

Нормальная физиология (Пособие для резидентуры)-1
.pdfАНАЛИЗАТОРЫ 16
лизация о прикосновении, давлении и движениях в суставах. Большинство нейронов вентробазального ядра таламуса возбуждается при механической стимуляции только их рецептивного поля; каждый нейрон возбуждается только одним типом рецепторов – например, медленно адаптирующимися рецепторами давления или рецепторами волосяных луковиц.
Вентробазальное ядро таламуса связано восходящими и нисходящими аксонами с первой и второй соматосенсорными зонами (S1-S2) коры больших полушарий головного мозга. Соматосенсорная кора представлена главным образом постцентральной извилиной. Она построена из функциональных единиц, представляющих собой нейронные колонки, ориентированные перпендикулярно к ее поверхности; каждая колонка имеет диаметр 0,2-0,5 мм. Нейроны каждой колонки функционально связаны только с рецепторами одного типа. Таким образом, колонки специализированы не только в отношении локализации, но и в отношении типа рецепторов. Каждая колонка соматосенсорной коры мозга благодаря большому числу содержащихся в ней нейронов (до 105) обладает значительными возможностями для обработки информации, поступающей с периферии. Среди нейронов в колонке существует своего рода иерархия, основанная на различных возбудительных и тормозных взаимодействиях между ними.
Первичные и вторичные сенсомоторные области в симметричных отделах коры мозга связаны между собой через мозолистые тела. В этих областях выявлено соматотопическое различие нейронов. Так, нейроны поля 3а по Бродману получают информацию от мышечных и суставных рецепторов, нейроны поля 2 возбуждаются при сдавливании кожи и практически не реагируют на легкое прикосновение. Нейроны поля 3б реагируют на легкое прикосновение к коже и сгибание отдельных волосков. Нейроны поля 1 реагируют как на легкое раздражение кожи, так и на ее сдавливание.
16.7. Мышечно-суставнаярецепция(проприорецепция)
Необходимым условием мышечной деятельности является получение информации о положении тела в пространстве и о степени сокращения каждой из мышц. Эта информация поступает в ЦНС от рецепторов вестибулярного аппарата, глаз, кожи, а также от мышечносуставных рецепторов (проприорецепторов).
Основные типы проприорецепторов – это механорецепторы, расположенные в суставных сумках, мышечные веретена, сухожильные органы Гольджи и кожные механорецепторы.
В суставных сумках заложено несколько видов рецепторов:
 мелкие тельца, похожие на тельца Руффини. Они реагируют на степень растяжения суставной сумки;
мелкие тельца, похожие на тельца Руффини. Они реагируют на степень растяжения суставной сумки;
 крупные быстро адаптирующиеся тельца типа телец Пачини;
крупные быстро адаптирующиеся тельца типа телец Пачини;
 сухожильные органы Гольджи;
сухожильные органы Гольджи;
 свободные окончания нервных волокон.
свободные окончания нервных волокон.
Суставные рецепторы изучены меньше, чем мышечные. Известно, что они реагируют на положение сустава и на изменения суставного угла, участвуя таким образом в системе обратных связей от двигательного аппарата /3/.
461
НОРМАЛЬНАЯ ФИЗИОЛОГИЯ
Вкаждой мышце есть волокна, которые короче и тоньше остальных. Они располагаются
ввиде небольших скоплений, окруженных соединительной капсулой, и имеют форму веретена – это мышечное веретено. В них имеются два отдела: мышечные волокна и ядерная сумка. Заключенные в капсулу мышечные волокна называются интрафузальными. Существуют два типа интрафузальных волокон: волокна с ядерной цепочкой (ядерно-цепочечные),
вкоторых клеточные ядра располагаются цепочкой, вдоль мышечного волокна, и волокна с ядерной сумкой (ядерно-сумчатые), в которых клеточные ядра располагаются ближе к центру
ввиде скопления, образуя выпуклость. Вокруг центральной части интрафузальных волокон спирально расположены терминали дендритов (афферентные нервные окончания), а эфферентные окончания (терминали аксонов) окружают периферические участки интрафузальных и экстрафузальных мышечных волокон (рис. 16.28).
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Нисходящие волокна от |
|||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Мышечное |
|
|
высших моторных центров |
|||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
веретено |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Задний рог |
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
7 |
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Iα-волокна |
|
|
|
|
Задний корешок |
||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||
8 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||
|
|
|
|
|
|
4 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
5 |
Мышца |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
6 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
3 |
|
|
|
|
|
|
|
|
|
Передний корешок |
||||||||||
|
|
|
|
|
|
|
|
|
|
|
α-Волокно |
|
|
|
||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Передний рог |
|
|||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||
|
|
|
1 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
γ-Волокно |
||||||||||||||
|
|
|
|
|
2 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
γ-Мотонейрон |
|
||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
α-Мотонейрон |
|
||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||
АБ
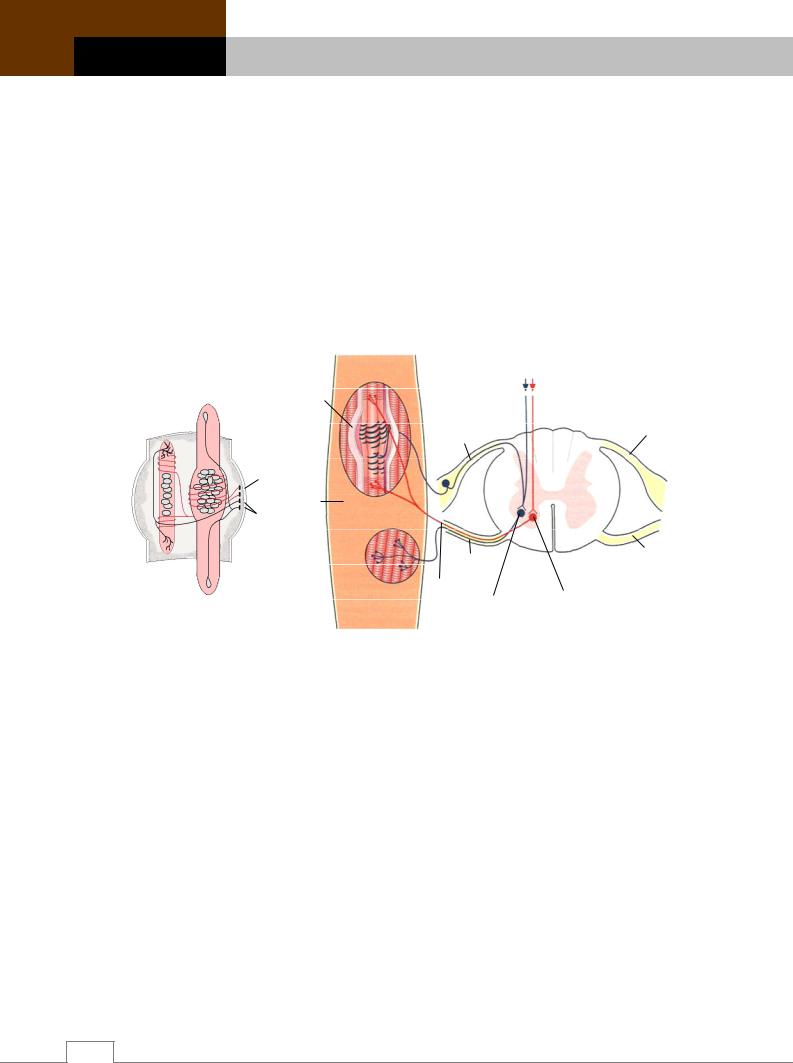
Рис. 16.28. Моторные элементы сегментов спинного мозга и мышечное веретено.
А – схема: 1 – интрафузальное мышечное волокно с ядрами, расположенными цепочкой; 2 – интрафузальное мышечное волокно с ядрами, расположенными в ядерной сумке;
3 – соединительнотканная капсула мышечного веретена; 4 – афферентное нервное волокно типа Ia; 5 – афферентное нервное волокна типа IIа; 6 – эфферентные гамма-нервные волокна; 7 – гамма-концевая пластинка; 8 – гамма-кустовидное нервное окончание. Б – строение и функция.
Мышечное веретено состоит из 6-10 интрафузальных мышечных волокон. В отличие от интрафузальных, обычные сократительные волокна называют экстрафузальными. Концы мышечных волокон веретен прикрепляются к двум соседним экстрафузальным волокнам с помощью сухожильных полосок, благодаря чему они расположены параллельно экстрафузальным мышечным волокнам. Поэтому при растяжении мышцы нагрузка на веретена увеличивается, а при сокращении уменьшается. Веретена снабжены двумя видами нервных окончаний: афферентные – окружающие ядерную сумку (типа Iа), и моторные (эфферентные) – иннервирующие интрафузальные мышцы и идущие от -мотонейронов спинного мозга. Скелетная мышца может содержать до 500 мышечных веретен, собранных в пучки и окруженных капсулой.
462

АНАЛИЗАТОРЫ 16
Сухожильные органы Гольджи располагаются в сухожилиях с обеих концов мышцы. Сухожильные рецепторы слабо реагируют на растяжение мышцы, но возбуждаются при ее сокращении. Они являются как бы источником информации о силе, развиваемой мышцей. К ним подходят 1-2 толстых миелинизированных афферентных нервных волокна группы Ib. На спинальном уровне они через интернейроны вызывают торможение мотонейронов собственной мышцы и возбуждение мотонейронов мышцы-антагониста /3/.
Оба типа проприорецепторов являются рецепторами растяжения. По волокнам Iа, отходящим от мышечных веретен импульсы поступают в передние рога спинного мозга, а отсюда в головной мозг, что обеспечивает информацию о длине мышцы. Пришедшие в спинной мозг афференты от мышечных веретен запускают рефлекс сокращения данной мышцы, так как их нервные окончания образуют возбуждающий синапс с α-мотонейронами данной мышцы. Мышечное веретено является рецептором, воспринимающим степень растяжения мышцы.
Веретена имеют и эфферентную иннервацию – интрафузальные волокна иннервируются аксонами -мотонейронов. Активация -мотонейронов приводит к повышению чувствительности веретен на длину и скорость удлинения мышцы.
Сокращаясь, мышца растягивает сухожилие. Это вызывает возбуждение нервных окончаний чувствительных нейронов спинного мозга. Они возбуждают тормозные нейроны спинного мозга, которые тормозят соответствующие двигательные нейроны, что предотвращает перерастяжения мышцы. Известно, что порог возбуждения рецепторов сухожильного органа Гольджи при механическом растяжении мышцы выше, чем аналогичный порог возбуждения мышечных веретен. Это позволяет мышце сокращаться без всяких помех до определенного предела.
Сигналы от сухожильного органа передаются через крупные быстропроводящие нервные волокна типа lb со средним диаметром 16 мкм, который лишь немного меньше диаметра волокон от первичных окончаний мышечного веретена. Как и волокна от первичных окончаний веретен, быстропроводящие нервные волокна типа lb передают сигналы в локальные области спинного мозга, а также, после синаптического переключения в заднем роге, через пути длинных волокон (например, спиноцеребеллярные тракты) в мозжечок, а через другие тракты – к коре большого мозга. Местный сигнал в спинном мозге возбуждает одиночный тормозной вставочный нейрон, который тормозит передний мотонейрон. Сухожильные органы Гольджи отслеживают степень напряжения мышцы.
Проводниковый и корковый отделы двигательного анализатора. Афферентная информация от мышечных, сухожильных и суставных рецепторов в составе задних корешков поступает через аксоны афферентных нейронов, находящихся в спинномозговых ганглиях, в спинной мозг, где частично переключается на α-мотонейроны или на вставочные нейроны (например, на клетки Реншоу), а частично направляется по восходящим путям в высшие отделы головного мозга. В частности, по спинно-мозжечковому пути проприоцептивная импульсация доставляется к мозжечку, а по пучкам Голля и Бурдаха, проходящим в дорсальных канатиках спинного мозга, она доходит до нейронов одноименных ядер, расположенных в продолговатом мозге. Расположенные там сенсорные нейроны второго порядка образуют мощный бульботаламический пучок, или медиальный лемниск, который заканчивается в задневентральных (специфических) ядрах таламуса. Аксоны таламических
463

НОРМАЛЬНАЯ ФИЗИОЛОГИЯ
нейронов (нейронов третьего порядка) оканчиваются в коре больших полушарий, главным образом, в соматосенсорной зоне коры (постцентральная извилина) и в области сильвиевой борозды, а также частично в двигательной области коры.
Регуляция активного мышечного тонуса. Высшие моторные центры в головном мозге через так называемые экстрапирамидные пути (красноядерно-покрышечно-преддверно-рети- кулярное и луковично-ретикулярное спинальные пути) активируют γ-мотонейроны и через мышечные веретена регулируют длину мышц и, следовательно, активный мышечный тонус. Активация γ-мотонейронов вызывает напряжение сократительных концов интрафузальных волокон, что растягивает центр мышечных веретен и приводит к активации Ia-волокон. Как уже было отмечено, импульс затем передается через задние рога в передние рога спинного мозга и возбуждает α-мотонейроны. α-Мотонейроны, в свою очередь, вызывают сокращение волокон скелетных мышц, что опосредованно уменьшает натяжение центральной части веретен. Эта рефлекторная реакция продолжается до тех пор, пока Iа-волокна не будут регистрировать натяжение (γ-петля). Активация красноядерно-спинномозговых влияний приводит к возбуждению - и -мотонейронов сгибательной и торможению соответствующих мотонейронов разгибателей. Нервные импульсы, идущие по преддверно-спинномозго- вому пути, вызывают обратные эффекты: активацию - и -мотонейронов разгибателей и торможение соответствующих мотонейронов, иннервирующих мышцы-сгибатели.
При нормальной позе (сидя, стоя или при ходьбе) в большинстве мышц, особенно в работающих против силы тяжести, регистрируется постоянная невысокая активность афферентных волокон мышечных веретен, что позволяет поддерживать определенный активный мышечный тонус. Мышцы, действующие против силы тяжести, например мышца, разгибающая позвоночник, в положении стоя находятся в постоянном небольшом натяжении, так как туловище под влиянием силы тяжести наклоняется несколько кпереди. За счет постоянной активации γ-мотонейронов достигается дополнительное растяжение средней части мышечных веретен, что вызывает активацию высших моторных центров для поддержания мышечного тонуса. Благодаря постоянной фоновой афферентной импульсации во время обычных движений мышечные веретена могут воспринимать как укорочение, так и растяжение мышцы. При усилении или уменьшении активности γ-мотонейронов изменяется чувствительность мышечных веретен и регулируется их активность при конкретных позе и движении. Таким образом, γ-мотонейроны устанавливают в каждой конкретной ситуации оптимальную рабочую область мышечных веретен, и они могут регистрировать мышечную длину и скорость ее изменения. Это позволяет устанавливать и поддерживать постоянный активный мышечный тонус, а следовательно, поддерживать равновесие и положение суставов и определенную позу (чувство позы и опоры).
Выполнение точных движений и регуляция тонкой моторики. Возможность регулировать чувствительность мышечных веретен, особенно вместе с произвольными движениями, играет большую роль. При произвольных движениях, например при разгибании в коленном суставе, происходит, помимо активации α-мотонейронов, и повышение активности γ-мотонейронов. За счет α-γ-коактивации устанавливается оптимальная длина мышечных волокон, чтобы поддерживать их чувствительность во время всего движения. При сокращении регистрируется растяжение мышцы и становятся возможными очень точные движения. Супраспинальные нервные центры могут изменять γ-активность в γ-петле
464

АНАЛИЗАТОРЫ 16
(изменение частоты импульсации по Iа-волокнам) и таким образом изменять активность связанных с ними α-мотонейронов.
Передача и переработка соматосенсорной информации. Чувствительность кожи и ощущение движения обусловлены проведением в мозг сигналов от рецепторов по двум основным путям (трактам): лемнисковому и спинноталамическому, значительно различающимся по своим морфологическим и функциональным свойствам. Третий путь – латеральный тракт Морина близок по ряду характеристик к лемнисковой системе.
Лемнисковый путь. Этот путь состоит из относительно толстых и быстропроводящих миелинизированных нервных волокон. Он передает в мозг сигналы о прикосновении к коже, давлении на нее и движениях в суставах. Отличительная особенность этого пути заключается
вбыстрой передаче в мозг наиболее точной информации, дифференцированной по силе и месту воздействия. Первые нейроны этого пути находятся в спинномозговом узле, их аксоны
всоставе задних столбов восходят к тонкому (ядро Голля) и клиновидному (ядро Бурдаха) ядрам продолговатого мозга, где сигналы передаются на вторые нейроны лемнискового пути. Часть волокон, в основном несущих сигналы от суставных рецепторов, оканчивается на мотонейронах сегментарного спинального уровня. Проприоцептивная чувствительность передается в спинном мозге также по дорсальному спинно-мозжечковому, спинно-церви- кальному и некоторым другим путям.
Впродолговатом мозге в тонком ядре сосредоточены в основном вторые нейроны тактильной чувствительности, а в клиновидном ядре вторые нейроны проприоцептивной чувствительности. Аксоны этих нейронов образуют медиальную петлю и после перекреста на уровне олив направляются в специфические ядра таламуса – вентробазальный ядерный комплекс. В этих ядрах концентрируются третьи нейроны лемнискового пути. Их аксоны направляются в соматосенсорную зону коры большого мозга.
По мере перехода на все более высокие уровни изменяются некоторые важные свойства нейронов лемнискового пути. Значительно увеличиваются размеры рецептивных полей нейронов. Ответы клеток становятся все более продолжительными: даже короткое прикосновение к коже вызывает залп импульсов, длящийся несколько секунд. Отмечено появление так называемых нейронов новизны, реагирующих на смену раздражителя. Несмотря на увеличение размеров рецептивных полей, нейроны остаются достаточно специфичными (нейроны поверхностного прикосновения, глубокого прикосновения, нейроны движения в суставах и нейроны положения или угла сгибания суставов). Для корковой части лемнискового пути характерна четкая топографическая организация, т.е. проекция кожной поверхности осуществляется в кору большого мозга по принципу «точка в точку». При этом площадь коркового представительства той или иной части тела определяется ее функциональной значимостью – формируется так называемый сенсорный гомункулю.
Удаление соматосенсорной зоны коры приводит к нарушению способности локализовать тактильные ощущения, а ее электростимуляция вызывает ощущение прикосновения, вибрации и зуда. В целом роль соматосенсорной зоны коры состоит в интегральной оценке соматосенсорных сигналов, во включении их в сферу сознания, полисенсорный синтез и в сенсорное обеспечение выработки новых двигательных навыков.
Спинно-таламический путъ.Этот путь значительно отличается от лемнискового. Его первые нейроны также расположены в спинномозговом узле, откуда они посылают в
465

НОРМАЛЬНАЯ ФИЗИОЛОГИЯ
спинной мозг медленнопроводящие немиелинизированные нервные волокна. Эти нейроны имеют большие нечеткие рецептивные поля, иногда включающие значительную часть кожной поверхности. Вторые нейроны данного пути локализуются в сером веществе спинного мозга, а их аксоны в составе восходящего спинно-таламического пути направляются после перекреста на спинальном уровне в вентробазальный ядерный комплекс таламуса (дифференцированные проекции), а также в вентральные неспецифические ядра таламуса, внутреннее коленчатое тело, ядра ствола мозга и гипоталамус. Локализованные в этих ядрах третьи нейроны спинно-таламического пути лишь частично дают проекции в соматосенсорную зону коры.
Спинно-таламический путь с более медленной передачей афферентных сигналов, со значительно менее четко дифференцированной информацией о разных свойствах раздражителя и с менее четкой топографической локализацией служит для передачи температурной, всей болевой и в значительной мере – тактильной чувствительности.
Болевая чувствительность практически не представлена на корковом уровне (раздражение коры большого мозга не вызывает боли), поэтому считают, что высшим центром болевой чувствительности является таламус, где 60% нейронов в соответствующих ядрах, четко реагирует на болевое раздражение. Таким образом, эта система играет важную роль в организации генерализованных ответов на действие болевых, температурных и тактильных раздражителей, сигналы о которых идут через структуры ствола, подкорковые образования и кору большого мозга.
Обонятельнаясистема
Рецепторы обонятельной системы расположены в области верхних носовых ходов. Обонятельный эпителий имеет толщину 100-150 мкм и содержит около 10 млн рецепторных клеток диаметром 5-10 мкм, расположенных между опорными клетками (рис. 16.29) /3/.
Обонятельная рецепторная клетка – первичная биполярная клетка, на поверхности которой имеется сферическое утолщение – обонятельная булава, из которой выступает 6-12 волосков длиной до 10 мкм. Булава является важным цитохимическим центром обонятельной клетки. Обонятельные волоски погружены в жидкую среду, вырабатываемую боуменовыми железами. Наличие подобных волосков в десятки раз увеличивает площадь контакта рецептора с молекулами пахучих веществ. От нижней части рецепторной клетки отходит немиелинизированный аксон. Аксоны всех рецепторов образуют обонятельный нерв, который проходит через основание черепа и вступает в обонятельную луковицу. Обонятельные клетки постоянно обновляются. Продолжительность жизни обонятельных клетки около 2 месяцев.
Молекулы пахучих веществ попадают в обонятельную слизь с постоянным током воздуха. Здесь они взаимодействуют с находящимся в волосках рецептора рецепторным белком, который активирует, как и в случае фоторецепции, ГТФ-связывающий белок (G-белок), а тот в свою очередь фермент аденилатциклазу, синтезирующую цАМФ. В результате этого в мембране рецептора открываются натриевые каналы, и генерируется рецепторный потенциал. Это приводит к импульсному разряду в аксоне рецептора – волокне обонятельного нерва.
466




НОРМАЛЬНАЯ ФИЗИОЛОГИЯ
ГЛАВА17. ВЫСШАЯНЕРВНАЯДЕЯТЕЛЬНОСТЬ
17.1. УчениеИ.П.Павловаовысшейнервнойдеятельности
Полушария большого мозга – их кора и ближайшие к ней подкорковые образования – являются высшим отделом ЦНС позвоночных животных и человека. Функция этого отдела – осуществление сложных рефлекторных реакций, составляющих основу высшей нервной деятельности (поведения) организма.
Впервые представление о рефлекторном характере деятельности высших отделов головного мозга было широко и подробно развито основоположником отечественной физиологии И.М.Сеченовым в его книге «Рефлексы головного мозга».
Идеи И.М.Сеченова получили блестящее развитие в замечательных трудах И.П.Павлова, открывшего пути объективного экспериментального исследования функций коры большого мозга, разработавшего метод условных рефлексов и создавшего учение о высшей нервной деятельности.
И.П.Павлов показал, что в то время как в нижележащих отделах ЦНС – подкорковых ядрах, мозговом стволе, спинном мозге – рефлекторные реакции осуществляются врожденными, наследственно закрепленными нервными путями, в коре большого мозга нервные связи вырабатываются заново в процессе индивидуальной жизни животных и человека в результате сочетания бесчисленных, действующих на организм и воспринимаемых корой раздражений.
Открытие этого факта позволило разделить всю совокупность рефлекторных реакций, происходящих в организме, на две основные группы – безусловные и условные рефлексы.
Различиябезусловныхиусловныхрефлексов
 Безусловные рефлексы по И.П.Павлову – наследственно передающиеся реакции, т.е. генетически детерминированы, врожденные. Безусловные рефлексы возникают на основе врожденных рефлекторных дуг. Причем большинство из них начинают функционировать сразу же после рождения.
Безусловные рефлексы по И.П.Павлову – наследственно передающиеся реакции, т.е. генетически детерминированы, врожденные. Безусловные рефлексы возникают на основе врожденных рефлекторных дуг. Причем большинство из них начинают функционировать сразу же после рождения.
Условные рефлексы – реакции приобретенные в процессе индивидуальной жизни. Биологическое значение условных рефлексов заключается в резком расширении числа сигнальных, значимых для организма раздражителей, что обеспечивает более высокий уровень адаптивного поведения. Условно-рефлекторный механизм лежит в основе формирования любого приобретенного навыка, в основе процесса обучения.
 Безусловные рефлексы являются видовыми, т.е. эти рефлексы свойственны всем представителям данного вида. Условные рефлексы – индивидуальны, у одних представителей одного и того же вида они могут быть, у других отсутствовать;
Безусловные рефлексы являются видовыми, т.е. эти рефлексы свойственны всем представителям данного вида. Условные рефлексы – индивидуальны, у одних представителей одного и того же вида они могут быть, у других отсутствовать;
470