

Лекция №5.
Тема: Организация наследственного материала.(II)
1. Уровни упаковки генетического материала.
2. Уровни структурно – функциональной организации наследственного материала.
3. Биосинтез белка в клетке.
4. Регуляция транскрипции у прокариот и эукариот.
5. Цитоплазматическая наследственность.
6. Генная инженерия.
1. Длина хромосом 0,2 – 5,0 мкм, их диаметр 0,2 – 2 мкм. Хромосомная ДНК эукариотической клетки упакована исключительно компактно. Например, самая маленькая хромосома человека – 22, содержит примерно 4.6ּ107п.н., что соответствует длине 1,4 см. Во время митоза эта хромосома укорачивается до 2 мкм, т.е. становится в 7000 раз компактнее. Очевидно, чтобы достичь такой плотности упаковки и сохранить эффективность основных генетических процессов, структура хромосомы должна иметь несколько уровней организации. Упаковка генетического материала достигается спирализацией (конденсацией) и связью его с белками.

Рис.1.Уровни компактизации хроматина, следующие после образования нуклеосом.
Нуклеосомный уровень. Нуклеосома – это глобула (октаэдр), содержащая по 2 молекулы четырех гистонов – (Н2А, Н2В, Н3, Н4), вокруг которой двойная спираль ДНК образует 2,2 витка (200 пар нуклеотидов). Нуклеосомная нить имеет d=10 -13нм. Длина ДНК уменьшается в 5 – 7 раз.
Супернуклеосомный уровень (соленоид).Нуклеосомная нить конденсируется, нуклеосомы «сшиваются» гистоном Н1, и образуется спираль d=25 нм. Виток спирали содержит 6 – 10 нуклеосом. Укорочение ДНК еще в 6 раз.
Хроматидный (петлевой) уровень.Супернуклеосомная нить спирализуется с образованием петель и изгибов, составляет основу хроматиды. Обнаруживается в профазе петель d=50нм. Нить ДНП укорачивается еще в 10 – 20 раз.
Уровень метафазной хромосомы.Хроматиды спирализуются и образуют эухроматиновые (слабо спирализованные) и гетерохроматиновые (сильно спирализованные) участки; происходит укорочение ДНП еще в 20 раз. Длина метафазных хромосом 2,3 – 1,1 мкм, d=0,2 – 5,0 мкм. Общий итог конденсации – укорочение нити ДНП в 10 000 раз.
2.Структурно – функциональными уровнями организации наследственного материала являются генный, хромосомный и геномный.
Ген – элементарная структура генного уровня организации. Так как гены относительно независимы друг от друга, возможно дискретное (раздельное) и независимое наследование (по третьему закону Менделя) и изменение отдельных признаков вследствие генных мутаций.
Гены эукариот находятся в хромосомах, образуя хромосомный уровень организации наследственного материала. Все гены одной хромосомы составляют группу сцепления и передаются вместе с этой хромосомой. На этом уровне происходит перекомбинация генов родителей у потомков при половом размножении и изменения структуры отдельных хромосом.
Набор генов, получаемых потомком о родителей, составляет его генотип. Геном – это гены гаплоидного набора хромосом. Действие генов в разных генотипах проявляется по – разному.Взаимодействуют между собой гены как одной хромосомы, так и разных хромосом. Нарушение набора хромосом приводит к геномным мутациям.
3.Биосинтез белка состоит из: транскрипции, активации аминокислот и узнавания ими своей т-рнк (рекогниция), трансляции.
Транскрипция: информация о первичной структуре белка закодирована в соответствующем участке ДНК. Посредник, копирующий и передающий эту информацию и-РНК. РНК – полимераза расщепляет молекулу ДНК, и на одной ее цепи по принципу комплементарности выстраиваются нуклеотиды РНК. Такая и-РНК является комплементарной копией одной из цепочек ДНК. И-РНК выходит в цитоплазму и располагается в малой субъединице рибосомы.
Рекогниция – узнавание каждой аминокислот своей т-РНК. Вначале аминокислота активируется и взаимодействует с т-РНК, образуя аминоацил-т-РНК. Т-РНК «приносят» аминокислот в рибосому.
Трансляция – перевод последовательности нуклеотидов и-РНК в последовательность аминокислот полипептида. Она начинается со связывания и-РНК с субъединицой рибосомы. Начальный этап трансляции – инициация: при этом к рибосоме всегда присоединяется метионин – т-РНК. Центральная часть трансляции называется элонгацией, окончание – терминацией. Внутри рибосомы находятся 2 кодона и-РНК: в аминоацильном и в пептидиальном центрах. Т-РНК с аминокислотой-1 подходит к аминоацильному центру (стадия инициации) и если антикодон т-РНК комплементарен кодону и-РНК, то происходит присоединение т-РНК с аминокислотой-1 к триплету и-РНК.
Затем рибосома передвигается на 1 кодон и-РНК, и т-РНК с аминокислотой-1 перемещается в пептидильный центр, а освободившемуся аминоацильному центру подходит т-РНК с аминокислотой-2 и устанавливается там, если антикодон т-РНК комплементарен кодону и-РНК. С помощью ферментов между аминокислотой-1 и аминокислотой-2, находящимися в рибосоме, устанавливается пептидная связь.
Одновременно разрушается связь между аминокислотой -1 и ее т-РНК, и т-РНК уходит из рибосомы за следующей аминокислотой. Рибосома опять перемещается на 1 триплет и процесс повторяется. Так наращивается полипептид (стадия элонгации), в котором аминокислота располагаются в соответствии спорядкам кодирующих их триплетов.

Рис. 4. Схема биосинтеза белка.
Окончание синтеза (стадия терминации) происходит, когда в аминоацильном центре появляется терминирующий кодон. После высвобождения синтезированного пептида и т-РНК, рибосома диссоциирует на субъединицы, готовые для трансляции новых и-РНК.
4.Регуляция транскрипции у прокариот была изучена в 1961 году с М.Жакоб, Ж. Моно и А. Львовым. Единица регуляции транскрипции у прокариотических организмов – оперон, в состав которого входят:

1. Промотор – место прикрепления РНК – полимеразы.
2. Ген – оператор – регулирует доступ РНК – полимеразы к структурным генам, взаимодействуя с регуляторными белками.
3. Инициатор – место начала считывания генетической информации.
4.Структурные гены - определяют синтез белков – ферментов, обеспечивающие цепь последовательных биохимических реакций.
5. Терминатор – последовательность нуклеотидов завершающая транскрипцию.
Ген – регулятор расположен вблизи оперона, он постоянно активен, на основе его информации синтезируется белок – репрессор.
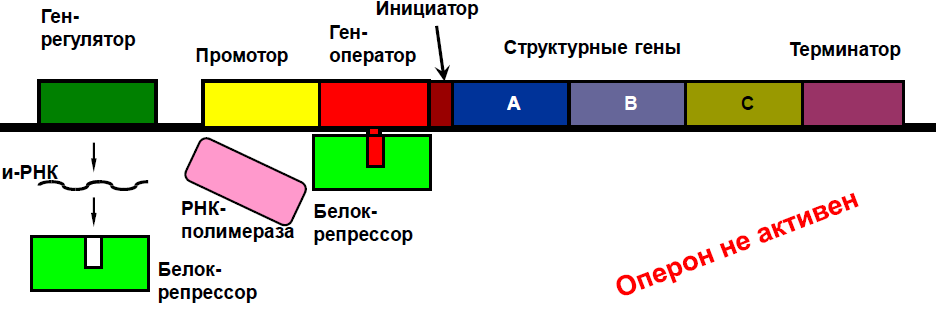
Белок репрессор образует химическое соединение с геном – оператором, и препятствует соединению РНК – полимеразы с промотором.
Главный механизм регуляции активности оперона – индукция.

Регуляции транскрипции у эукариот была изучена Г.П. Георгиевым в 1972 году.
Единица транскрипции – транскриптон, состоящей из неинформативной (акцепторной) и информативной (структурной) зон.
Неинформативная зона: промотор, инициатор, регуляторные последовательности.
Информативная зона: структурный ген, имеющий мозаичную экзон – интронную структуру. Интроны – вставки из неинформативных участков ДНК. Экзоны – последовательности ДНК, содержащие информацию о структуре полипептида. Заканчивается транскриптон терминатором.

Особенности регуляции экспрессии генов эукариот:
1. Работу транскриптона контролирует несколько генов – регуляторов, дающие информацию для синтеза регуляторных белков и факторов транскрипции.
2. Для включения транскриптона необходимо множество регулирующих компонентов, необходимых для сборки транскрипционного комплекса.
3. Первичныйтранскрипт (про-и-РНК) содержит информацию об экзонах и интронах. Для его превращения в и-РНК необходим процесс созревания.
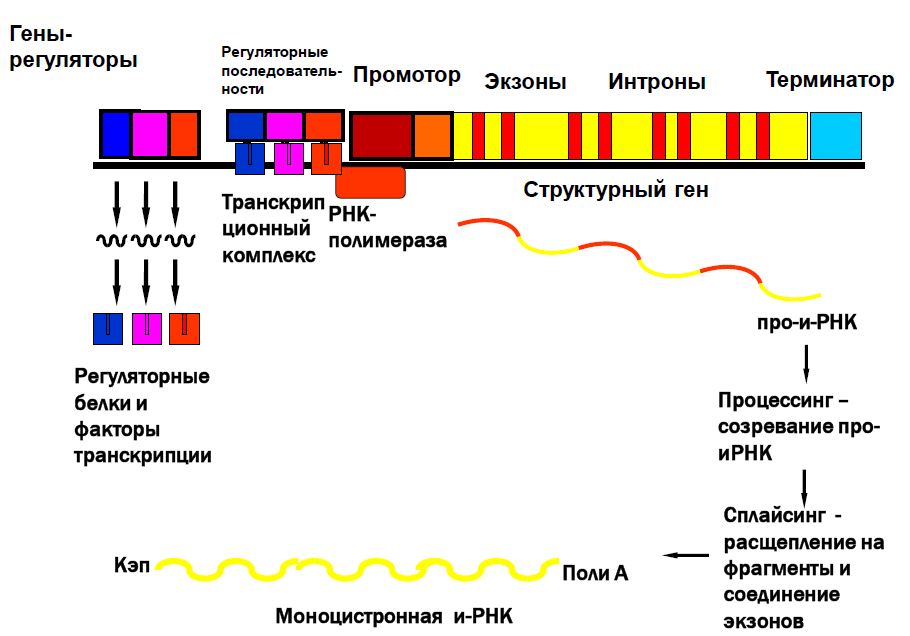
4. Процессинг – модификация концов про-и-РНК и сплайсинг.
5. Кэпирование на 5′- концеиполиаденилированиена 3′- конце. Кэп («шапочка»изтрифосфометилгуанозина) и полиадениловый «хвост» защищают и-РНК от действия нуклеаз.
6. Сплайсинг – вырезание интронов и стыковка экзонов.
7. Сплайсинг осуществляет сложный комплекс мя-РНП и белков, называемых сплайсосомой.
8. Образованная и-РНК является моноцистронной.
9. Альтернативныйсплайсинг – в результате процессинга одного и того же первичного транскрипта, могут образовываться разные и-РНК, и как следствие, синтезироваться разные полипептиды.
5. Основная генетическая информация организма содержится в клеточном ядре. В 1908 г. К. Корренс описал внеядерную (цитоплазматическую) наследственность. Генетический материал содержат митохондрии и пластиды. Эти единицы в отличие от ядерных генов, называются плазмогенами. В цитоплазме клеток может находиться ДНК вирусов и плазмиды бактерий – кольцевые двухцепочечные ДНК.
У человека с цитоплазматической наследственностью связаны болезнь Лебера (нейрит с атрофией зрительного нерва) и анэнцефалия.
Цитоплазматическое наследование идет по материнской линии, через яйцеклетки, так как сперматозоиды практически не содержат цитоплазмы.
Критериями цитоплазматической наследственности являются:
- отсутствие расщепления признаков в потомстве по законам Менделя;
- невозможность выявить группы сцепления;
- различные результаты возвратного скрещивания;
Известны несколько видов цитоплазматической наследственности.
Митохондриальная наследственность описана Б. Эфрусси в 1949 г. Он обнаружил, что примерно 1% колоний хлебных дрожжей образуют карликовые колонии. Их рост тормозится потому, что произошла мутация плазмогенов и их митохондрии не имеет дыхательных ферментов. Геном митохондрий человека представлен кольцевой молекулой ДНК, содержащей 16569 пар нуклеотидов. В митохондриальной ДНК имеется очень мало некодирующихучастков и транскрибируются обе ее цепочки. Имеются данные о некоторых болезнях человека, которые являются следствием мутаций митохондриальных генов (например: митохондриальнаяцитопатия, несращение верхних дуг позвонков, старческое слабоумие, паркинсонизм и др.).
Пластидную наследственность описал К. Корренс в 1908 году. Растение ночная красавица имеет пестрые листья. Произошла мутация, и в части пластид не образуется хлорофилл. Пластиды при размножении распределяются неравномерно. Часть клеток получает нормальные пластиды и имеет зеленые листья; часть клеток получает пластиды, не имеющие хлорофилла – листья белые и растение погибает; часть клеток получает и зеленые (нормальные) и мутантные пластиды – растения имеют пестрые листья (зеленые с белыми пятнами).
Псевдоцитоплазматическая наследственность связана с попаданием в клетку вируса или чужеродной (бактериальной) ДНК. Примером может быть предрасположенность некоторых мышей к опухолям молочной железы. Если нормальных мышат кормит самка «раковой линии», все мыши будут иметь опухоли молочной железы, и наоборот: если мышат «раковой линии» кормит здоровая самка, все мышата будут здоровы. Причиной фактора молока у мышей оказался вирус. Вторым примером может быть гибель XY-зигот дрозофил, которую вызывает спирохета, попадающая в мужские гаметы.