

ЗООЛОГИЯ / 1-ые вопросы билета / Документ Microsoft Office Word
.docx31.
Нервная система книдарий и гребневиков
Наиболее примитивными животными, у которых есть нервная система, считаются книдарии. У полипов она представляет собой примитивную субэпителиальную нервную сеть (нервный плексус), оплетающую всё тело животного и состоящую из нейронов разного типа (чувствительных и ганглиозных клеток), соединённых друг с другом отростками (диффузная нервная система), особенно плотные их сплетения образуются на оральном и аборальном полюсах тела. Раздражение вызывает быстрое проведение возбуждения по телу гидры и приводит к сокращению всего тела, в связи с сокращением эпителиально-мускульных клеток эктодермы и одновременно их расслаблением в энтодерме. Медузы устроены сложнее полипов, в их нервной системе начинает обособляться центральный отдел. Помимо подкожного нервного сплетения у них имеются ганглии по краю зонтика, соединённые отростками нервных клеток в нервное кольцо, от которого иннервируются мышечные волокна паруса и ропалии — структуры, содержащие различные органы чувств (диффузно-узловая нервная система). Бо́льшая централизация наблюдается у сцифомедуз и особенно кубомедуз. Их 8 ганглиев, соответствующие 8 ропалиям, достигают достаточно крупных размеров.
Нервная система гребневиков включает субэпителиальное нервное сплетение со сгущениями вдоль рядов гребных пластинок, которые сходятся к основанию сложно устроенного аборального органа чувств. У некоторых гребневиков описаны находящиеся рядом с ним нервные ганглии.
Нервная система первичноротых
Плоские черви имеют уже подразделенную на центральный и периферический отделы нервную систему. В целом нервная система напоминает правильную решётку — такой тип строения был назван ортогоном. Она состоит из мозгового ганглия, у многих групп окружающего статоцист (эндонного мозга), который соединен с нервными стволами ортогона, идущими вдоль тела и соединенные кольцевыми поперечными перемычками (комиссурами). Нервные стволы состоят из нервных волокон, отходящих от рассеянных по их ходу нервных клеток. У некоторых групп нервная система довольно примитивна и близка к диффузной. Среди плоских червей наблюдаются следующие тенденции: упорядочивание подкожного сплетения с обособлением стволов и комиссур, увеличение размеров мозгового ганглия, который превращается в центральный аппарат управления, погружение нервной системы в толщу тела; и, наконец, уменьшение числа нервных стволов (у некоторых групп сохраняются лишь два брюшных (боковых) ствола).
У немертин центральная часть нервной системы представлена парой соединённых двойных ганглиев, расположенных над и под влагалищем хоботка, соединённых комиссурами и достигающих значительного размера. От ганглиев идут назад нервные стволы, обычно их пара и расположены они по бокам тела. Они также соединены комиссурами, расположены они в кожно-мускульном мешке или в паренхиме. От головного узла отходят многочисленные нервы, наиболее сильно развиты спинной нерв (часто двойной), брюшной и глоточный.
У брюхоресничных червей имеется надглоточный ганглий, окологлоточное нервное кольцо и два поверхностных боковых продольных ствола, соединённых комиссурами.
У нематод имеется окологлоточное нервное кольцо, вперёд и назад от которого отходят по 6 нервных стволов, наиболее крупные — брюшной и спинной стволы — тянутся вдоль соответствующих гиподермальных валиков. Между собой нервные стволы связаны полукольцевыми перемычками, иннервируют они соответственно мышцы брюшных и спинных боковых лент. Нервная система нематоды Caenorhabditis elegans была закартирована на клеточном уровне. Каждый нейрон был зарегистрирован, прослежено его происхождение и большинство, если не все, нейронные связи известны. У этого вида нервная система обладает половым диморфизмом: мужская и гермафродитная нервная система имеют разное количество нейронов и групп нейронов, чтобы выполнять полоспецифические функции.
У киноринх нервная система состоит из окологлоточного нервного кольца и вентрального (брюшного) ствола, на котором, в соответствии с присущей им сегментацией тела, группами расположены ганглионарные клетки.
Схоже устроена нервная система волосатиков и приапулид, но их вентральный нервный ствол лишен утолщений.
У коловраток имеется крупный надглоточный ганглий, от которого отходят нервы, особенно крупные — два нерва, идущие через всё тело по бокам кишечника. Более мелкие ганглии лежат в ноге (педальный ганглий) и рядом с жевательным желудком (ганглий мастакса).
У скребней нервная система очень проста: внутри влагалища хоботка имеется непарный ганглий, от которого отходят тонкие веточки вперёд к хоботку и два более толстых боковых ствола назад, они выходят из влагалища хоботка, пересекают полость тела, а затем по её стенкам идут назад.
У кольчатых червей имеется парный надглоточный нервный узел, окологлоточными коннективами (коннективы в отличие от комиссур соединяют разноимённые ганглии) соединённый с брюшной частью нервной системы. У примитивных полихет она состоит из двух продольных нервных тяжей, в которых располагаются нервные клетки. У более высокоорганизованных форм они образуют парные ганглии в каждом сегменте тела (нервная лестница), а нервные стволы сближаются. У большинства же полихет парные ганглии сливаются (брюшная нервная цепочка), у части сливаются и их коннективы. От ганглиев отходят многочисленные нервы к органам своего сегмента. В ряду полихет происходит погружение нервной системы из-под эпителия в толщу мышц или даже под кожно-мускульный мешок. Ганглии разных сегментов могут концентрироваться, если сливаются их сегменты. Аналогичные тенденции наблюдаются и у олигохет. У пиявок нервная цепочка, лежащая в брюшном лакунарном канале, состоит из 20 или более ганглиев, причём в один объединяются первые 4 ганглия (подглоточный нервный узел) и последние 7.
У эхиурид нервная система развита слабо — окологлоточное нервное кольцо соединено с брюшным стволом, но нервные клетки рассеяны по ним равномерно и нигде не образуют узлов.
У сипункулид имеется надглоточный нервный ганглий, окологлоточное нервное кольцо и лишённый нервных узлов брюшной ствол, лежащий на внутренней стороне полости тела.
Тихоходки имеют надглоточный ганглий, окологлоточные коннективы и брюшную цепочку с 5 парными ганглиями.
Онихофоры имеют примитивную нервную систему. Мозг состоит из трёх отделов: протоцеребрум иннервирует глаза, дейтоцеребрум — антенны, а тритоцеребрум — переднюю кишку. От окологлоточных коннектив отходят нервы к челюстям и ротовым сосочкам, а сами коннективы переходят в далёкие друг от друга брюшные стволы, равномерно покрытые нервными клетками и соединённые тонкими комиссурами.
Нервная система членистоногих
У членистоногих нервная система слагается из парного надглоточного узла, состоящего из нескольких соединённых нервных узлов (головной мозг), окологлоточных коннектив и брюшной нервной цепочки, состоящей из двух параллельных стволов. У большинства групп головной мозг делится на три отдела — прото-, дейто- и тритоцеребрум. Каждый сегмент тела имеет по паре нервных ганглиев, но часто наблюдается слияние ганглиев с образованием крупных нервных центров; например, подглоточный нервный узел состоит из нескольких пар сросшихся ганглиев — он контролирует слюнные железы и некоторые мышцы пищевода.
В ряду ракообразных в целом наблюдаются те же тенденции, что и у кольчатых червей: сближение пары брюшных нервных стволов, слияние парных узлов одного сегмента тела (то есть образование брюшной нервной цепочки), слияние её узлов в продольном направлении по мере объединения сегментов тела. Так, у крабов имеется лишь две нервные массы — головной мозг и нервная масса в груди, а у веслоногих и ракушковых раков образуется единственное компактное образование, пронизанное каналом пищеварительной системы. Головной мозг раков состоит из парных долей — протоцеребрума, от которого отходят зрительные нервы, имеющие ганглиозные скопления нервных клеток, и дейтоцеребрума, иннервирующего антенны I. Обычно добавляется и тритоцеребрум, образованный слившимися узлами сегмента антенн II, нервы к которым обычно отходят от окологлоточных коннективов. У ракообразных имеется развитая симпатическая нервная система, состоящая из мозгового отдела и непарного симпатического нерва, имеющего несколько ганглиев и иннервирующего кишечник. Важную роль в физиологии раков играют нейросекреторные клетки, расположенные в различных частях нервной системы и выделяющие нейрогормоны.
Головной мозг многоножек имеет сложное строение, образован, скорее всего, многими ганглиями. Подглоточный ганглий иннервирует все ротовые конечности, от него начинается длинный парный продольный нервный ствол, на котором в каждом сегменте приходится по одному парному ганглию (у двупарноногих многоножек в каждом сегменте, начиная с пятого, по две пары ганглиев, расположенных одна за другой).
Нервная система насекомых, также состоящая из головного мозга и брюшной нервной цепочке, может достигать значительного развития и специализации отдельных элементов. Головной мозг состоит из трёх типичных отделов, каждый из которых состоит из нескольких ганглиев, разделённых прослойками нервных волокон. Важным ассоциативным центром являются «грибовидные тела» протоцеребрума. Особенно развитый мозг у общественных насекомых (муравьёв, пчёл, термитов). Брюшная нервная цепочка состоит из подглоточного нервного узла, иннервирующего ротовые конечности, трёх крупных грудных узлов и брюшных узлов (не более 11). У большинства видов не встречается во взрослом состоянии более 8 ганглиев, у многих и они сливаются, давая крупные ганглиозные массы. Может доходить до образования только одной ганглиозной массы в груди, иннервирующей и грудь, и брюшко насекомого (например, у некоторых мух). В онтогенезе зачастую происходит объединение ганглиев. От головного мозга отходят симпатические нервы. Практически во всех отделах нервной системы имеются нейросекреторные клетки.
У мечехвостов головной мозг внешне не расчленён, но имеет сложное гистологическое строение. Утолщённые окологлоточные коннективы иннервируют хелицеры, все конечности головогруди и жаберные крышки. Брюшная нервная цепочка состоит из 6 ганглиев, задний образован слиянием нескольких. Нервы брюшных конечностей соединены продольными боковыми стволами.
Нервная система паукообразных имеет чёткую тенденцию к концентрации. Головной мозг состоит только из протоцеребрума и тритоцеребрума в связи с отсутствием структур, которые иннервирует дейтоцеребрум. Метамерность брюшной нервной цепочки яснее всего сохраняется у скорпионов — у них большая ганглиозная масса в груди и 7 ганглиев в брюшке, у сольпуг их только 1, а у пауков все ганглии слились в головогрудную нервную массу; у сенокосцев и клещей нет разграничения между нею и головным мозгом.
Морские пауки, как и все хелицеровые, не имеют дейтоцеребрума. Брюшная нервная цепочка у разных видов содержит от 4-5 ганглиев до одной сплошной ганглиозной массы.
Нервная система моллюсков
У примитивных моллюсков хитонов нервная система состоит из окологлоточного кольца (иннервирует голову) и 4 продольных стволов — двух педальных (иннервируют ногу, которые связаны без особого порядка многочисленными комиссурами, и двух плевровисцеральных, которые расположены кнаружи и выше педальных (иннервируют внутренностный мешок, над порошицей соединяются). Педальный и плевровисцеральный стволы одной стороны также связаны множеством перемычек.
Схоже устроена нервная система моноплакофор, но педальные стволы соединяются у них только одной перемычкой.
У более развитых форм образуется в результате концентрации нервных клеток несколько пар ганглиев, которые смещаются к переднему концу тела, причём наибольшее развитие получает надглоточный узел (головной мозг).
Нервная система иглокожих примитивная, состоит из трёх отдельных частей, построенных по радиальному плану: нервное кольцо и радиальные нервные тяжи. В оральной стенке тела лежит две нервные системы — чувствительная и двигательная. В аборальной стенке тела — только двигательная. Органы чувств иглокожих достаточно разнообразны, но примитивны по строению. Они диффузно распределены по телу в виде различных чувствительных клеток (функции осязания, химического чувства, зрения). Светочувствительные клетки могут быть собраны в глазках. У морских звёзд глазки расположены на концах лучей, а у морских ежей — вокруг анального отверстия.
Сенсиллы (от лат. sensilis — чувствующий) — простейшие кожные органы чувств у беспозвоночных, выполняющие роль органов осязания, вкуса, обоняния. Все сенсиллы возникли из одного первоначального типа, путем специализации для восприятия определённого раздражения. Сенсилла представляет собой видоизмененный участок покровов тела (волосок, ямка, пластинка и т. п.), к которому подходят отростки одного или нескольких чувствительных нейронов. Наружная часть образована клетками покровов и представлена волоском (осязательные сенсиллы) либо тонкой кутикулярной пластинкой, часто в форме купола или конуса, погруженной в специальную ямку (обонятельные и вкусовые сенсиллы).
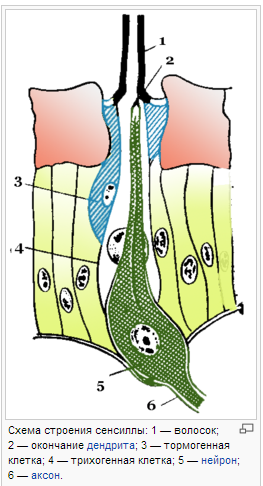
32.
Даже простейшие беспозвоночные животные обладают способностью к фототропизму благодаря своему, пусть крайне несовершенному, зрению.
У беспозвоночных встречаются очень разнообразные по типу строения и зрительным возможностям глаза и глазки — одноклеточные и многоклеточные, прямые и обращенные (инвертированные), паренхимные и эпителиальные, простые и сложные.
У членистоногих часто присутствует несколько простых глаз (иногда непарный простой глазок — например, науплиальный глаз ракообразных) или пара сложных фасеточных глаз. Среди членистоногих некоторые виды имеют и простые, и сложные глаза: так, у ос два сложных глаза и три простых глаза (глазка). У скорпионов 3-6 пар глаз (1 пара — главные, или медиальные, остальные — боковые), у щитня — 3. В эволюции фасеточные глаза произошли путем слияния простых глазков. Близкие по строению к простому глазу глаза мечехвостов и скорпионов, видимо, возникли из сложных глаз трилобитообразных предков путем слияния их элементов.
33.
У беспозвоночных органы равновесия. представлены статоцистами, или слуховыми пузырьками, имеющими различное строение и местоположение. У большинства беспозвоночных — это впячивания эктодермы, которые сообщаются с наружной средой при помощи канала или отшнуровываются, образуя замкнутый пузырёк. Внутри статоцистов расположены особые твёрдые образования — Статолиты. Внутренняя полость статоциста, как правило, выстлана чувствующими клетками, снабженными ресничками. Обычно статолит имеет большую плотность, чем окружающая его жидкость, и поддерживается чувствительными волосками. Если статолит окружен чувствительными волосками со всех сторон, то при любом изменении положения тела животного в пространстве будут раздражаться смещенным статолитом соответствующие группы волосков. Волосковые клетки ракообразных представляют собой первичные чувствующие Нейроны. Статоцисты медуз и морских ежей — маленькие колбовидные выпячивания наружных покровов тела, внутри которых также находятся статолиты. Но в этом случае реснитчатые клетки расположены снаружи статоциста среди эпителиальных клеток, окружающих его, либо в наружной стенке самого статоциста. У сцифоидных кишечнополостных имеется 8 статоцистов, расположенных радиально по краю мантии. У насекомых не обнаружено настоящих статоцистов. У некоторых водяных клопов и гладыша роль статоцистов выполняют покрытые чувствительными волосками наружные участки тела, удерживающие воздушные пузырьки (т. н. газовый статолит). Наиболее сложно устроены Р. о. головоногих моллюсков: статоцисты в виде пузырьков помещаются у них в капсуле головного хряща; однако даже у осьминога их удаление вызывает лишь незначительные нарушения в способности к ориентации. Возбуждение чувствующих клеток статоцистов передаётся в центральные отделы нервной системы. Механизмы ответных реакций животных, лишённых нервной системы, менее ясны. Многие Рецепторы равновесия дают сигналы двух типов — статические, связанные с положением тела, и динамические, связанные с ускорением.
34.
Первые животные появились более 3.3 млрд. лет назад в архейский период архерозойской эры. Такие примитивные животные продолжают существовать и в наше время и их объединяют в тип простейшие. Несмотря на примитивность, они за миллиарды лет своего существования очень хорошо приспособились к разным условиям обитания, и это позволяет им сосуществовать наравне с более высокоорганизованными животными. Простейшие (корненошки, жгутиковые, инфузории и споровики) повсеместно распространены и их насчитывается более 30 тыс. видов.
Роль органов у простейших выполняют клеточные органеллы, например, жгутики и реснички - органы движения, сократительные вакуоли - органы выделения и т.п. Органеллы могут быть высоко специализированы, как, например, трихоцисты некоторых инфузорий - короткие палочки в наружном слое клеточной цитоплазмы, которые при раздражении животного выстреливаются наружу, превращаясь в длинную упругую нить, вонзаются в тело врага или добычи и вносят в него ядовитое вещество. Базальные тельца ресничек инфузорий связаны между собой волоконцами, которые обеспечивают согласованную работу ресничек (которых может быть 10 - 15 тыс.), - зачаток нервной системы, а у неко-торых инфузорий есть сократительные актин-миозиновые волоконца - зачаток мышечной системы движения. Демонстрируя высокую степень специализации различных клеточных органелл, инфузории являются пределом сложности строения одноклеточного организма. Последующее повышение сложности организации могло быть связано только с появленим многоклеточности.
Многоклеточный организм отличается от колонии клеток тем, что клетки, по крайній мере, частично утрачивают самостоятельность и начинают специалиироваться для выполнения разных функций. Простейшие многоклеточные животные двуслойные: имеют слой внутренних жгутиковых клеток для пищеварения (энтодерму) и слой наружных защитных клеток (экзодерму). Общепризнано, что многоклеточные животные произошли от колонии жгутиковых клеток, однако как произошло такое превращение, остается спорным вопросом.
Существует несколько гипотез. Гастрейная гипотеза Геккеля предполагает, что стенка шарообразной колонии втянулась внутрь, и возник двуслойный организм с первичным ртом, ведушим в первичную пищеварительную полость. Такое преобразование (бластулы в гаструлу) действительно происходит в онтогенетическом развитии всех многоклеточных животных. Однако эта гипотеза не отвечает на вопрос, почему в эволюции произошло такое впячивание. По гипотизе Бючли предком мнооклеточных была не шаро- видная колония, а колония в форме двуслойной пластинки: нижний слой клеток – для питания, а верхний - для защиты (такие организмы есть и сейчас - это трихоплакс).
Пластинка свернулась, и образовалась пищеварительная полость. Наконец, в соответствии с гипотезой Мечникова питание первых многоклеточных и колониальных жгутиковых сходно: насытившиеся отдельные клетки погружались вглубь колонии и там переварювали пищу. Со временем это разделение на внешнее и внутреннее стало постоянным. Но в любом случае эволюционное развитие первых многоклеточных животных, вероятно, происходило так, как показано на рис. 196.
Губки - одни из самых примитивных многоклеточных организмов. Они имеют п’ять типов специализированных клеток (рис. 197), но взаимозависимость между клетками тела еще очень мала. Об этом свидетельствует тот факт, что если губку протереть через сито, то образовавшаяся кашица, состоящая из отдельных клеток и групп клеток, оказывается способной к восстановлению целого организма. У губок сохраняется самое примитивное внутриклеточное пищеварение, и они способны как к половому, так и вегетативному (почкованием) размножению.
Кишечнополостные - это тоже двуслойные животные, но число специализированных клеточных элементов у них возрастает до 6 - 7 (рис. 198), и в дополнение к внутриклеточному пищеварению добавляется пищеварение в пищеварительной полости (кишке). У кишечнополостных появляется диффузная нервная система и первые органы чувств - простые глаза, служащие только для различения света и тьмы, и статоцисты (органы равновесия). Они становятся способны к движению благодаря эпителиально-мышечным клеткам с развитой системой актин-миозиновых волокон. Так же, как губки, кишечнополостные способны как к половому, так и к вегетативному размножению. Кишечнополостные животные имеют радиальную симметрию из-за прикрепленного или малоподвижного образа жизни, когда пища может появиться с любой стороны.
Переход к активному движению сопровождался появлением переднего конца тела, которым животное наталкивается на препятствия и на добычу. На этом конце начинают скапливаться нервные клетки и органы чувств, он становится головным, возникает двусторонняя (билатеральная) симметрия. Такая симметрия тела возникает у плоских червей, перешедших от плавания к ползанию по дну. Но у них еще сохраняется черты, оставшиеся от радиальносимметричных кишечнополостных предков: например, рот у планарии расположен все еще в середине живота, а кишечник не имеет анального отверстия и разветвлен как на правую и левую стороны тела, так и по радиусам от глотки во все стороны. Однако строение ресничных плоских червей отличается значительно более высокой организацией по сравнению с кишечнополостными животными (рис. 199).
Плоские черви, как и все эволюционно следующие за ними животные, трехслойны: в процессе онтогенеза у них формируется не два, как у губок и кишечнополостных, а три зародышевых листка. Между эктодермой, образующей покровы, и энтодермой, из которой построен кишечник, у них еще имеется промежуточный зародышевый листок - мезодерма. Важная особенность строения плоских червей - это наличие у них кожно-мышечного мешка. Так называется совокупность эпителия и расположенной непосредственно под ним сложной системы дифференциро- ванных мышечных волокон. Впервые в эволюции у плоских червей возникает выделительная система. Пламенные клетки, имеющие внутри бьющиеся реснички, создают ток воды, который выносит собираемые отходы метаболизма в собирательные трубочки, ведущие к мельчайшим порам на поверхности животного (рис. 199). Нервная система состоит из парного мозгового ганглия и идущих от него назад нервных стволов, соеди- ненных кольцевыми перемычками. Тело плоских червей может быть достаточно протяженным, но сплющено в спинно-брюшном направлении, так как дыхательной и кровеносной систем у них нет, и кислород поступает в тело червя путем диффузии через всю поверхность тела. Плоские черви гермафродиты, но некоторые из них способны к бесполому размножению посредством поперечного деления перетяжкой.
Тело плоских червей не имеет полости. Полость тела, отличающаяся от пищеварительной полости тем, что не имеет сообщения с наружней средой появилась впервые у круглых червей. Полость тела заполнена жидкостью и выполняет несколько функций:
1) несжимаемая жидкость придает упругость телу червя (гидроскелет);
2) через полость тела происходит транспорт веществ, усвоенных из пищи, от кишечника к мускулатуре и половой системе;
3) через полость тела осуществляется вынос продуктов обмена к органам выделения, которыми служат специальные кожные железы. Таким образом, полость тела играет роль внутренней среды организма. Кроме того, у этого типа червей появилась задняя кишка и анальное отверстие, кишечник стал сквозным (рис. 200). Снаружи круглые черви одеты сложно устроенной многослойной кутикулой, которая вместе с гидроскелетом создает опору для мускулатуры. Из-за жесткой кутикулы рост сопровождается линькой. В отличие от плоских червей они раздельнополы и неспособны к регенерации.
35.
Бесполое и половое размножение
Бесполое размножение. В животном мире существует целый ряд способов размножения, например, прямое деление или почкование, характерные для низших беспозвоночных, а также партеногенез, отмечаемый даже у высших позвоночных. Совершенно очевидно, что именно бесполое размножение представляет собой наиболее простой и наименее энергоемкий путь увеличения численности особей. Однако, почему-то, в процессе эволюции возник сложный, сопряженный с множеством проблем и условностей, процесс полового размножения.
Половое размножение.
1.Конъюгация. Половое размножение появляется у животных уже на самых низших ступенях эволюционной лестницы. Так, уже у простейших одноклеточных микроорганизмов - инфузорий, размножающихся прямым делением, наблюдается так называемая конъюгация, представляющая собой некий аналог полового процесса. В процессе конъюгации две инфузории как бы срастаются, на время, в течение которого обмениваются наследственной информацией. Затем инфузории разъединяются, и затем каждая продолжает делиться сама по себе.
2. Партеногенез. У целого ряда беспозвоночных, а также многих позвоночных животных отмечается такое явление, как партеногенез, при котором самки откладывают яйца или рожают живых детенышей без участия самцов. Вылупляются из этих яиц или рождаются подобным образом только самки, причем интересно, что в природе существуют целые локальные популяции подобных видов. Подобная популяция скальных ящериц была обнаружена в Армении. В остальных местах своего обитания этот, весьма обычный вид, размножается обычным образом.
Партеногенез в экспериментальных условиях возможен даже у млекопитающих. Для этого необходимо каким-либо образом стимулировать к делению неоплодотворенную яйцеклетку, что осуществимо разными путями, например, просто нанесением укола микроиглой.
3. Гермафродитизм. У целого ряда беспозвоночных животных имеет место гермафродитизм, при котором у каждой особи имеются как мужские, так и женские половые железы. Среди наиболее известных животных гермафродитами являются, например, дождевые черви, пиявки и многие виды улиток. Однако, несмотря на то, что у подобных животных каждая особь производит и яйцеклетки, и сперматозоиды, они спариваются друг с другом, производя взаимный обмен половыми клетками. У крупных, не имеющих раковин, моллюсков аплизий или морских зайцев, обитающих в прибрежной зоне моря, в процессе оплодотворения может одновременно принимать участие до 10-12 особей, играя сразу как роли самцов, так и роли самок.
Однако, несмотря на наличие у животных-гермафродитов половых клеток обоих типов, они крайне редко прибегают к самооплодотворению, и напротив, обычно стремятся к спариванию с другими, предпочтительно не родственными особями.
4. Смена пола. У некоторых животных, в том числе у целого ряда видов рыб, с возрастом происходит смена пола. Так, у хорошо известных аквариумистам рыбок-меченосцев довольно обычным является превращение немолодых самок в самцов. При этом у них вырастает мечевидный отросток на хвосте, имеющийся только у самцов, и они начинают проявлять типичное половое поведение, успешно оплодотворяя самок. Аналогичное явление наблюдается и у небольших рыбок морских юнкеров, обитающих в прибрежной зоне Черного моря. У некоторых видов рыб отмечен и обратный процесс: превращение самцов в самку.