіо-васкулярного центрів
входить значна кількість ретикулярних нейронів. Ретикулярні нейрони виявлено в складі слиновидільних ядер, а також ядер блукаючого нерва.
Таким чином, РФ приймає участь у здійсненні життєво важливих функцій - дихання й кровообігу, у регуляції діяльності системи травлення, обміну речовин, теплообміну та інших вегетативних функцій.
8.2. функції висхідного відділу ретикулярної формації
Висхідний відділ РФ є системою аферентних нейронів, які впливають на розташовані вище відділи мозку (рис. 8.2, див. додаток на стор- 207).
Вплив висхідного відділу РФ було вперше встановлено Г.Мегуном і Д.Моруцці (І 949 р.). Електричне подразнення ретикулярних ядер стовбура за допомогою хронічно вживлених у мозок електродів відтворювало у спля-чоїкішки генералізовані зміни електричної активності кори мозку, властиві для активації, пов'язаної з неспанням. Повільні хвилі в складі електроенцефалограми (ЕЕГ) замінювались на швидкі, асинхронні. Такі зміни електричної активності мозку співпадали з реакцією пробудження тварини. Це стало приводом для того, щоб назвати висхідний відділ РФ «висхідною активуючою системою».
Додатковий доказ активуючого висхідного впливу РФ стовбура було отримано ще раніше, у 1937 р. ГБремером. Він робив у кішок перерізки мозку на двох різних рівнях. По-перше - на інтерколікулярному, відокремлюючи головний мозок від стовбура ("сегуеаи ізоіе"). Це препарат "сидячого мозку". Оскільки ретикулярна формація стовбура відокремлювалась від півкуль, реакції пробудження не відбувалось, не зважаючи на те, що зорова і нюхова аферентація зберігалась. Другий варіант - перерізка між спинним і довгастим мозком ("епссрЬаіе ізоіе")- У такому разі ознаки неспання можна було відтворити завдяки збереженню зв'язків головного мозку зі стовбуровими структурами.
Певний час висхідний відділ РФ називали «центр-енцефалічною системою» (Пенфілд, Боньалле, Гасто та ін.), виходячи з того, що стан неспання є необхідною умовою для свідомості, уваги та Інших проявів психічної діяльності. Ллє згодом було обгрунтовано і тепер загально визнано, Що РФ, як й інші структури мозку, знаходиться під постійним контролем йори великих півкуль.
Пізніше в складі висхідної частини РФ (у таламусі) було виявлено ядра, які сприяють переходу до сну зі стану неспання (П.М.Сєрков).
Слід відзначити також, що жорсткий поділ РФ на висхідний і низхідний відділи досить умовний, тому що мультиполярні нейрони можуть спрямовувати свої гілочки до розташованих вище й нижче структур і бути таким чином одночасно в складі висхідної й низхідної систем.
Для виконання ретикулярною формацією функцій надзвичайно важливе постійне підтримання її нейронів на певному рівні активності, що забезпечується надходженням імпульсів по колатералях від провідникових шляхів і мозкових центрів. Особливо важливу роль у цьому відіграє система трійчастого нерва (Сєрков, Макулькін).
Ще однією суттєвою характеристикою ретикулярних нейронів є їх підвищена хемочутливість до різних гуморальних агентів - медіаторів та інших фізіологічно активних речовин атакож фармакологічних речовин. Усе це важливо для виконання РФ функцій, пов' язаних із підтриманням гомеостазу.
Отже, ядра РФ приймають участь у забезпеченні багатьох функцій, соматичних та вегетативних; у певній мірі - і в рузомовій діяльності теж.