

2 курс / Нормальная физиология / Общий_курс_физиологии_человека_и_животных_Том_2_Ноздрачев_А_Д_,
.pdfпоявлению на ЭКГ интегрального вектора — направленного вниз зубца Q.
Зубец R является самым высоким в ЭКГ. Он представляет собой период распространения возбуждения по основаниям желудочков, в то время как зубец S отражает полный охват возбуждением желудочков, когда вся поверхность сердца стала электроотрицательной и исчезла, таким образом, разность потенциалов между отдельными участками сердца. Комплекс QRS совпадает с реполяризацией предсердий. Его длительность составляет 0,06—0,09 с.
Зубец Т отражает восстановление нормального потенциала мембраны клеток миокарда, т. е. реполяризации миокарда. Этот зубец является самой изменчивой частью ЭКГ, так как реполяризация происходит не одновременно в разных волокнах миокарда. Сегмент ТР совпадает с периодом покоя сердца — общей паузой и диастолой. Общая длительность комплекса QRST равна примерно 0,36 с. Электрокардиограмма и основные показатели деятельности сердца в различные фазы сердечного цикла приведены на рис. 8.15.
Рис. 8.15. Процессы, соответстующие определенным фазам сердечного цикла: I — давление в аорте, II
— давление в желудочке, III — давление в предсердии, IV — изменение объема желудочков, V — венный пульс; VI — электрокардиограмма; VII — тоны сердца; 1 — закрытие атриовентрикулярных клапанов и начало сокращения желудочков, 2 — раскрытие аортальных клапанов; 2—3 — фаза быстрого изгнания, 3— 4 — фаза медленного изгнания, 4—5 — протодиастолическая фаза, 6 — закрытие аортальных клапанов, 6—7 — быстрое наполнение; от отметки 7 до начала систолы предсердий — поздняя диастолическая фаза; отметка времени на кривой V (0,1 с) соответствует всем трем нижним кривым
Относительная продолжительность отдельных фаз ЭКГ млекопитающих имеет сходный характер, несмотря на то, что ритм сердца варьирует у них в очень широких пределах. Например, у мыши при частоте сердечных сокращений 635 в 1 мин интервал PR составляет 57 %, QT — 42 %, у крокодила при частоте сердечных сокращений 36 в 1 мин соответственно PR — 30 %, QT — 70 %. Во время зимней спячки, например, у ежа сердце сокращается в редком ритме, поэтому интервал PR будет несравненно длиннее других компонентов ЭКГ.
В разных областях сердца во время сердечного цикла процессы деполяризации и реполяризации возникают не одновременно. В связи с этим колеблется и разность потенциалов между ними. Условную линию, соединяющую две точки ЭКГ с наибольшей разностью потенциалов, называют электрической осью сердца. В отдельные периоды она характеризуется разной величиной и направленностью, т. е. обладает свойством векторной величины. Одновременная регистрация величины разности потенциалов и характера электрической оси сердца называется вектор-кардиограммой.
У беспозвоночных электрограмма нейрогенного сердца состоит из быстрых осцилляции
и медленных колебаний. Эти разряды соответствуют потенциалам пейсмекерных ганглиозных клеток. Их удается зарегистрировать от одиночных нейронов. У членистоногих форма и продолжительность аналогичных разрядов зависят от многих факторов таких, например, как температура.
У ракообразных в сердечных ганглиях небольшое число нервных клеток образует единую систему, обладающую ритмической активностью. Клетки эти мелкие и крупные. Мелкие являются пейсмекерами, крупные — моторными нейронами, воспроизводящими ритм пейсмекеров. От пейсмекерных нейронов регистрируются медленные потенциалы. Моторные клетки разряжаются частыми импульсами в виде залпов. Они не влияют на пейсмекерные клетки своими разрядами, но могут оказывать электротоническое действие. Характер взаимоотношений между ними предполагает наличие реверберирующих контуров.
8.4. Регуляция работы сердца
Относительное постоянство параметров внутренней среды организма, быстрое и точное приспособление гемодинамики к широкому разнообразию условий, в которых находится организм, достигаются благодаря в высшей степени совершенным механизмам регуляции сердечной деятельности. У млекопитающих, птиц, рептилий и амфибий эта регуляция осуществляется принципиально одинаково и складывается из присущих самому сердцу собственных механизмов, а также внесердечных воздействий. К внутрисердечным регуляторным механизмам относят внутриклеточные, регуляцию межклеточных взаимодействий и собственно внутрисердечные нервные механизмы. Внесердечные воздействия представлены нервной и гуморальной регуляцией.
8.4.1. Внутриклеточная регуляция
Внутриклеточный уровень регуляции заключается в способности кардиомиоцитов при выполнении ими специфической сократительной функции синтезировать различные белки в соответствии с уровнем их разрушения. Синтез белков происходит дифференцированно олагодаря существованию специальных ауторегуляторных механизмов. Процесс этот осуществляется кардиомиоцитами во взаимодействии с соединительно-тканными клетками.
Особенность кардиомиоцитов заключается в цикличности их обменных процессов, связанных с ритмом сердечной деятельности. Наиболее быстрый распад богатых энергией соединений — АТФ и гликогена — происходит в момент систолы и соответствует комплексу QRS электрокардиограммы. Ресинтез и восстановление уровня этих веществ успевает полностью осуществиться за время диастолы. Поэтому в чрезвычайных условиях при усиленной работе сердца одним из компенсаторных механизмов, адаптирующих деятельность сердца к воздействиям, является удлинение фазы диастолы.
Кардиомиоциты способны избирательно адсорбировать из циркулирующей крови и накапливать в цитоплазме вещества, поддерживающие и регулирующие их биоэнергетику, а также соединения, повышающие потребность клеток в кислороде. Последнее имеет большое приспособительное значение, так как эти влияния посредством гомеометрического механизма регуляции сократительной силы сердца вызывают увеличение силы его сокращений. Так, возникающая благодаря внутриклеточным механизмам адсорбция из крови катехоламинов (адреналина, норадреналина) обеспечивает усиление сердечного кровотока, что сопровождается более сильным растяжением клеток миокарда во время диастолы. (Известно, что чем больше растянута миофибрилла, тем сильнее она способна к сокращению.) Следовательно, чем больше крови притекло к сердцу, тем сильнее оно растянуто во время диастолы и тем интенсивнее его сокращение. Этот механизм лежит в основе закона Франка — Старлинга (см. разд.
8.2.2).
8.4.2.Межклеточная регуляция
Всердечной мышце межклеточная регуляция связана с наличием вставочных дисков — нексусов, обеспечивающих транспорт неооходимых веществ, соединение миофибрилл, переход возбуждения с клетки на клетку. Такая организация позволяет миокарду реагировать на возбуждение как синцитий. В случае выпадения функции нексусов нарушается одновременность прихода возбуждения к отдельным клеткам, синхронность сокращения мышечных волокон, появляется нарушение сердечного ритма. Межклеточная регуляция включает также взаимодействие кардиомиоцитов с соединительно-тканными клетками, составляющими строму сердечной мышцы. Наряду с механической опорной функцией соединительнотканные клетки являются источником пополнения кардиомиоцитов высокомолекулярными органическими соединениями, постоянно необходимыми как для функции, так и для поддержания структуры клетки.
8.4.3.Внутрисердечная нервная регуляция
Внутрисердечный уровень регуляции является автономным, хотя он включен и в сложную иерархию центральной нервной регуляции. Собственная нервная регуляция сердца осуществляется метасимпатической нервной системой, нейроны которой располагаются в интрамуральных ганглиях сердца (см. разд. 5.4.3). Метасимпатическая нервная система обладает полным набором функциональных элементов, необходимых для самостоятельной рефлекторной деятельности: сенсорными клетками, интегрирующим интернейронным аппаратом, двигательными нейронами.
Сенсорные нейроны обслуживают не только внутрисердечные механизмы регуляции. По их аксонам, проходящим в составе блуждающих и симпатических нервов, чувствительная импульсация может достигать высших отделов нервной системы. В свою очередь, на вставочных и моторных метасимпатических нейронах синаптически оканчиваются преганглионарные волокна блуждающего нерва и сердечных симпатических ветвей, т. е. метасимпатические нейроны — общий конечный путь для импульсов внутрисердечного и центрального происхождения
Вдеятельности метасимпатической нервной системы участвует более 20 известных сейчас медиаторов и модуляторов межнейронной передачи возбуждения. Долгое время считали, что внутрисердечный нервный аппарат является лишь местом переключения центробежных импульсов с преганглионарных волокон блуждающего нерва.
Интракардиальный метасимпатический нервный аппарат регулирует ритм сердечных сокращений, скорость предсердно-желудочкового проведения, реполяризацию кардиомиоцитов, скорость диастолического расслабления.
Функции сердечной части метасимпатической нервной системы особенно отчетливо выступают при трансплантациях сердца у теплокровных животных. После дегенерации всех нервных волокон внесердечного происхождения (симпатических и парасимпатических) она остается неповрежденной в структурном и функциональном отношении. При разных нагрузках на организм с гомотрансплантированным сердцем кровообращение в большом круге меняется так же, как в нормальных условиях, и целиком обеспечивает потребности организма.
Вспециальных опытах при гемодинамическом разобщении правой и левой половин сердца Г. И. Косицким установлено, что растяжение механорецепторов одной из камер изолированного сердца сопровождается усилением сокращения не только этой камеры, но
идругих камер, диастолическая длина мышечных волокон которых не изменилась. Эти реакции других камер в отличие от растягиваемой и реагирующей согласно закону Франка — Старлинга исчезают при действии ганглиоблокаторов.
Местные сердечные рефлексы, осуществляемые метасимпатической нервной системой, регулируют уровень сердечной деятельности в соответствии с потребностями общей гемодинамики организма. Например, раздражение рецепторов растяжения при усилении притока крови и переполнении коронарных сосудов сопровождается ослаблением, силы сердечных сокращений, уменьшением притока крови. В результате в артериальную систему перекачивается меньший объем крови. Она задерживается в венах, обладающих большей емкостью, предотвращая, таким образом, внезапный выброс излишней крови в артерии, который мог бы привести к тяжелым последствиям для организма.
Опасно для организма и уменьшение сердечного выброса. Оно может вызвать критическое для жизни падение артериального давления. При недостаточном растяжении механорецепторов сердца из-за слабого заполнения его камер кровью возникает процесс рефлекторного возрастания силы сердечных сокращений, что в результате приводит к более интенсивному перекачиванию крови в аорту, усилению притока из вен и более полному заполнению камер во время диастолы.
Следовательно, осуществляемые метасимпатической нервной системой внутрисердечные периферические рефлексы выполняют в организме защитную роль, поддерживая стабильность наполнения кровью артериальной системы.
8.4.4. Экстракардиальная нервная регуляция
Экстракардиальный уровень регуляции обеспечивают специальные, супраспинальные и корковые механизмы, передающие свои влияния по волокнам блуждающих и симпатических нервов.
Эволюция внесердечных регуляторных нервных механизмов шла по пути специализации и совершенствования. У моллюсков сердце снабжено ускоряющими и тормозными нервными волокнами, идущими в составе висцеральных нервов. У одних видов моллюсков при стимуляции этих волокон преобладает торможение сердечной деятельности. При этом один залп импульсов может вызвать задержку одного сердечного сокращения. У других видов более выражено ускорение, у третьих — вслед за ускорением наступает торможение. Ускоряющие волокна имеют более высокий порог, чем тормозные. При повторном раздражении может возрастать амплитуда сердечных сокращений.
Строение экстракардиальных путей ракообразных более совершенно. Из ЦНС к сердцу направляется несколько нервов. Они регулируют пейсмекерный ганглий и сердечную мышцу. У отдельных видов к сердцу подходят две пары ускоряющих и одна пара тормозных нервов, берущих начало в подглоточном ганглии.
Внесердечные механизмы регуляции у высших позвоночных принципиально одинаковы. Деятельностью сердца у них непосредственно управляют сердечные центры продолговатого мозга и моста влияния которых передаются сердцу по блуждающим и симпатическим нервам (рис. 8.16).
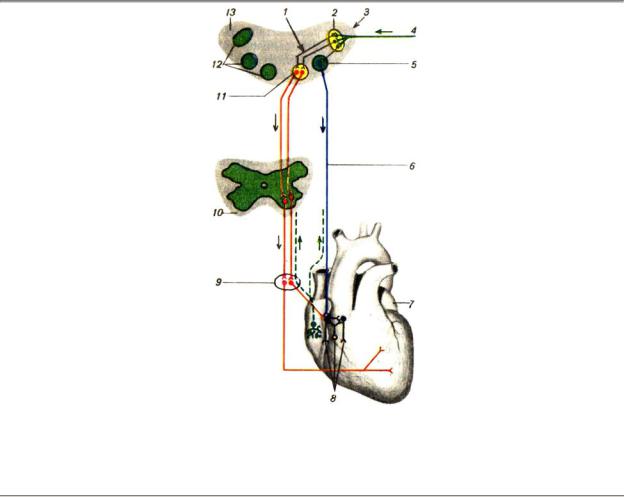
Рис. 8.16. Общая иннервация сердца: 1 — мозжечковые влияния, 2 — ядро солитарного тракта, 3 — гипоталамические влияния, 4 — афференты от периферических механо- и хеморецепторов, 5 — ядро блуждающего нерва, 6 — блуждающий нерв, 7 — сердце, 8 — метасимпатическая нервная сеть, 9 — симпатический узел, 10 — спинной мозг, 11 — ядро сосудодвигательного центра, 12 — стволовые центры, 13 — ствол мозга
Регистрация электрических процессов в одиночных чувствительных нервных волокнах предсердия выявила существование двух типов механорецепторов. На активное напряжение реагируют А-рецепторы, на пассивное растяжение отвечают В-рецепторы. Волокна от этих специализированных рецепторов идут в составе блуждающего нерва. Помимо того, имеется большая группа свободных нервных окончаний, локализующихся преимущественно в субэндокардиальном сплетении. Их волокна проходят в составе симпатических путей и передают сигналы болевой чувствительности.
Вагусные влияния. В продолговатом мозгу располагается заднее ядро блуждающего нерва. Аксоны клеток этого ядра в составе правого и левого нервных стволов направляются к сердцу и образуют синапсы на моторных метасимпатических нейронах интрамуральных ганглиев. Волокна правого блуждующего нерва распределяются преимущественно в правом предсердии. В соответствии с этим связанные с ним метасимлатические нейроны инвервируют миокард, коронарные сосуды и особенно густо пронизывают синусно-предсердный узел. Напротив, волокна левого блуждающего нерва через посредство метасимпатических нейронов передают свои влияния предсердножелудочковому узлу. В результате такой структурной топографии стимуляция правого блуждающего нерва сказывается преимущественно на частоте сердечных сокращений, левого — на предсердно-желудочковом проведении.
Впервые влияние блуждающих нервов на сердце обнаружили братья Э. и Г. Веберы (1845). Они установили, что стимуляция этих нервов уменьшает частоту сокращений сердца и ослабляет их силу. Как было показано позднее, сильные раздражения периферического конца перерезанного блуждающего нерва или введение ацетилхолина в сердце сопровождаются замедлением сердечного ритма, или так называемым отрицательным хронотропным эффектом. Одновременно возникает уменьшение амплитуды сокращений
сердца — отрицательный инотропный эффект, понижается возбудимость сердечной мышцы — отрицательный батмотропный эффект, уменьшается скорость проведения возбуждения в сердце — отрицательный дромотропный эффект. При этом иногда возникает полная блокада проведения возбуждения в предсердно-желу-дочковом узле.
Сильное раздражение блуждающих нервов может вызвать полную остановку сердечной деятельности, однако прекратившиеся вначале сокращения сердца, несмотря на продолжающееся раздражение, способны постепенно восстановиться. Это явление получило название ускользания сердца из-под влияния блуждающего нерва.
Во время умеренной стимуляции блуждающего нерва хронотропный эффект из-за замедления диастолической деполяризации может изменяться. При этом мембранный потенциал сердечных клеток приобретает пороговое значение позднее, чем в обычных условиях. Во время сильных раздражений диастолическая деполяризация исчезает совсем и возникает гиперполяризация клеток-водителей ритма. Отрицательный инотропный эффект связан с укорочением ПД, хотя форма последнего остается неизменной. Слабое раздражение блуждающих нервов может быть подобным действию симпатических нервов. Это явление объясняется тем, что в составе интрамуральных метасимпатических эффекторных нейронов кроме холинэргических находятся нейроны с адренэргическим или другим видом передачи, обладающие более низким порогом возбуждения.
Эффект блуждающего нерва при одной и той же силе раздражения может иногда сопровождаться и противоположными реакциями. Это связано со степенью наполнения сердца и сердечных сосудов кровью, т. е. с активностью собственного внутрисердечного нервного аппарата. При переполнении или значительном наполнении раздражение волокон блуждающего нерва сопровождается появлением тормозных реакций. Напротив, при слабом наполнении сердца и соответственно слабом возбуждении механорецепторов внутрисердечной нервной сети проявляются стимулирующие влияния.
Следовательно, наполнение кровью артериальной системы посредством насосной функции сердца регулируется двумя дублирующими механизмами его деятельности — собственными внутрисердечным и центральным, передаваемым по волокнам блуждающего нерва.
Симпатические влияния. Влияние на сердце раздражения симпатического нерва впервые исследовали И. Ф. Цион (1867), а затем в 80-х гг. одновременно И. П. Павлов и В. Гаскелл. Оказалось, что симпатический нерв, как и блуждающий, влияет на все стороны сердечной функции. Однако это влияние имеет противоположную направленность по сравнению с раздражением блуждающего нерва и проявляется в учащении сокращений сердца — положительный хронотропный эффект, усилении сокращений предсердий и желудочков — положительный инотропный эффект, улучшении проведения возбуждения в сердце — положительный дромотропный эффект повышении возбудимости сердца — положительный батмотропный эффект.
Среди симпатических ветвей, идущих к сердцу, как установил И. П. Павлов, находятся волокна, раздражение которых вызывает избирательное увеличение силы сердечных сокращений, это усиливающий нерв сердца. По мнению И. П. Павлова, этот нерв играет трофическую роль, он оказывает влияние на проведение возбуждения в сердечной мышце. Посредством его стимуляции можно снять блокаду проведения возбуждения в предсердно-желудочковом узле.
При положительном инотропном эффекте восходящая часть кривой, отражающей сокращение предсердий и желудочков, бывает круче, уменьшается интервал от начала сокращения до пика. При этом увеличивается скорость расслабления миокарда. Положительный дромотропный эффект относится только к предсердно-желудочковому узлу. Симпатическая стимуляция усиливает в нем проведение возбуждения и тем самым укорачивает интервал между сокращениями предсердий и желудочков. Что касается батмотропии, под которой понимают влияние на возбудимость ткани, то стимуляция симпатических нервов повышает возбудимость лишь в том случае, если она была перед

этим снижена. (Некоторые авторы считают понятие батмотропного эффекта неясным и путаным и предлагают вообще от него отказаться.)
Вызываемые раздражением симпатических волокон эффекты возникают через большие промежутки времени, превышающие 10 с, и продолжаются еще длительное время после окончания раздражения. Характерно, что при одновременном раздражении симпатических
иблуждающих нервов преобладает действие на сердце блуждающих нервов.
Воснове эффектов, вызываемых стимуляцией периферических концов тех и других нервов, лежит выделение биологически активных веществ, с помощью которых осуществляется передача возбуждения. Впервые это было установлено австрийским фармакологом О. Леви (1921). Он соединил два изолированных сердца лягушки с двумя концами одной и той же канюли (рис. 8.17). Сильное раздражение блуждающего нерва одного из сердец вызывало остановку не только иннервируемого этим нервом сердца, но и другого, интактного, связанного с первым только общим раствором канюли. Следовательно, при раздражении первого сердца в раствор выделялось вещество, влияющее на второе сердце. Это вещество было названо «вагусштофф» и оказалось впоследствии ацетилхолином. При аналогичном раздражении симпатического нерва сердца было получено другое вещество — «симпатикусштофф», представляющее собой адреналин или норадреналин, сходные по своему химическому строению.
Рис. 8.17. Опыт О. Леви. А — схема опыта; Б — кривая сердечных сокращений при раздражении блуждающего нерва: 1 — денервированное сердце лягушки, 2 — сердце лягушки с оставшейся веточкой блуждающего нерва, 3 — стимулятор, 4 — вагусная остановка раздражаемого сердца, 5 — остановка второго сердца
Действие ацетилхолина на сердце в первую очередь основано на повышении мембранной проницаемости для иона К+, препятствующей развитию деполяризации. С этим же явлением связано запаздывание в синусно-предсердном узле медленной диастолической деполяризации, что сопровождается укорочением ПД кардиомиоцитов и в итоге ослаблением сокращения сердца. В предсердно-желудочковом узле усиленный отток К+ противодействует входящему току ионов Са2+. Ацетилхолин быстро разрушается и оказывает только местное действие, связываясь с определенными молекулярными структурами клеток — холинорецепторами. Антагонистом ацетилхолина и раздражающих влияний блуждающего нерва в сердце является атропин.
По-иному действует на возбудимые структуры сердца норадреналин. Его действие связано с ростом мембранной проницаемости для Са2+, что сопровождается повышением степени сопряжения возбуждения и сокращения миокарда, а также потенциалозависимой проницаемости для К+. Норадреналин может в определенных пределах восстанавливать изменение функции сердца при сдвигах содержания калия в крови во время чрезмерных физических нагрузок или других состояний. Сниженная избытком К+ возбудимость и проводимость предсердий и желудочков восстанавливается норадре-налином. Норадреналин разрушается значительно медленнее, чем ацетилхолин, поэтому его взаимодействие с адренорецепторами сердечных клеток, как и раздражение симпатических волокон, сопровождается более продолжительным эффектом.
8.4.5. Гуморальная регуляция
Гуморальная регуляция деятельности сердца осуществляется биологически активными веществами, выделяющимися в кровь и лимфу из эндокринных желез, а также ионным составом межклеточной жидкости. Эта регуляция в наибольшей степени присуща адреналину, секретируемому мозговым слоем надпочечников. Адреналин выделяется в кровь при эмоциональных нагрузках, физическом напряжении и других состояниях (см. разд. 5.7.2; 6.4.1). Его взаимодействие с бета-адренорецепторами кардиомиоцитов приводит к активации внутриклеточного фермента аденилатциклазы. Последний ускоряет образование циклического АМФ (цАМФ). В свою очередь, цАМФ необходим для превращения неактивной фосфори-лазы в активную. Активная фосфорилаза обеспечивает снабжение миокарда энергией путем расщепления внутриклеточного гликогена с образованием глюкозы. Адреналин повышает также проницаемость клеточных мембран для ионов Са2+.
Важное значение имеет гормон поджелудочной железы — глюкагон. Он оказывает на сердце положительный инотропный эффект путем стимуляции аденилатциклазы. Гормон щитовидной железы — тироксин — увеличивает частоту сердечных сокращений и повышает чувствительность сердца к симпатическим воздействиям. Гормоны коры надпочечников — кортикостероиды, биологически активный полипептид — ангиотензин, гормон энтерохромаффинных клеток кишки — серотонин — увеличивают силу сокращений миокарда.
Большое влияние на деятельность сердечной мышцы оказывает лонный состав среды. Повышение содержания во внеклеточной среде К+ угнетает деятельность сердца. При этом вследствие изменения градиента концентрации иона увеличивается проницаемость мембран для К+, падают возбудимость, скорость проведения возбуждения и длительность ПД. В этих условиях синусно-предсердный узел перестает выполнять роль водителя ритма. Подобным образом на сердце влияют ионы НСО3- и Н+ Ионы Са2+ повышают возбудимость и проводимость мышечных волокон, активируя фосфорилазу и обеспечивая сопряжение возбуждения и сокращения.
8.4.6. Тонус сердечных нервов
Как уже было показано, первая ступень иерархии нервных центров, регулирующих нормальную работу сердца, представлена местными рефлексами, которые осуществляются метасимпатической нервной системой. Второй ступенью являются ядра блуждающих и симпатических центров, обладающих характерным тоническим свойством, Под тонусом обычно понимают длительное, то усиливающееся, то ослабевающее возбуждение, не сменяющееся торможением (см. разд. 5.4.4).
У человека и большинства млекопитающих работа предсердий и синусно-предсердного узла постоянно контролируется блуждающим и симпатическим нервами, в то время как желудочки находятся под контролем преимущественно симпатических нервов. В существовании тонуса блуждающих нервов легко убедиться, перерезав у животного оба блуждающих нерва. В этом случае частота сердечных сокращений возрастает почти вдвое. Отсюда может быть сделан вывод, что автоматическая деятельность сердца все время подвергается угнетению импульсами, идущими по волокнам блуждающего нерва. Наличие этих разрядов легко прослеживается при регистрации эфферентной импульсации. В покое тонус блуждающих нервов преобладает над тонусом симпатическим.
Тонус блуждающих нервов (см. разд. 5.4.4) возникает в результате притока импульсации от рецептивных зон дуги аорты, каротидного синуса, кроме того, он обусловлен восходящими активирующими влияниями ретикулярной формации, клеткам которой свойственна постоянная импульсная активность. Она связана с конвергенцией
коллатералей огромного числа чувствительных путей, а также с высокой чувствительностью ее клеточных мембран.
В поддержании тонуса участвуют различные гуморальные раздражители, такие, как гормоны, уровень парциального давления СО2. Тонус блуждающих нервов находится в зависимости и от фаз дыхательного цикла. Во время выдоха он повышается, что влечет за собой урежение частоты сердечных сокращений. Это обычное в нормальных условиях состояние называют дыхательной аритмией. Дыхательная аритмия исчезает после атропинизации животных или перерезки у них блуждающих нервов.
Определенным тонусом обладают также симпатические клетки, посылающие свои аксоны к сердцу. После леререзки всех симпатических путей или удаления основных источников симпатической иннервации сердца — звездчатых ганглиев — ритм сердца собаки снижается на 15—25 %. При полной симпатической и парасимпатической денервации сердца оно начинает сокращаться в ритме, который задается синуснопредсердным узлом. Этот собственный ритм сердца несколько выше, чем ритм интактного сердца.
Таким образом, перерезка блуждающих и симпатических нервов, равно как и их раздельная стимуляция, приводит к противоположным изменениям различных показателей сердечной деятельности. Однако это не означает, что действие этих нервов на сердце взаимно нейтрализуется. Напротив, нормальная работа сердца определяется взаимодействием влияний, поступающих к нему по блуждающим и симпатическим путям.
8.4.7. Гипоталамическая регуляция
Ядра гипоталамуса являются следующей ступенью в иерархии нервных центров, регулирующих сердечную деятельность (см. разд. 6.2). В процессе филогенеза эта функция возникает с первым появлением гипоталамуса среди позвоночных — у рыб. Гипоталамус рыб может облегчать и тормозить сердечные рефлексы. У Млекопитающих посредством прямых связей гипоталамус соединяется с ядрами блуждающих нервов в продолговатом мозгу и боковыми рогами спинного мозга, т. е. с центрами парасимпатической и симпатической иннервации сердца.
У наркотизированных животных при раздражении различных зон гипоталамуса удалось выявить точки, стимуляция которых сопровождается изменением либо только ритма сердца, либо только силы сокращений или расслабления миокарда левого желудочка. Следовательно, в гипоталамусе существуют отдельные структуры, регулирующие определенные функции сердца. При раздражении одной и той же точки гипоталамуса возникающие эффекты чаще всего обусловлены одновременно и симпатическими, и парасимпатическими механизмами. По своей силе и выраженности эти реакции значительно превосходят реакции в естественных условиях, где они не возникают изолированно друг от друга. Непосредственное участие в регуляции работы сердца принимают паравентрикулярное, вентромедиальное и преоптическое ядра, мамиллярные тела.
Гипоталамус — интегративный центр, который может изменять параметры сердечной деятельности для обеспечения текущих потребностей организма и всех его систем при различных поведенческих реакциях. Однако осуществляемая им перестройка деятельности висцеральных органов не является самостоятельной, а зависит от сигналов, поступающих из лимбической системы и коры головного мозга.
8.4.8. Корковая регуляция
Кора головного мозга является высшей ступенью в иерархии механизмов целенаправленного управления деятельностью сердца (см. разд. 5.5.7). Электрическая стимуляция почти любого участка коры мозга вызывает ответы сердечно-сосудистой
системы. Они проявляются особенно отчетливо при раздражении моторной и премоторной зон коры, поясной извилины, орбитальной поверхности лобных долей, передней части височной доли. Эти активные области совпадают с зонами проекций в кору больших полушарий блуждающего нерва и его сердечных ветвей. Анатомическое соседство пунктов, ответственных за возникновение двигательных реакций и реакций сердца, имеет важное значение для оптимального обеспечения сердцем поведенческих реакций индивидуума.
При относительно нормальном функционировании организма кора мозга, как и гипоталамус, способна тормозить или активировать работу сердца. Использование в экспериментах хирургической декортикации или распространяющейся калиевой депрессии изменяет характер исходных сердечных рефлексов, снимая модулирующее влияние коры полушарий.
Кора головного мозга является органом психической деятельности, обеспечивающим целостные приспособительные реакции организма. Многочисленные наблюдения показывают, что работа сердца изменяется под влиянием условий, влияющих на деятельность коры мозга: боли, страха, радости, тревоги, ярости и т. п. Например, у спортсменов наблюдается так называемое предстартовое состояние, проявляющееся учащением сердцебиения. Оно связано с активацией корой мозга гипоталамогипофизарной системы и надпочечников, выделяющих адреналин в кровь. Сигналы, непосредственно предвещающие возникновение этих ситуаций или возможность их наступления, способны по механизму условного рефлекса вызвать перестройку функций сердца, чтобы обеспечить предстоящую деятельность организма.
Влияние коры полушарий на работу сердца подтверждается многочисленными наблюдениями с образованием экстероцептивных и интероцептивных условных рефлексов. В обоих случаях модулирующие влияния коры направлены к центрам продолговатого и спинного мозга, где формируется эфферентная импульсация к сердцу. Она приспосабливает работу сердца к текущим или предстоящим потребностям организма в целом, отдельных органов и систем и, наконец, самого сердца. При очень сложных ситуациях, действиях чрезвычайных раздражителей могут возникнуть нарушения и срывы высших регуляторных механизмов (неврозы). В этих случаях параллельно с расстройствами поведенческих реакций возможны существенные нарушения деятельности сердца.
8.4.9. Рефлекторная регуляция
Емкость сосудистого русла варьирует в широких пределах в зависимости от условий существования организма в окружающей среде, а также при изменении деятельности любых его органов. В связи с этим частота и сила сердечных сокращений могут рефлекторно увеличиваться или уменьшаться. Изменение деятельности сердца обычно происходит параллельно с изменением состояния сосудов. Регуляция сердечной деятельности является сложным рефлекторным актом при участии центральных и периферических механизмов. По эфферентным нервным волокнам к сердцу передаются импульсы, возникающие рефлекторно в ответ на раздражение различных интеро- и экстероцепторов.
В системе кровообращения существует ряд рефлексов, имеющих важное гемодинамическое значение. Характерным признаком этих рефлексов является постоянство в расположении их рецептивных полей. К числу таких полей относятся (см. рис. 8.16) устья полых вен, дуга аорты и каротидный синус (область разветвления сонной артерии на наружную и внутреннюю). В этих рецептивных полях, именуемых иначе рецептивными зонами, сосредоточено огромное количество механо- и хеморецепторов, тонко реагирующих на различные изменения гемодинамики. Возникающие в этих рецепторах импульсы по чувствительным волокнам поступают к циркуляторным центрам