

Моногибридное скрещивание
Опыты Мендель проводил на горохе. При скрещивании сортов гороха, имеющих желтые и зеленые семена (скрещивались гомозиготные организмы или чистые линии), все потомство (т.е. гибриды первого поколения) оказалось с желтыми семенами. Противоположный признак (зеленые семена) как бы исчезает. Обнаруженная закономерность получила название правило единообразия (доминирования) гибридов первого поколения (или первый закон Г.Менделя).
Опыты по скрещиванию записывают в виде схем:
А – ген желтой окраски
а – ген зеленой окраски
Р - (parents – родители)
F - (filii – дети)
Р ♀АА х ♂аа
ж з
G (А) (а)
F1 Аа – 100% желтые
Итак, все гибриды первого поколения оказываются однородными: гетерозиготными по генотипу и доминантными по фенотипу.
Таким образом, первое правило (закон) Менделя единообразия гибридов первого поколения можно сформулировать следующим образом: при скрещивании гомозиготных особей, отличающихся друг от друга по одной паре альтернативных признаков, все потомство в первом поколении единообразно как по фенотипу, так и по генотипу
.
Правило расщепления. Второй закон Менделя
Если скрестить гибриды первого поколения между собой, во втором поколении появляются особи, как с доминантными, так и с рецессивными признаками, т.е. возникает расщепление в определенном численном соотношении. В опытах с горохом желтых семян оказывается в три раза больше, чем зеленых. Эта закономерность получила название второго закона (правило) Менделя, или закона (правило) расщепления.
Р ♀ Аа х ♂ Аа
ж ж
G (А) (а) (А) (а)
F2 АА; Аа, Аа; аа
желтые зеленые
Расщепление по фенотипу 3:1, по генотипу 1АА:2Аа:1аа
Второй закон (правило) Менделя: при скрещивании двух гетерозиготных особей, анализируемых по одной альтернативной паре признаков (т.е. гибридов), в потомстве ожидается расщепление по фенотипу 3:1 и по генотипу 1:2:1.
Ди- и полигибридное скрещивание. Третий закон Менделя
При дигибридном скрещивании родительские организмы анализируются по двум парам альтернативных признаков. Мендель изучал такие признаки как окраску семян и их форму. При скрещивании гороха с желтыми и гладкими семенами с горохом, имеющим зеленые и морщинистые семена, в первом поколении все потомство оказалось однородным, проявились только доминантные признаки – желтый цвет и гладкая форма. Следовательно, как и при моногибридном скрещивании здесь имело место правило единообразия гибридов первого поколения или правило доминирования.
А – ген желтого цвета
а – ген зеленого цвета
В – ген гладкой формы
в – ген морщинистой формы
Р ♀ААВВ х ♂аавв
ж. гл. з. морщ.
G (АВ) (ав)
F1 АаВв – желтые гладкие
При скрещивании гибридов первого поколения между собой произошло расщепление по фенотипу:
Р ♀ АаВв х ♂АаВв
|
|
АВ |
Ав |
аВ |
ав |
|
АВ |
ААВВ-ж.гл |
ААВв-ж.гл |
АаВВ-ж.гл |
АаВв-ж.гл. |
|
Ав |
ААВв-ж.гл |
ААвв-ж.м. |
АаВв-ж.гл |
Аавв-ж.м |
|
аВ |
АаВВ-ж.гл |
АаВв-ж.гл |
ааВВ-з.гл. |
ааВв-з.гл |
|
ав |
АаВв-ж.гл |
Аавв-ж.м |
ааВв-з.гл |
аавв-з.м |
9 частей – желтых гладких
3 части – желтых морщинистых
3 части – зеленых гладких
1 часть – зеленых морщинистых
Из этого скрещивания видно, что во втором поколении имелись не только особи с сочетанием признаков родителей, но и особи с новыми комбинациями признаков.
Кроме того, Мендель обнаружил, что каждая пара признаков (цвет и форма) дала расщепление приблизительно в отношении 3:1, то есть как при моногибридном скрещивании. Отсюда был сделан вывод, что каждая пара альтернативных признаков при ди- и полигибридном скрещивании наследуется независимо друг от друга.
Третье правило или третий закон Менделя формулируется следующим образом: при скрещивании гомозиготных особей отличающихся двумя (или более) парами альтернативных признаков, во втором поколении наблюдается независимое наследование и комбинирование признаков, если гены, определяющие их, расположены в различных гомологичных хромосомах.
Кроме законов, Мендель сформулировал две гипотезы: факторальную и гипотезу «чистоты гамет», с помощью которых он попытался объяснить установленные закономерности.
Факторальная гипотеза указывает на то, что в клетках содержится фактор (ген), который и несет признак. Родители передают потомкам не признаки, а эти факторы.
Гипотеза «чистоты гамет»: организм по каждому признаку несет два наследственных фактора (один от отца, второй от матери). Эти наследственные факторы, находясь в клетках, не сливаются друг с другом и при формировании гамет расходятся в разные гаметы.
Анализирующее скрещивание
Рецессивный аллель проявляется только в гомозиготном состоянии. Поэтому о генотипе организма проявляющего рецессивный признак можно судить по фенотипу.
Гомозиготная и гетерозиготная особи, проявляющие доминантные признаки по фенотипу неотличимы. Для определения генотипа производят анализирующее скрещивание и узнают генотип родителей по потомству.
Анализирующее скрещивание заключается в том, что особь, генотип которой не ясен, но должен быть выяснен скрещивается с рецессивной формой. Если от такого скрещивания все потомство окажется однородным, значит анализируемая особь гомозиготна, если же произойдет расщепление, то она гетерозиготна
Р ♀ АА х ♂ аа
G (А) (а)
F1 Аа
Р ♀ Аа х ♂ аа
G (А) (а) (а)
F2 Аа; аа
1:1
Как видно из схемы, при анализирующем скрещивании для потомства гетерозиготной особи характерно расщепление 1:1.
Лекция 10
Хромосомная теория наследственности
План
1.Пол как наследственный признак.
2.Определение пола.
3.Наследование признаков, сцепленных с полом и ограниченных полом.
4.Сцепление генов. Опыты и правило Моргана.
5.Основные положения хромосомной теории наследственности.
Хромосомная теория наследственности - раздел генетики о локализации наследуемых факторов в хромосомах клеток.
Суть ее заключается в том, что все признаки и свойства живых организмов определяются генами, расположенными в хромосомах клетки в линейном порядке. Основателем ее явился американский генетик Томас Морган, теория разработана его сотрудниками в начале ХХ века.
Пол как наследственный признак
Одним из признаков у многих живых организмов является пол (мужской и женский).
Пол - это совокупность морфологических, физиологических, биохимических и поведенческих признаков организма, которые обуславливают репродукцию (воспроизведение).
Пол - это генетически обусловленный признак, т.е. пол зародыша определяется генетическими механизмами - комбинацией половых хромосом. Половые хромосомы определяют первичные половые признаки.
Под их влиянием формируются половые железы (семенники и яичники), выделяющие мужские и женские половые гормоны. Половые гормоны определяют развитие вторичных половых признаков того или другого пола и проявляются у организмов в период полового созревания (у человека в 12 - 15 лет). Вторичные половые признаки - различие в размерах и пропорциях тела, тембр голоса, степень развития волосяного покрова и др.
Определение пола
У большинства организмов пол определяется в момент оплодотворения (сингамно) и регулируется хромосомным набором зиготы, его называют хромосомный тип определения пола.
У человека и млекопитающих женские особи имеют в кариотипе две Х - хромосомы (ХХ), а мужские Х- и У- хромосомы. Женские особи являются гомогаметными, поскольку формируют лишь один тип гамет, а мужские - гетерогаметными, т.к. образуют два типа гамет.
У птиц и бабочек гетерогаметным полом является женский пол, а гомогаметными - мужской. У них половые хромосомы обозначают как Z и W.
У некоторых клопов, жуков самки имеют состав половых хромосом ХХ, самцы же несут всего одну из них - (ХО).
Совершенно особый тип определения пола имеется у перепончатокрылых, в частности, у пчел. У этих насекомых самки развиваются из оплодотворенных яиц и клетки их тела диплоидны, а самцы развиваются партеногенетически из неоплодотворенных яиц и имеют гаплоидные клетки.
Хромосомная теория наследования пола утверждает, что гены определяющие развитие пола, локализованы в половых хромосомах. У человека гены, обуславливающие развитие женского пола, находятся в Х - хромосоме, а гены мужского пола - в У - хромосоме.
Основные типы хромосомного определения пола
Иначе обстоит дело у дрозофилы. При изучении наследования пола у мухи - дрозофилы было установлено, что у самки две одинаковые Х - хромосомы, а у самца Х- и У- - хромосомы. У дрозофилы гены, определяющие женский пол, находятся в Х - хромосоме, а У - хромосома генетически инертна и гены, определяющие развитие мужского пола, находятся в аутосомах. Оплодотворенные яйцеклетки, имеющие диплоидный набор хромосом и две Х - хромосомы дают начало самкам (2А + ХХ), а имеющие диплоидный набор аутосом и одну Х - хромосому дают самцов(2А+ХУ).
Опыты подтвердили, что чем больше наборов аутосом, тем больше были выражены признаки мужского пола. Из этого был сделан вывод, что У - хромосома у мухи - дрозофилы не имеет существенного значения для определения мужского пола.
Кроме сингамного способа определения пола существует эпигамный способ дифференцировки пола, который определяется воздействием условий окружающей среды, например, у морского кольчатого червя Bonnelia определяется условиями развития яйцеклеток (прогамный способ, как у коловраток).
Наследование признаков, сцепленных с полом и ограниченных полом
Сцепленными с полом называют признаки, развитие которых обусловлено генами, расположенными в половых хромосомах.
Если ген находится в У-хромосоме, то он наследуется у человека, млекопитающих - от отца к сыну, а у птиц - от матери к дочери.
Если ген располагается в Х-хромосоме, наследование будет более сложным. Впервые особенности наследования генов, сцепленных с Х-хромосомой были открыты в опытах на дрозофиле при изучении наследования окраски глаз. Красная окраска глаз доминирует над белой.
При скрещивании гомозиготной красноглазой самки с белоглазым самцом в F1 все потомство красноглазое.
Р ♀ ХАХА х ♂ХªУ
кр. бел.
G (XА) (Xª) (У)
F1 XАXª х XАУ
кр. кр.
G (ХА) (Xª) (XА) (У)
F2 ХАХА , ХАХª, ХАУ, ХªУ
кр. кр. кр. бел.
При скрещивании гибридов первого поколения друг с другом в F2 происходит расщепление по фенотипу: 3 красноглазые особи: 1 белоглазая, и белоглазыми бывают только самцы.
При реципрокном (обратном) скрещивании, когда самка гомозиготная по гену белоглазости, скрещивается с красноглазым самцом, расщепление в F1 по фенотипу 1:1. При этом белоглазыми будут только самцы. При скрещивании мух F1 между собой в F2 будут мухи с обоими признаками в равном отношении 1:1, как среди самок, так и среди самцов.
Р ♀ ХªХª х ♂ХА У
бел. кр.
G (Хª) (ХА) (У)
F1 ХАХª х ХªУ
кр. бел.
G (ХА) (Хª) (Хª) (У)
F2 ХАХª, ХªХª, ХАУ, ХªУ
кр. бел. кр. бел.
Из результатов опыта Моргана можно заключить, что гены окраски глаз расположены только в половой Х-хромосоме. Результаты скрещивания зависят от того, какой пол несет доминантный признак.
Наследование, сцепленное с полом, необходимо отличать от наследования, ограниченного полом.
Развитие признаков (ограниченных полом) обусловлено генами, расположенными в аутосомах (а не в половых хромосомах), но на проявление признаков в фенотипе сильно влияет пол, т.е. развитие признака зависит от влияния половых гормонов.
Например, облысение со лба, типично для мужчин. У мужчин под действием мужских половых гормонов (тестостерона) ген ведет себя как доминантный. Еще пример, тембр голоса - баритон и бас - характерны только для мужчин.
Сцепление генов. Опыты и правило Моргана
Изучение сцепленного с полом наследования стимулировало изучение сцепления между генами, находящимися в аутосомах.
Для любого организма характерно видовое постоянство хромосом в кариотипе. Генов, определяющих признаки, у организмов намного больше, чем хромосом. Например, у мухи дрозофилы 8 хромосом в соматических клетках, а генов около 1000. Значит, в каждой хромосоме находится много генов.
Гены, локализованные в одной хромосоме, образуют группы сцепления.
Число групп сцепления равно гаплоидному числу хромосом.
Наследование генов, локализованных в одной хромосоме, называется сцепленным наследованием.
В начале ХХ века Т. Морган и его сотрудники описали явление сцепления генов - совместную передачу групп генов из поколения в поколение.
Опыты проводились на мухах дрозофилах с учетом двух пар альтернативных признаков:
Цвет тела – серый (В) и черный (в)
Длина крыльев – нормальные (V) и короткие (v)
У мухи - дрозофилы окраски тела и длины крыльев находятся в одной паре гомологичных хромосом, т.е. относятся к одной группе сцепления, что и было доказано в опытах. Запись генотипов при сцеплении видоизменяется: генотип записывается BV
═
bv
(две черточки означают, что организм диплоидный).
При скрещивании мух, имеющих серый цвет тела и нормальные крылья с мухами черного цвета и короткими крыльями в F1 все мухи имели серый цвет тела и нормальные крылья.
Запись схемы скрещивания:
P ♀BV ♂bv
═ x ═
BV bv
сер.нор. чер.кор.
G (BV) (bv)
BV
F1 ═
bv
Провели анализирующее скрещивание. В первом случае скрестили гибридного самца с серым телом и нормальными крыльями
с черной самкой, имеющей короткие крылья, и в результате скрещивания получили 2 типа потомков, похожих на родителей в соотношении 1:1.
Ген серого цвета тела и ген нормального строения крыльев передаются вместе, а ген черного цвета тела и ген коротких крыльев тоже вместе. Этот опыт демонстрирует полное сцепление. Причина его заключается в том, что гены, обуславливающие два различных признака, лежат в одной хромосоме. Это видно на схеме полного сцепления:
Р ♀bv ♂BV
═ x ═
bv bv
G ( bv) ( BV ) (bv)
BV bv
F1 ═ ═
bv bv
1:1
Другой результат получается, если для анализирующего скрещивания брать гибридную самку с серым телом и нормальными крыльями и рецессивного по обоим признакам самца.
В этом случае появилось четыре типа потомков в соотношении:
серых длиннокрылых - 41, 5 %; серых короткокрылых - 8, 5 %,
черных длиннокрылых - 8, 5% , черных короткокрылых - 41, 5 %.
В этом случае имеет место неполное сцепление:
Р ♀BV ♂ bv
═ x ═
bv bv
G ( BV) (bv)
некроссов. ( bv)
(Bv) (bV)
кроссов.
BV bv Bv bV
F1 ═ ═ ═ ═
bv bv bv bv
41,5% 41,5% 8,5% 8,5%
Причиной неполного сцепления генов является кроссинговер (перекрест хромосом) и обмен участками между гомологичными хромосомами в профазе I деления мейоза при созревании половых клеток. Кроссинговер происходит у самок, у самцов дрозофил кроссинговер не происходит (установлено, что для особей гетерогаметного пола характерна более низкая частота кроссинговер).
При изучении результатов скрещивания получается 4 фенотипа среди потомков:
17%(8, 5+8, 5) особей образовались из кроссоверных гамет
83%(41, 4+41, 5) особей образовались из некроссоверных гамет.
Частота кроссинговера зависит от расстояния между генами в хромосоме. Расстояние между генами выражается в процентах кроссинговера между ними и обозначается в морганидах.
Изучение Морганом сцепления генов представляет собой закономерное биологическое явление.
Гены, локализованные в одной хромосоме, наследуются, сцеплено, причем сила сцепления зависит от расстояния между генами. Эта закономерность получила название правило Моргана.
.
Основные положения хромосомной теории наследственности
Основные положения хромосомной теории наследственности сводятся к следующему:
- носителями наследственной информации являются хромо-
сомы и расположенные в них гены,
- гены расположены в хромосоме в линейном порядке друг за
другом в определенных локусах. Аллельные гены занимают
одинаковые локусы гомологичных хромосом,
- гены, расположенные в одной хромосоме, образуют группы
сцепления и наследуются преимущественно вместе. Число
групп сцепления равно гаплоидному набору хромосом,
- между гомологичными хромосомами возможен обмен участ-
ками - кроссинговер, который нарушает сцепление генов.
Сила сцепления зависит от расстояния между генами,
- процент кроссинговера пропорционален расстоянию между
генами. За единицу расстояния принимается 1 морганида,
которая равна 1% кроссинговера,
- при неполном сцеплении в сумме вероятность некроссовер-
ных гамет (гибридов) всегда больше, чем 50%,
- при расстоянии в 50 морганид и больше признаки наследуют-
ся независимо, несмотря на то, что локализованы в 1 хромо-
соме.
Лекция 11
Молекулярная генетика
План
1. Этапы развития молекулярной генетики.
2. Генетический код и его свойства.
3. Функционально-генетическая классификация генов.
4. Функциональная активность генов (экспрессия генов).
5. Регуляция экспрессии генов у про - и эукариот.
Молекулярная генетика занимается изучением структурно-функциональной организации генетического аппарата клеток и механизма реализации наследственной информации.
Этапы развития молекулярной генетики
Молекулярная генетика выделилась из биохимии и сформировалась как самостоятельная наука в 50-х годах прошлого столетия. Рождение этой науки связано с рядом важных биологических открытий:
1). В 20-40 гг. ХХ века было установлено, что носителем наследственной информации в клетке является молекула ДНК, а не белок, как считали раньше. Были получены прямые доказательства роли ДНК в наследственности. Это явления трансформации, трансдукции, половой процесс у бактерий, строение вируса, а также обнаружение почти полной идентичности химического состава ДНК (но не белков) у всех представителей данного биологического вида.
Данные о видовой специфичности строения ДНК получены в начале 50-х годов Эрвином Чаргаффом и его сотрудниками. Любая ДНК состоит из четырех типов нуклеотидов, содержащих одно из четырех азотистых оснований: аденин, тимин, гуанин, цитозин. Чаргафф с сотрудниками обнаружил, что соотношения между нуклеотидами, входящими в состав ДНК одинаковы в пределах вида и различаются у разных видов. Кроме того, число адениловых нуклеотидов равно числу тимидиловых, а число гуаниловых равно числу цитозиловых. Это открытие послужило главным ключом к выявлению структуры молекулы ДНК.
Трансформация у бактерий – это передача от одной бактериальной клетки другой наследственного материала. В 1928 году Фред Гриффит изучал трансформацию у двух штаммов пневмококка: вирулентного (вызывающего гибель мышей) и авирулентного (не вызывающего гибели).
Если убить вирулентные бактерии и вводить их вместе с живыми авирулентными, то часть животных все равно погибнет. Этот опыт свидетельствовал о том, что часть генетического материала из вирулентных бактерий проникла в авирулентные и сделала их вирулентными. После того как вирулентные клетки разделили на отдельные компоненты: углеводы, липиды, белки и ДНК, было обнаружено, что именно ДНК наделило живые бактерии генетическим признаком, которого у них до этого не было.
Трансдукция – это явление, заключающееся в том, что вирусы, выходя из бактериальной клетки, в которой они паразитировали, могут захватывать с собой часть ДНК и, перемещаясь в новые клетки передавать им признаки прежних хозяев.
Так, при внесении в неподвижные бактерии вирусов, размножавшихся в клетках подвижных культур, некоторые из бактерий становятся подвижными.
Половой процесс у бактерий – коньюгация, когда происходит обмен молекулами ДНК, и хотя новые бактерии не образуются, но их наследственный материал изменяется, т.к. происходит рекомбинация генетического материала.
Строение вируса или бактериофага: вирус состоит из молекулы ДНК (или РНК), заключенной в белковую оболочку. С помощью радиоизотопов было доказано, что именно ДНК проникает в клетку, а белковая оболочка остается снаружи. В результате образуются новые фаговые частицы. Таким образом, ДНК является носителем генетической информации и дает начало новым бактериофагам.
Косвенные доказательства: в соматических клетках разных органов содержится одинаковое количество ДНК, вдвое большее, чем в половых. Количество белков варьирует и не всегда больше, чем в половых.
2).Следующий этап развития молекулярной генетики связан с таким важным открытием, как установление структурной организации молекулы ДНК– в 1953 году. Крик и Уотсон установили, что ДНК состоит из двух спирально-закрученных цепей.
В настоящее время принято говорить о первичной, вторичной и третичной структуре ДНК: Первичной структурой ДНК называют линейную полинуклеотидную цепь, в которой мононуклеотиды соединены 3`,5`- фосфоэфирными связями. Модель Крика и Уотсона – это вторичная структура ДНК, основанная на принципе комплементарности и получившая название двойной спирали. Один виток спирали состоит из 10 нуклеотидов, размер витка 3,4 нм.
Дополнительное структурирование в пространстве двуспиральной молекулы ДНК (в конденсированных хромосомах) с образованием суперспирали является третичной структурой.
3).Определение направления передачи информации – «один ген – один фермент» - Бидл и Татум – 50-е годы. (В настоящее время – один ген – один полипептид).
4).Расшифровка генетического кода – Ниренберг, Очоа (К 1964 году расшифрованы коды для всех аминокислот). Выяснение механизма экспрессии генов у прокариот и ее регуляции – Франсуа Жакоб и Жан Моно – 50-е годы.
5).70-е годы и до настоящего времени – выявление особенностей экспрессии генов у эукариот. Развитие генетической инженерии.
Генетический код и его свойства
Генетический код – это система записи информации о последовательности расположения аминокислот в белках с помощью последовательности расположения нуклеотидов в молекуле ДНК.
Свойства генетического кода:
1.Код триплетен – каждая аминокислота зашифрована последовательностью трех нуклеотидов, называемых триплетом или кодоном.
2.Код вырожден – каждая аминокислота кодируется более чем одним кодоном. (Исключения – аминокислоты метионин и триптофан. Они кодируются одним кодоном.)
3.Код универсален – одни и те же триплеты кодируют одни и те же аминокислоты у всех организмов.
4.Генетический код не перекрывающийся – триплеты нуклеотидов не перекрывают друг друга. У некоторых вирусов обнаружено перекрывание, но это исключение.
5.Имеются стартовый и терминальный кодоны. Стартовый – единый для всех – АУГ. Терминальный трех видов – УАГ, УАА, УГА.
Функционально-генетическая классификация генов
В настоящее время ген рассматривается как единица функционирования наследственного материала. Ген – это участок молекулы ДНК, ответственный за синтез одного полипептида.
Различают три вида генов:
- структурные;
- модуляторы;
- регуляторы.
Структурные гены несут информацию об аминокислотах в белках, ферментах, а также о последовательности нуклеотидов в молекулах р-РНК и т-РНК.
Гены-модуляторы влияют на функционирование структурных генов, могут смещать в ту или иную сторону процесс развития признака. Они подразделяются на:
1)ингибиторы или супрессоры (эпистатичные гены),
2)интенсификаторы – могут повышать способность структурных генов к мутациям,
3)модификаторы – могут влиять на структурные гены по типу комлементарности.
Гены-регуляторы контролируют синтез регуляторных белков, а также время включения различных структурных генов в процессе индивидуального развития.
Свойства генов:
1. Дискретность действия – каждый ген действует как самостоятельная единица наследственности.
2. Стабильность – при отсутствии мутаций ген передается в ряду поколений в неизмененном виде.
3. Специфичность действия – каждый ген влияет на развитие своего определенного признака.
4. Плейотропия – способность одного гена обеспечивать развитие одновременно нескольких признаков (синдром Марфана).
5. Присутствие в виде двух у диплоидных и большего числа аллелей у полиплоидных организмов.
6. Действие гена дозировано, при изменении числа доз гена в организме изменяется признак (например, при болезни Дауна происходит увеличение до трех доз генов 21 хромосомы)
Функциональная активность генов или экспрессия генов
У прокариот она осуществляется в два этапа: транскрипция и трансляция.У эукариот есть еще стадия процессинга.
Экспрессия генов заключается в синтезе на молекуле ДНК молекулы и-РНК, комлементарной ей (или транскрипции – переписывание, считывание биологической информации) и дальнейшее ее использование для синтеза белка. Единицей транскрипции в ДНК является транскриптон, превышающий по размерам структурные гены. Транскриптон в клетках эукариот состоит из неинформативной (акцепторной) и информативной зоны. Неинформативная зона начинается геном-промотором (участок из 80 нуклеотидов), к которому присоединяется фермент РНК-полимераза, катализирующая процесс считывания. У прокариот один вид РНК-полимеразы, у эукариот три.
За геном промотором находятся гены-операторы, которые связывают регуляторные белки (белки, включающие и прекращающие транскрипцию).
Информативная зона состоит из структурных генов, располагающихся за генами операторами. Структурные гены эукариот разделены спейсерами – участками ДНК, не несущими информации.
Кроме того, в составе самих структурных генов есть информационные участки – экзоны и неинформационные – интроны. У пркариот спейсеров, экзонов и интронов нет.
В каждой фазе жизненного цикла в клетке транскрибируется только 10% структурных генов, а остальные гены не активны, но часть из них может включаться в других фазах жизненного цикла.
В результате транскрипции у прокариот сразу образуется м-РНК (зрелая РНК) и сразу же начинается процесс трансляции.
У эукариот транскрибируется большая молекула и-РНК, содержащая все неинформативные участки. Она называется РНК-предшественница или пре-РНК. Поэтому за транскрипцией наступает процессинг, в результате, которого разрушаются все неинформативные участки: акцепторная зона, спейсеры и интроны, а оставшиеся экзоны сшиваются (сплайсинг).
На этапе процессинга к начальному отрезку образовавшейся РНК присоединяется последовательность нуклеотидов, называемая колпачком, к концевому – последовательность остатков аденина – поли- A.
В процессинге происходит модификация нуклеотидов в РНК, например, их метилирование, гидрирование.
И только после этих превращений образуется зрелая м-РНК, которая начинается вводной последовательностью, называемой лидером, и заканчивается концевой последовательностью – трейлером. Лидер – вводная последовательность нуклеотидов, комплементарная последовательности в молекуле р-РНК малой субъединице рибосом, которая обеспечивает прикрепление и-РНК к малой субъединице. Трейлер включает нонсенс-кодон и поли-А последовательность.
Кодовым элементом м-РНК является триплет нуклеотидов, называемый кодоном. Каждому кодону соответствует определенная аминокислота.
Первичной структурой РНК является, как и в ДНК, линейная цепь полинуклеотидов, в которой мононуклеотиды соединены 3`,5`-фосфоэфирными связями.
Вторичная структура РНК – изогнутая цепь, а третичная представляет собой нить, намотанную на катушку, роль катушки играет особый транспортный белок – информатор.
Образующаяся м-РНК идет в цитоплазму к месту синтеза полипептида (белка), т.е. к рибосомам.
На рибосомах осуществляется процесс трансляции. Трансляция – это механизм, с помощью которого последовательность нуклеотидов в молекулах м-РНК переводится в специфическую последовательность аминокислот в полипептидной цепи.
Трансляция складывается из трех стадий:
1. Инициация – начало синтеза полипептида.
2. Элонгация – удлинение полипептида.
3. Терминация – окончание синтеза полипептида.
На этапе инициации меньшая субъединица рибосомы узнает стартовый кодон АУГ м-РНК и прикрепляется к ней. АУГ занимает первую позицию. После этого присоединяется большая субъединица рибосомы и в ней начинается собственно синтез белка. К большой субъединице подходит т-РНК с аминокислотой.
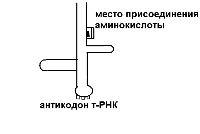
В молекуле т-РНК одна ее часть присоединяет аминокислоту, а другая – антикодон, спаривается с колоном м-РНК, определяющим эту аминокислоту.

В большой субъединице есть:
1 – аминоацильный участок
2 – пептидильный участок
В т-РНК, антикодон которой комплементарен кодону АУГ
м-РНК приносит аминокислоту метианин и останавливается в аминоацильном участке. Стадия инициации контролируется факторами инициации. У прокариот их 3, у эукариот 6-7.
Вторая стадия – элонгации начинается с перемещения т-РНК с метианином в пептидильный участок, а на ее место приходит другая т -РНК с новой аминокислотой. Между двумя аминокислотами образуется пептидная связь. Тем временем рибосома продвигается вдоль м-РНК, на рибосоме оказывается новый кодон, к которому вскоре присоединяется своим антикодоном соответствующая т-РНК.
Все это повторяется многократно, до тех пор, пока рибосома не
дойдет до одного из терминальных кодонов (УАА, УАГ, УГА).
Этим кодонам нет антикодона, нет аминокислоты. Наступает фаза терминации. К одной молекуле м-РНК прикрепляется обычно много
рибосом и образуются полисомы.
В процессе трансляции наряду с факторами инициации и элонгации важное значение имеет фермент, связывающий т-РНК с аминоацильным участком – аминоацил т-РНКаза.
Регуляция экспрессии генов у про- и эукариот
У прокариот она осуществляется на уровне промотора, оператора
и трансляции.
У эукариот регуляция происходит на всех этапах экспрессии: транскрипции, процессинга и трансляции.
Подробнее остановимся на регуляции экспрессии генов у бактерий.
Регуляция экспрессии генов у прокариот
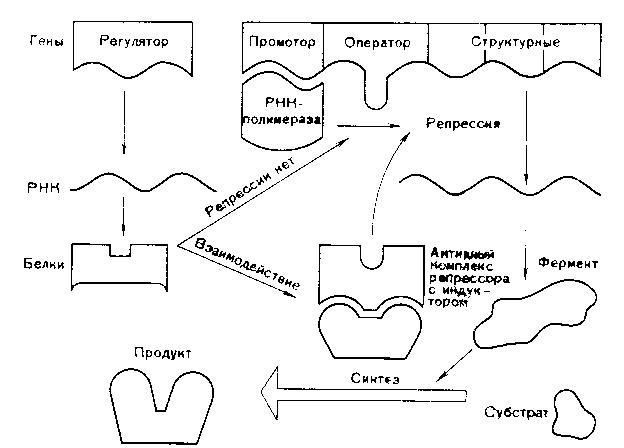
Схема регуляции транскрипции структурных генов прокариотической клетки по типу репрессии

Схема регуляции транскрипции структурных генов прокариотической клетки по типу индукции
В 1961 году Жакоб и Моно установили, что у бактерий под контролем гена-регулятора синтезируется белок-репрессор, который регулирует активность других генов. Белок-репрессор связывается с геном-оператором и блокирует его, вследствие чего транскрипция (считывание информации) становится невозможной и клетка не может синтезировать соответствующие белки. Но если в клетку проникает какой-либо субстрат, для расщепления которого необходимо синтезировать ферменты, то происходит следующее: субстрат присоединяется к репрессору и лишает его способности блокировать ген-оператор.
К ДНК присоединяются РНК-синтезирующие ферменты и начинается транскрипция генов. Синтез белка-фермента прекращается, если субстрат полностью расщепляется, белок-репрессор освобождается и снова блокирует гены-операторы. Это пример регуляции по типу индукции. Также существует регуляция по типу репрессии: когда конечные продукты биохимической реакции, соединяясь с неактивным белком-репрессором, образуют комплекс, блокирующий работу гена-оператора.
Антитерминация – заключается в том, что происходит игнорирование терминальных кодонов, процесс экспрессии продолжается.
Регуляция у эукариот
1)Регуляция генной активности у эукариот намного сложнее, чем у бактерий. У эукариот она происходит не только на уровне клетки. Существуют системы регуляции организма как целого. Огромную роль в регуляции играют гормоны, но регулируют они процессы синтеза белков лишь в клетках-мишенях. Гормоны связываются с белками-рецепторами, расположенными в мембранах таких клеток и включают системы изменения структуры клеточных белков. Те, в свою очередь, могут влиять как на процессы транскрипции, так и процессы трансляции. Каждый гормон через систему посредников активирует свою группу генов. Так адреналин включает синтез ферментов, расщепляющих гликоген мышц до глюкозы, а инсулин влияет на образование гликогена из глюкозы в печени.
2)На стадии транскрипции белки-гистоны участвуют в процессах регуляции генной активности у эукариот. Непременное условие – это деконденсация участка, где происходит транскрипция.
3)Регуляция на уровне трансляции направлена на сохранение стабильности м-РНК, а эффективность трансляции осуществляется с помощью факторов инициации, элонгации, терминации.
Лекция 12.
Изменчивость
План
1.Определение и формы изменчивости: модификационная, комбинативная, мутационная.
2.Мутагенные факторы.
3.Классификация мутаций.
4.Устойчивость и способы репарации генетического материала.
5.Закон гомологических рядов наследственной изменчивости Н.И.Вавилова.
Определение и формы изменчивости
Генетика изучает два основных свойства живых существ - наследственность и изменчивость.
Изменчивость - свойство организмов приобретать новые признаки и особенности индивидуального развития под влиянием факторов среды.
Изменчивость - один из важнейших факторов эволюции, обеспечивающих все многообразие живой природы.
Различают два вида изменчивости:
1. Фенотипическую (ненаследственную, модификационную);
2. Генотипическую (наследственную):
а) комбинативную;
б) мутационную.
Модификационная изменчивость – форма изменчивости, не связанная с изменением генотипа и вызванная влиянием факторов среды.
Модификационная изменчивость имеет особенности:
- не затрагивает наследственную основу организма и поэтому модификации не передаются по наследству, то есть от родителей к потомству,
- изменения направлены, происходят закономерно, их можно предсказать,
- имеют адаптивное (приспособительное) значение,
- имеют массовый (групповой) и обратимый характер,
- определенный фактор внешней среды вызывает сходное изменение у всех особей данного вида.
Модификационная изменчивость имеет предел. Границы изменчивости признака, обусловленные генотипом называются нормой реакции. Она может быть узкой, когда признак изменяется незначительно (цвет глаз), и широкой, когда признак изменяется в широких пределах (рост, масса тела).
В медицине часто приходится устанавливать норму реакции для оценки max и min количественных показателей (уровень гормонов, ферментов, гемоглобина и др.)
Комбинативная изменчивость – это наследственная изменчивость, обусловленная перекомбинацией имеющихся генов и хромосом, без изменения структуры генов и хромосом (наследственного материала). Этот тип изменчивости проявляется уже на стадии образования половых клеток.
Источниками комбинативной изменчивости являются процессы, происходящие в мейозе и в результате оплодотворения:
1. Рекомбинация генов при кроссинговере в профазе1 мейоза.
2. Рекомбинация хромосом в ходе мейоза (независимое расхождение хромосом и хроматид при мейозе)
3. Комбинация хромосом в результате слияния гамет при оплодотворении (случайное сочетание гамет при оплодотворении).
Комбинативная изменчивость обеспечивает генотипическое разнообразие людей, объясняет наличие признаков у детей и внуков от родственников по отцовской и материнской линии.
Мутационная изменчивость – способность генетического (наследственного) материала изменяться и эти изменения наследуются в потомстве.
В основе мутационной изменчивости лежат мутации.
Мутации – это внезапные изменения генетического материала под влиянием среды и передающиеся по наследству.
Частота мутаций зависит от вида организма, от возраста, от фазы онтогенеза, стадии гаметогенеза, может происходить в половых и соматических клетках, иметь рецессивный и доминантный характер. Например, у человека до 6% гамет несут мутантные гены.
Процесс образования мутаций называется мутагенезом.
Факторы, вызывающие мутации называются мутагенными.
Мутации первоначально действуют на генетический материал особи, а через генотип изменяется и фенотип.
Мутагенные факторы
Факторы, вызывающие мутации называются мутагенными факторами (мутагенами) и подразделяются на:
1. Физические;
2. Химические;
3. Биологические.
К физическим мутагенным факторам относятся различные виды излучений, температура, влажность и др. Наиболее сильное мутагенное действие оказывает ионизирующее излучение – рентгеновские лучи, α-, β-, γ- лучи. Они обладают большой проникающей способностью.
При действии их на организм они вызывают:
а) ионизацию тканей – образование свободных радикалов (ОН) или (Н) из воды, находящейся в тканях. Эти ионы вступают в химическое взаимодействие с ДНК, расщепляют нуклеиновую кислоту и другие органические вещества;
б) ультрафиолетовое излучение характеризуется меньшей энергией, проникает только через поверхностные слои кожи и не вызывает ионизацию тканей, но приводит к образованию димеров (химические связи между двумя пиримидиновыми основаниями одной цепочки, чаще Т-Т). Присутствие димеров в ДНК приводит к ошибкам при ее репликации, нарушает считывание генетической информации;
в) разрыв нитей веретена деления;
г) нарушение структуры генов и хромосом, т.е. образование генных и хромосомных мутаций.
К химическим мутагенам относятся:
- природные органические и неорганические вещества (нитриты, нитраты, алкалоиды, гормоны, ферменты и др.);
- синтетические вещества, ранее не встречавшиеся в природе (пестициды, инсектициды, пищевые консерванты, лекарственные вещества).
- продукты промышленной переработки природных соединений – угля, нефти.
Механизмы их действия:
а) дезаминирование – отщепление аминогруппы от молекулы аминокислот;
б) подавление синтеза нуклеиновых кислот;
в) замена азотистых оснований их аналогами.
Химические мутагены вызывают преимущественно генные мутации и действуют в период репликации ДНК.
К биологическим мутагенам относятся:
- Вирусы (гриппа, краснухи, кори)
- Невирусные паразитические организмы (грибы, бактерии, простейшие, гельминты)
Механизмы их действия:
а) вирусы встраивают свою ДНК в ДНК клеток хозяина.
б) продукты жизнедеятельности паразитов-возбудителей болезней действуют как химические мутагены.
Биологические мутагены вызывают генные и хромосомные мутации.
Классификация мутаций
Различают следующие основные типы мутаций:
1. По способу возникновения их подразделяют на спонтанные и индуцированные.
Спонтанные – происходят под действием естественных мутагенных факторов внешней среды без вмешательства человека. Они возникают в условиях естественного радиоактивного фона Земли в виде космического излучения, радиоактивных элементов на поверхности земли.
Индуцированные мутации вызываются искусственно воздействием определенных мутагенных факторов.
2. По мутировавшим клеткам мутации подразделяются на генеративные и соматические.
Генеративные – происходят в половых клетках, передаются по наследству при половом размножении.
Соматические – происходят в соматических клетках и передаются только тем клеткам, которые возникают из этой соматической клетки. Они не передаются по наследству.
3. По влиянию на организм:
Отрицательные мутации – летальные (несовместимые с жизнью); полулетальные (снижающие жизнеспособность организма); нейтральные (не влияющие на процессы жизнедеятельности); положительные (повышающие жизнеспособность). Положительные мутации возникают редко, но имеют большое значение для прогрессивной эволюции.
4. По изменениям генетического материала мутации подразделяются на геномные, хромосомные и генные.
Геномные мутации – это мутации, вызванные изменением числа хромосом. Могут появляться лишние гомологичные хромосомы. В хромосомном наборе на месте двух гомологичных хромосом оказываются три – это трисомия. В случае моносомии наблюдается утрата одной хромосомы из пары. При полиплоидии происходит кратное гаплоидному увеличение числа хромосом. Еще один вариант геномной мутации – гаплоидия, при которой остается только одна хромосома из каждой пары.
Хромосомные мутации связаны с нарушением структуры хромосом. К таким мутациям относятся утраты участков хромосом (делеции), добавление участков (дупликация) и поворот участка хромосом на 180° (инверсия).
Генные мутации, при которых изменения происходят на уровне отдельных генов, т.е. участков молекулы ДНК. Это может быть утрата нуклеотидов, замена одного основания на другое, перестановка нуклеотидов или добавление новых.
Устойчивость и репарация генетического материала
Устойчивость к изменениям генетического материала обеспечивается:
1. Диплоидным набором хромосом.
2. Двойной спиралью ДНК.
3. Вырожденностью (избыточностью) генетического кода
4. Повтором некоторых генов.
5. Репарацией нарушений структуры ДНК
Наличие механизмов репарации – обязательное условие существования биологических существ.
Репарация генетического материала – это процесс, обеспечивающий восстановление поврежденной структуры молекулы ДНК.
В ДНК клетки ежедневно происходит множество случайных изменений.
Большинство эффективно исправляются (репарируются) с помощью специальных ферментных систем.
Впервые репарация молекулы ДНК была установлена в 1948 году. А в 1962 году был описан один из способов репарации – световая репарация или фотореактивация.
Было установлено, что при ультрафиолетовом облучении вирусов-фагов, бактерий и простейших наблюдается резкое снижение их жизнедеятельности, даже гибель.
Если воздействовать на них видимым светом, то выживаемость их значительно увеличивается.
Оказалось, что под действием ультрафиолета в молекуле ДНК образуются димеры (химические связи между двумя основаниями одной цепочки, чаще Т-Т), образование димеров препятствует считыванию информации.
Видимый свет активирует ферменты, разрушающие димеры.
Второй способ репарации – темновая репарация, была изучена в 50-е годы ХХ века.
Темновая репарация протекает в четыре стадии с участием четырех групп ферментов. Ферменты образовались в ходе эволюции и направлены на поддержание стабильности генетической информации клетки.
1. Фермент эндонуклеаза находит поврежденный участок и рядом с ним разрывает нить ДНК.
2. Фермент эктонуклеаза «вырезает» (удаляет) поврежденный участок.
3. ДНК-полимераза по принципу комплементарности синтезирует фрагмент ДНК на месте разрушенного.
4. Лигаза «сшивает» синтезированный фрагмент с основной нитью ДНК.
Доказана возможность репарации ДНК при повреждении обеих ее нитей. При этом информация может быть получена с и-РНК (фермент ревертаза).
Закон гомологических рядов наследственной изменчивости Н.И.Вавилова
Известно, что мутирование происходит в различных направлениях. Однако, это многообразие подчиняется определенной закономерности, обнаруженной в 1920 году Н.И.Вавиловым. Он сформулировал закон гомологических рядов наследственной изменчивости: «Виды и роды, генетически близкие, характеризуются сходными рядами наследственной изменчивости с такой правильностью, что зная ряд форм в пределах одного вида, можно предвидеть существование параллельных форм у других видов и родов».
Можно сказать, что у родственных видов, имеющих общее происхождение возникают сходные мутации.
Одни и те же болезни встречаются у животных и у человека. Так, например, альбинизм наблюдается во всех классах позвоночных животных и человека. Брахидактилия (короткопалость) отмечена у крыс, овец, собак, человека. Мышечная дистрофия – у мышей, крыс, лошадей, человека.
Закон гомологических рядов позволяет предвидеть возможность появления мутаций, которые могут быть использованы в селекции для создания новых ценных для хозяйства форм.
Лекция 13
Методы изучения генетики человека
План
1.Генеалогический метод.
2.Близнецовый метод.
3.Метод дерматоглифики.
4.Цитогенетический метод.
5.Метод гибридизации соматических клеток.
6.Онтогенетический метод.
7.Популяционно-статистический метод.
8.Метод моделирования.
9.Иммунологический метод.
10.Биохимический метод.
Генеалогический метод
Типы наследования и формы проявления генетических задатков у человека весьма многообразны и для дифференциации между ними требуются специальные методы анализа, в первую очередь – генеалогический, предложенный Ф.Гальтоном.
Генеалогический метод или изучение родословных предусматривает прослеживание признака в семье или роду с указанием типа родственных связей между членами родословной. В медицинской генетике этот метод обычно называют клинико-генеалогическим, поскольку речь идет о наблюдении патологических признаков с помощью приёмов клинического обследования. Генеалогический метод относится к наиболее универсальным методам в генетике человека. Он широко применяется при решении теоретических и практических проблем:
1) для установления наследственного характера признака,
2) при определении типа наследования и пенетрантности генотипа,
3) выявление сцепления генов и картирование хромосом,
4) при изучении интенсивности мутационного процесса,
5) при расшифровке механизмов взаимодействия генов,
6) при медико-генетическом консультировании.
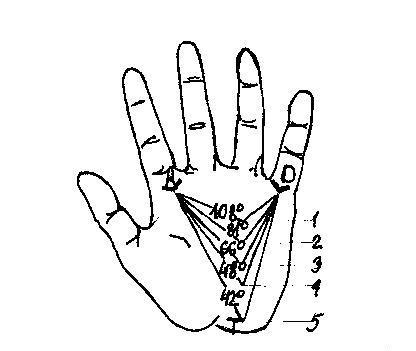
Суть генеалогического метода сводится к выяснению родственных связей и прослеживанию признака среди близких и дальних прямых и непрямых родственников. Технически он складывается из двух этапов: составления родословных и генеалогического анализа.
Составление родословной
Сбор сведений о семье начинается с пробанда, которым называется лицо, первым попавшее в поле зрения исследователя.
Дети одной родительской пары (родные братья и сестры) называются сибсами. Семьей в узком смысле, или ядерной семьей, называют родительскую пару и их детей. Более широкий круг кровных родственников лучше обозначать термином «род». Чем больше поколений вовлекается в родословную, тем она обширнее. Это влечёт за собой неточность полученных сведений и, следовательно, неточность родословной в целом. Часто люди не знают даже числа своих двоюродных братьев и сестер, не говоря уже о каких-то признаках у них и их детей.
Для наглядности готовят графическое изображение родословной. Для этого обычно пользуются стандартными символами. Если рассматриваемых признаков в родословной много, то можно прибегать к буквенным или штриховым различиям внутри символов. Схема родословной обязательно сопровождается описанием обозначений под рисунком – легендой, что исключает возможность неправильных истолкований.
Генеалогический анализ
Целью генеалогического анализа является установление генетических закономерностей.
1 этап – установление наследственного характера признака. Если в родословной встречается один и тот же признак несколько раз, то можно думать о наследственной его природе. Однако надо прежде всего исключить возможность экзогенного накопления случаев в семье или роду. Например, если один и тот же патогенный фактор действовал на женщину во время всех беременностей, то у нее могут родиться несколько детей с одинаковыми аномалиями. Или же какой-то фактор действовал на многих членов семьи, необходимо сличить действие сходных внешних факторов. С помощью генеалогического метода были описаны все наследственные болезни.
2 этап – установление типа наследования и пенетрантности гена. Для этого используют принципы как генетического анализа, так и статистические методы обработки данных из родословной.
3 этап – определение групп сцепления и картирования хромосом, до недавнего времени основывающегося только на генеалогическом методе. Выясняют сцепленные признаки и процесс кроссинговера. Этому способствуют разработанные математические методы.
4 этап – изучение мутационного процесса. Он применяется в трех направлениях: при изучении механизмов возникновения мутаций, интенсивности мутационного процесса и факторов, вызывающих мутации. Особенно широко генеалогический метод применяется при изучении спонтанных мутаций, когда надо различать «спорадически» возникшие случаи от «семейных».
5 этап – анализ взаимодействия генов в клинической генетике был сделан С.Н.Давиденковым (1934, 1947) по анализу полиморфизма заболеваний нервной системы.
6 этап – в медико-генетическом консультировании для составления прогноза без генеалогического метода обойтись нельзя. Выясняют гомо- или гетерозиготность родителей и рассматривают вероятность рождения детей с теми или иными признаками.
Близнецовый метод исследования
Исследование близнецов – один из основных методов генетики человека. Существуют однояйцевые близнецы, возникающие из одной яйцеклетки, оплодотворенной одним сперматозоидом. Возникают они из-за разделения зиготы на два генетически идентичных друг другу и всегда однополых зародыша.
Разнояйцовые близнецы развиваются из разных яйцеклеток, оплодотворенных разными сперматозоидами. Генетически они различаются как братья и сестры одних родителей.
При помощи близнецового метода можно изучить:
1) Роль наследственности и среды в формировании физиологических и патологических особенностей организма. В частности, изучение наследственной передачи людьми некоторых болезней. Изучение экспрессивности и пенетрантности генов, вызывающих наследственные заболевания.
2) Конкретные факторы, усиливающие или ослабляющие влияние внешней среды.
3) Корреляцию признаков и функций.
Особенно важна роль близнецового метода в изучении проблемы «генотип и среда».
Сравнивают обычно три группы близнецов: ДБ в одинаковых условиях, ОБ в одинаковых условиях, ОБ в разных условиях.
При изучении близнецов определяют частоту, степень совпадения (конкордантности) тех или иных признаков.
При изучении роли наследственности в происхождении того или иного признака производят расчет по формуле К.Хольцингера.
Коэффициент наследуемости - Н
Н =
% сходства ОБ - % сходства РБ
=
% сходства ОБ - % сходства РБ
100 - % сходства РБ
При Н=1 вся изменчивость в популяции обусловлена наследственностью.
При Н=0 вся изменчивость вызвана средовыми факторами. Влияние среды С выражается формулой: С=100% - Н, где Н – коэффициент наследуемости. Например, конкордантность монозиготных (однояйцевых) близнецов 3%. Тогда Н =
67 – 3 = 64 = 0,7 или 70 %. С = 100 – 70 = 30 %
100 – 3 97
Итак, данный признак на 70% обусловлен наследственностью, а на 30% - влиянием факторов внешней среды.
Другой пример. Группы крови по системе АВО у ОБ =100%, т.е. полностью зависит от наследственности.
Частота совпадения групп крови и некоторых заболеваний у близнецов (в %)
|
№ п/п |
Признаки или болезни |
ОБ |
РБ |
|
1 |
группы крови АВО |
100 |
64 |
|
2 |
корь |
98 |
94 |
|
3 |
коклюш |
97 |
93 |
|
4 |
шизофрения |
86 |
14 |
|
5 |
свинка |
82 |
74 |
|
6 |
эпилепсия |
67 |
3 |
|
7 |
Врожденный стеноз привратника |
67 |
3 |
Метод дерматоглифики
Это наука, изучающая наследственную обусловленность рисунков, которые образуют линии кожи на кончиках пальцев, ладонях и подошвах человека.
Оказалось, что у каждого народа, у каждой расы, у каждого человека рисунки имеют свои особенности, и на ладонях они строго индивидуальны. На это впервые обратил внимание Ф.Гальтон, который предложил английской уголовной полиции по отпечаткам пальцев идентифицировать преступников.
Дерматоглифические исследования имеют важное значение в криминалистике, в определении зиготности близнецов, в диагностике ряда наследственных заболеваний, а так же в отдельных случаях спорного отцовства.
Ладонный рельеф очень сложен. В нем выделяют ряд полей, подушечек и ладонных линий. Подушечек на ладони 11, их делят на 3 группы:
1)пять концевых (эпликальных) подушечек на концевых фалангах пальцев.
2)четыре межпальцевые подушечки, располагаются против межпальцевых промежутков.
3)две ладонные проксимальные подушечки тенар и гипотенар. У основания большого пальца – тенар, у противоположного края ладони- гипотенар.
На наиболее возвышенных частях подушечек заметны кожные гребешки. Это линейные утолщения эпидермиса, которые представляют собой модифицированные чешуйки кожи. Кожные гребешки идут потоками, как на ладонях, так и на пальцевых подушечках. Точки встречи этих потоков образуют трирадиусы или дельты.
Гребешковые узоры обычно изучают под лупой. Отпечатки узоров, при помощи типографской краски, делают на чистой белой, лучше мелованной, бумаге или целлофане. Как на кончиках пальцев, так и на ладонных возвышениях могут наблюдаться различные папиллярные узоры в виде завитков, петель и дуг, открытых в ульпарную или радиальную стороны. На тенаре и гипотенаре чаще бывают дуги. На средней и основной фалангах пальцев гребешковые линии идут поперек пальцев, образуя различные узоры – прямые, серповидные, волнообразные, дугообразные и их сочетания. В среднем на одном пальце бывают 15-20 гребешков.
Рисунок ладони.
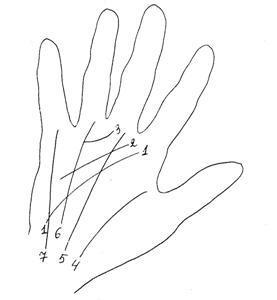
1 – поперечная проксимальная борозда, линия прижатия 4 пальцев
2 - поперечная средняя борозда, линия прижатия 3 пальцев
3 – поперечная дистальная борозда, линия прижатия 2 пальцев
4 – борозда большого пальца
5 – продольная срединная борозда от запястья к основанию 3го пальца
6 – продольная промежуточная борозда от запястья к основанию 4го пальца
7 – продольная ульнарная борозда, от запястья к основанию 5го пальца
1 – синдром Патау
2 – синдром Дауна
3 – синдром Шерешевского-Тернера
4 – норма
5 – синдром Клайнфельтера
При изучении кожного рельефа ладони исследуют:
1) Ход главных ладонных линий А, В, С, Д 1,2,3,4,5,6,7.
2) Ладонные узоры на тенаре и гипотенар.
3) Пальцевые узоры (форму узоров, гребневый счет)
4) Осевые трирадиусы.
Аналогичные исследования проводят и на подошвах ног. Направление главной ладонной линии Д у родителей и их детей одинаковое.
Изучение больных с хромосомными болезнями (болезнь Дауна, синдром Клайнфельтера) показало, что у них меняется не только рисунок пальцевых и ладонных узоров, но и характер основных сгибательных борозд на коже ладоней.
Несколько меньшую выраженность имеют дерматоглифические отклонения у больных с такими дефектами развития, как врожденные пороки сердца и магистральных сосудов, незаращения мягкого и твердого неба, верхней губы и т.д.
Установлены изменения в характере пальцевых и ладонных узоров при проказе, шизофрении, сахарном диабете, раке, ревматизме, полиомиелите и других заболеваниях.
Цитогенетический метод
Этот метод позволяет с помощью микроскопа исследовать структуры клетки – хромосомы. С помощью метода микроскопии изучен кариотип организма человека (хромосомный набор клеток организма). Установлено, что многие заболевания и дефекты развития связаны с нарушением числа хромосом и их строения. Этот метод позволяет изучить также действие мутагенов на состав и строение хромосом. Цитогенетический метод связан с временными культурами тканей (обычно лейкоцитов) и получением метафазных ядер с укороченными, утолщенными хромосомами, деление которых останавливают на стадии метафазной пластинки колхицином. Если в кариотипе изучаются половые хромосомы, то этот метод позволяет исследовать половой хроматин в соматических клетках.
Гибридизация соматических клеток
Гибридные клетки обладают определенными свойствами, позволяющими определять локализацию гена или сцепление гена. Потеря хромосом человека из некоторых типов гибридных клеток позволяет получать клоны с отсутствием определенной хромосомы. Наиболее употребительны гибриды соматических клеток человек – мышь.
Прослеживание за наличием биохимического генетического маркера в гибридных клонах по мере элиминации хромосом человека может привести к обнаружению локализации гена, если признак исчезает из клеток, как только они меняются определенными хромосомами. Цитогенетический анализ большого числа клонов и сопоставления результатов с присутствием большого числа генетических маркеров позволяет подметить сцепленные гены и их локализацию. Дополнительно используют информацию, при использовании клонов от инвалидов с транслокациями и другими хромосомными аномалиями.
Этим методом была установлена локализация гена фосфоглицераткиназы в длинном плече Х-хромосомы, т.е. место гибридных клеток позволяет установить:
1) локализацию гена
2)сцепление генов
3)картирование хромосом
Свыше 160 локусов определены с помощью метода гибридных соматических клеток.
Онтогенетический метод
Позволяет изучить закономерности проявления какого-либо признака или заболевания в процессе индивидуального развития. Выделяют несколько периодов развития человека. Антенатальный (развитие до рождения) и постнатальный. Большинство признаков человека формируются в фазу морфогенеза антенатального периода. В фазу морфогенеза постнатального периода заканчивается формирование коры головного мозга и некоторых других тканей и органов, формируется иммунологическая система организма, которая достигает наивысшего развития через 5-7 лет после рождения ребенка. В постморфогенетический период развиваются вторичные половые признаки.
В морфогенетический период изменение активности генов происходит по двум типам:
1)включение и выключение генов
2)усиление и ослабление действия генов
В постморфогенетический период развития первый тип изменения активности генов почти отсутствует, происходит лишь небольшое включение отдельных генов – например, генов, определяющих вторичные половые признаки, развитие некоторых наследственных заболеваний. Выключение же генов в этом периоде более значительное. Репрессируется активность многих генов, связанных с выработкой меланина (в результате происходит поседение), а также генов, связанных с выработкой γ-глобулинов (повышается восприимчивость к заболеваниям). Подавляются многие гены в клетках нервной системы, мышечных клетках и т.д.
Репрессия генов осуществляется на уровне транскрипции, трансляции, посттрансляции. Однако основной тип изменения активности генов на этом этапе – усиление и ослабление действия генов. Может изменяться доминирование генов, что вызывает изменение внешних признаков, особенно в период полового созревания. Меняется соотношение половых гормонов и соответственно признаки пола. Репрессивные гены с возрастом могут оказывать большое влияние на развитие того или иного признака. Например, ген фенилкетонурии в гетерозиготном состоянии изменяет психику человека.
Популяционно-статистический метод исследования
Представляет собой метод математического подсчета тех или иных генов и соответствующих признаков в определенных популяциях. Теоретической основой данного метода является закон Харди-Вайнберга.
Этим методом установлено, что все гены человеческой популяции по частоте встречаемости можно разделить на 2 категории:
1)имеющие универсальное распространение, к которым относится большинство генов. Например, ген дальтонизма, имеющийся у 7%мужчин и более чем у 13%женщин. Ген амавротической идиотии, встречающийся у населения Европы с частотой 4 на 10 000 населения.
2)гены, встречающиеся преимущественно в определенных районах. Например, ген серповидно-клеточной анемии распространен в странах, где свирепствует малярия. Ген врожденного вывиха бедра, имеющий высокую концентрацию у аборигенов северо-востока нашей страны.
Метод моделирования
Закон гомологических рядов Н.И.Вавилова (виды и роды генетически близкие обладают сходными рядами наследственной изменчивости) позволяет с определёнными ограничениями экстраполировать экспериментальные данные на человека.
Биологическая модель наследственного заболевания на животном часто является более удобной для исследования, чем больной человек. Оказалось, что у животных имеется около 1300 наследственных болезней, так же, как у человека. Например, у мышей – 100, у крокодилов – 50, у крыс – 30. на модели гемофилии А и В у собак показано, что она обусловлена рецессивным геном, расположенным в Х-хромосоме.
Моделирование мышечной дистрофии у мышей, хомяков и кур дало возможность понять патогенетическую сущность этого заболевания. Было установлено, что при этом заболевании поражается не нервная система, а непосредственно мышечные волокна.
Начальные механизмы галактоземии были выяснены на модели кишечной палочки. И у человека, и у бактерий неспособность усваивать галактозу вызвана одинаковым наследственным дефектом – отсутствием активного фермента – галактоза-1-фосфатилуридилтрансферазы.
Иммунологический метод исследования
Этот метод основан на изучении антигенного состава клеток и жидкостей человеческого организма – крови, слюны, желудочного сока и т.п. Чаще всего исследуют антигены форменных элементов крови: эритроцитов, лейкоцитов, тромбоцитов, а также белков крови. Различные виды антигенов эритроцитов образуют системы групп крови.
В начале ХХ столетия К.Ландштейнер и Я.Янский показали, что в зависимости от характера реакций между эритроцитами и плазмой крови, всех людей можно разделить на 4 группы. В дальнейшем было доказано, что реакции эти происходят между белковыми веществами эритроцитов, которые были названы агглютино-генами, и белками сыворотки крови, которые были названы агглютининами.
Группы крови определяются антигенами, содержащими липидную и белковые фракции, и которые находятся на поверхности эритроцитов. Белковая часть антигена контролируется геном, который работает на ранних этапах развития эритроцита. Антигены специфичны для каждой группы крови.
Всего сейчас известно 14 систем эритроцитарных групп крови, в которые входят более 100 различных антигенов. В системе групп крови АВО на поверхности эритроцитов формируется два антигена под контролем генных аллелей Iа, Iв.
Бернштейн в 1925 году показал, что есть третья аллель Iо, которая не контролирует синтез антигена. Таким образом, в системе АВО групп крови существует три аллеля, но у каждого человека имеется только два из них. Если расписать возможные мужские и женские гаметы в решетке Пеннета, то можно проследить, какие возможные комбинации групп крови будут у потомков.
Группы крови АВО у потомков в зависимости от групп крови у родителей
Иммунологические методы применяют для обследования пациентов и их родственников при подозрении на иммунодефецитные состояния (агаммаглобулинемия, дисгаммаглобулинемия, атаксия-телеангиэктазия и другие), при подозрении на антигенную несовместимость матери и плода, при пересадке органов и тканей, при установлении истинного родства, в случаях медико-генетического консультирования, при необходимости изучения генетических маркеров при диагностике сцепления генов или при определении наследственной предрасположенности к заболеваниям, при установлении зиготности близнецов.
Определение групповой принадлежности крови имеет практическое значение в разнообразных генетических исследованиях:
1)при установлении зиготных близнецов
2)при установлении сцепления генов.
3) в судебно-медицинской экспертизе в случаях спорного отцовства или материнства. Известно, что у ребенка не могли появится антигены, которых нет у родителей.
Система групп крови М была открыта К.Ландштейнером и И.Левиным в 1927 году (в этой группе антитела к соответствующим антигенам не продуцируются). В системе имеется два аллеля M,N.
Гены, определяющие фактор М и N, являются кодоминантными, т.е. если встречаются вместе, то оба и проявляются. Таким образом, существуют гомозиготные генотипы MM и NN, и гетерозиготные MN. В популяциях европейцев генотипы ММ встречаются примерно в 36%, NN – в 16%, MN – в 48%.
А гены соответственно:
М=36 + 48 = 60%
 2
2
 N=16
+ 48 = 40%
N=16
+ 48 = 40%
2
Резус – фактор
Как показали исследования ученых, 85% европейцев имеют эритроцитарный антиген, общий с антигеном обезьян вида макака-резус. У 15% людей резус-антигенов на поверхности эритроцитов нет.
Система групповых резус-антигенов очень сложна. Предполагается, что резус-антигены контролируются тремя тесно сцепленными локусами С, Д и Е в двух хромосомах и наследуются доминантно. Поэтому возможны по каждому локусу три генотипа: гомозиготные резус-положительные, гетерозиготные резус-положительные и гомозиготные резус-отрицательные.
Наиболее иммуногенным является антиген Д. Антигены С и Е являются менее активны.
В 1962 году было установлено наличие эритроцитарного изоантигена Хд, передающегося через половую Х-хромосому. По этому антигену всех людей можно разделить на Хд-положительных и Хд-отрицательных. Среди Хд-положительных женщин встречается 88%, а среди мужчин – 66%. Если оба родителя Хд-отрицательны, то все их дети (как девочки, так и мальчики) будут Хд-отрицательны. Если отец Хд-положительный, а мать Хд-отрицательна, то их дочери будут Хд-положительными, а сыновья Хд-отрицательными. Если мать Хд-положительна, а отец Хд-отрицателен, то их сыновья будут Хд-положительными т.е. тип наследования «крест-накрест». Дочери же могут быть как Хд-положительными, так и Хд-отрицательными, в зависимости от гомозиготности матери. Ген Хд – группы локализован в коротком плече Х-хромосомы. Система Хд используется для изучения анеуплоидий (аномального числа Х-хромосом у ребенка с трисомией Х, синдрома Клайнфельтера, синдрома Шерешевского-Тернера и др.). Предполагается, что Хд-несовместимость матери и плода (мать Хд-отрицательная, а плод Хд-положительный) приводит к уменьшению частоты рождения девочек.
Биохимический метод
Позволяет, с одной стороны, изучить количество ДНК в клетках человека в норме и патологии, с другой – определять наследственные дефекты обмена веществ при помощи:
1)определения аномального белка (структурных белков или ферментов), которые образуются в результате биохимических реакций;
2)определения промежуточных продуктов обмена, которые появляются вследствие генетического блока прямой реакции обмена.
Например, при фенилкетонурии аминокислота фенилаланин не превращается в тирозин. Происходит увеличение её концентрации в крови и уменьшение концентрации тирозина. Фенилаланин при этом превращается в фенилпировиноградную кислоту и ее производные – фенилмолочную, фенилуксусную и фенилацетилглутаминовую.
Эти соединения обнаруживают в моче больного при помощи хлорного железа FeCl3 или 2,4 – динитрофенилгидразина.
Лекция14