

3 курс / Общая хирургия и оперативная хирургия / Хирургическая_анатомия_венечных_артерий,_Бокерия_Л_А_,_Беришвили
.pdfБульбовентрикуляр- |
Атрезия правого атрио- |
Единственный желудочек |
Норма |
ная петля |
вентрикулярного отверстия |
|
|
Рис. 13. Изменения в строении венечно-артериальной системы на различных этапах формиро вания желудочков (на примере сердец с различным строением желудочков).
А - срез сердца (тотчас ниже атриовентрикулярных клапанов) на этапе образования бульбовентрикулярной петли; Б - срез сердца с атрезией правого атриовентрикулярного отверстия; В - срез сердца с единственным желудочком левожелудочкового типа; Г - срез нормального зрелого сердца. Вокругбульбарные (А), бульбовентрикулярные (В) и во кругжелудочковые (С, D, Е) венечно-артериальные кольца соединяются в точках Вх и By на линии стыка примитив ных бульбуса и желудочка. Линия Вх-Ву - ретробульбарный сегмент сплетения. Во всех сердцах сохраняется соеди нение остатков этих колец в точке Вх (в точке 1). Во всех остальных сердцах ретробульбарный сегмент венечно-арте- риального сегмента разорван (адсорбирован) задним артериальным клапаном (на рисунке - аортой). Его остатки пред ставлены левой ВА (простирающейся между 2-м лицевым синусом аорты и точкой 1) и правой ВА (простирающейся между 1-м лицевым синусом и точкой 2).
Начиная с сердец с атрезией правого атриовентрикулярного отверстия (Б) бульбовентрикулярное кольцо уже не соеди няется с остальными кругами в точке 2. Оно соединяется (точка 3) только с вокругпредсердным кольцом. По мере фор мирования синусной части (точечная штриховка) рвутся вокругжелудочковые круги и удаляются друг от друга точки 2 и 3. Разрыв круга Вьессена во многом определяется размерами выпускной камеры (В). Г - окончательное состояние системы венечных артерий в дефинитивном сердце. Разорванные вокругжелудочковые круги образуют дефинитивные ВА. При этом ОВ, ДВ и ВТК, являющиеся дериватами кругов С, D и Е, отходят от точки 1.
Всвязи с серьезными структурными изменениями в строении правого желудочка ВА, питающие его стенки, отходят от разных точек: конусная артерия (КА) - от точки 2, ветвь острого края (ВОК) - от точки 3 и ЗМЖВ - от точки 4 в об ласти задней межжелудочковой борозды. Из приведенной схемы понятно, что ЗМЖВ обнаруживается в нормальных сердцах и в некоторых сердцах с единственным желудочком, но не выявляется при атрезии правого атриовентрику лярного отверстия. В этих сердцах артерия, располагающаяся на границе желудочка и выпускника справа-сзади, представляет собой ветвь острого края (отходящую от точки 3).
Всвязи с развитием инфундибулярной камеры круг Вьессена в нормальном сердце, как правило, разорван.
вентрикулярного канала в сторону ПЖ, а благодаря нескольким факторам: экстенсив ному росту ограниченного участка приточного компонента, переднему перемещению задней стенки бульбуса и его росту, а также раскрытию бульбовентрикулярного отвер стия вследствие перечисленных выше изменений (см. рис. 14). Приведенная состав ная форма роста ПЖ также отличается от общепринятой (только ростом бульбуса), приведенной S. Rao (1982). Если зона экстенсивного роста располагается полностью в составе примитивного желудочка, процесс развития желудочкового миокарда осуще ствляется без интеграции тканей приточного компонента в состав ПЖ, что в итоге обусловливает формирование сердец с единственным желудочком.
Таким образом, процесс формирования ПЖ влечет за собой разрыв вокругжелудочковых кругов, осуществляя тем самым формирование задней межжелудочковой ветви (ЗМЖВ) правой ВА (из правого компонента круга Е) и диагональной ветви левой ВА (из левого компонента круга Е), а также заднебоковой ветви правой ВА (из право го компонента круга D) и ветви тупого края левой В А (из левого компонента круга D). В результате перемещения правого края вокругбульбарного круга из него формирует ся ветвь острого края правой ВА (см. рис. 9 и 13). Левый край этого круга формирует переднюю межжелудочковую ветвь (ПМЖВ) левой ВА, теряющей связь с ветвью ост рого края только по мере роста трабекулярной части и всего ПЖ .
Как следует из приведенных материалов, процесс роста желудочков и связанный с ним разрыв вокругжелудочковых кругов ответственны не только за формирование де финитивных ВА, определяемых в зрелом сердце, но и участвуют в формировании из вестных (правого, левого и сбалансированного) типов (доминантности) кровоснабже ния миокарда. Этот же процесс определяет и то, ветвью какой (правой или левой) ВА
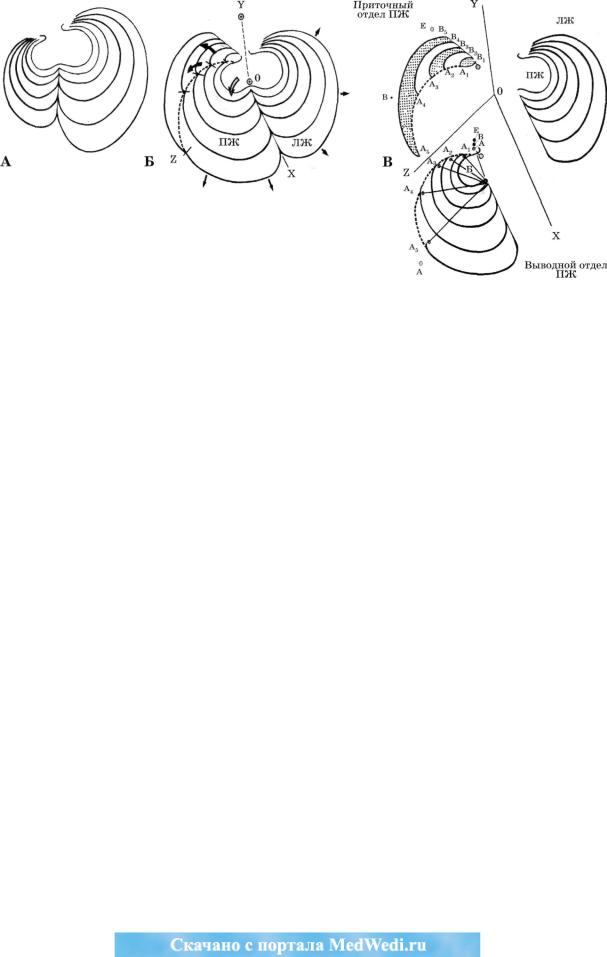
Рис. 14. Схема перемещения точек соединения венечно-артериальных колец с ростом правого желудочка.
А - известная схема роста желудочков из книги S. Rao «Tricuspid atresia» (N. Y., 1982, P. 32). Схема предусматрива ет концентрический рост желудочковых полостей в течение внутриутробной жизни. Как показали сопоставления сер дец с различной степенью дифференциации желудочкового комплекса (см. рис. 13), формирование правого желудоч ка отличается от такового, приведенного в книге. Для его формирования необходимо развитие синусной части и инте грация последнего в состав правого желудочка. За счет интенсивного роста этой части правого желудочка формирует ся приточный отдел и смещается вперед выводной. Этот же процесс предполагает пространственную переориентацию
ираскрытие бульбовентрикулярного окна. Будучи первоначально отверстием между бульбусом и примитивным желу дочком, в дефинитивном сердце оно разграничивает приточный и выводной отделы. Б - общая схема формирования и роста желудочкового комплекса, разработанная нами на основании приведенных данных. В соответствии с приведен ными материалами нами проанализированы изменения в венечных артериях. В - раздельная схема роста приточного
ивыводного отделов правого и левого желудочков.
Первоначально вокругбульбарные (А), бульбовентрикулярное (В) и вокругжелудочковое (Е) кольца сливаются (см. в бульбусе (Б), из которого формируется выводной отдел нормального сердца).
Перемещения точки отхождения вокругбульбарного кольца (А) с ростом ПЖ определяют точки Ах -А5 . А - точка отхождения конусной артерии от правой ВА. Перемещения точки отхождения бульбовентрикулярного кольца (В) с рос том приточного отдела правого желудочка определяют точки Bj—В5. В — точка, отхождения ветви острого края. Е — точ ка отхождения ЗМЖВ.
Формирование дефинитивного ЛЖ из примитивного желудочка осуществляется за счет концентрического роста послед него (при утере правожелудочковой части синусного отдела и приобретении аортальной части артериального конуса).
На схеме Б: 0Y - проекция задней межжелудочковой перегородки, ОХ - проекция передней межжелудочковой перегород ки, 0Z - проекция наджелудочкового гребня. Светлой изогнутой стрелкой показано переднее смещение выводного отдела правого желудочка, темными изогнутыми стрелками показаны направления развития приточного отдела правого желу дочка, пунктирной изогнутой кривой - граница между приточным и выводным отделами ПЖ на разных этапах его роста, малые черные стрелки - направление роста выводного отдела правого и левого желудочков. Линии 0 - Ад -А5 показывают раскрытие и изменение ориентации бульбовентрикулярного окна.
будет зад небоковая левожелуд очковая артерия в будущем зрелом сердце. Абсолютно идентично и формирование типов кровоснабжения предсердия, обусловленное разры вом задней части вокругпредсердного круга (С) (см. рис. 9,Б).
Если разрыв вокругжелудочкового круга (Е) происходит правее креста сердца (или задней межжелудочковой борозды), формируется левый тип доминантности, ле вее - правый. Если целостность этого круга сохранена за счет вокругпредсердного кру га, формируется сбалансированный тип кровоснабжения миокарда. Как свидетельст вует анализ собственного материала, несогласующегося, кстати, с большинством дан ных отечественных исследователей (Р. Н. Яшвили, 1962; А. Т. Юрченко, 1969; С. В. Са мойлова, 1970; Л.А.Арсентьева, 1972; В. В. Кованое и Т. Н.Аникина, 1974; И. X. Рабкин, 1977), в подавляющем числе наблюдений формируется правый тип доминантнос ти. Обусловлено это, на наш взгляд, двумя факторами: во-первых, разрыв венечного сплетения правее креста сердца требует разрыва трех кругов (С, Д, Е), левее - одного (D); во-вторых, рост левого желудочка больше удаляет правые края вокругбульбарных и вокругжелудочковых кругов от его левого края, чем от правого (см. рис. 9).
Аналогично при разрыве первого вокругжелудочкового круга (Е) сразу левее крес та сердца заднебоковая левожелудочковая ветвь перфузируется левой ВА, а если на удалении, левее от него, - правой ВА. Вероятность возникновения того или иного ва рианта кровоснабжения (правого или левого) в этих случаях одинакова (см. рис. 9).
Таковы основные изменения в вокругжелудочковых кругах и перемещения в бульбовентрикулярном круге, связанные с формированием ПЖ . Но процесс интег рации приточного отдела в состав ПЖ обусловливает и ротацию конотрункуса против часовой стрелки. На наш взгляд, этот фактор определяет хронологию присоединения ВА (правой и левой) к аорте.
Согласно данным литературы, сроки появления В А не вызывают разногласий. Сначала формируется левая ВА, затем - правая (Л. Г. Шелия, 1985; Н. Bennett, 1936; KHirakow, 1983;A.Bogers, 1989).
Как следует из материалов, приведенных в скрупулезной работе A. Bogers (1989), положение аорты и легочной артерии в сердце плода к моменту присоединения ВА и в нормальном зрелом сердце практически одинаково, что, по мнению автора, ис ключает факт ротации конотрункуса или абсорбции конуса в процессе формирования сердца. Между тем однотипность ориентации оси конотрункуса в сердце плода и ново рожденного, как и сохранение количественных пропорций между выводными отдела ми ПЖ и ЛЖ, обнаруженных автором, на наш взгляд, вовсе не означает отсутствия ротации конуса в формирующемся сердце, и уж во всяком случае не исключает ее. Бо лее того, мы полагаем, что в процессе интеграции приточного компонента бульбарный отдел обязательно перемещается вперед, ротируя при этом конотрункус против часо вой стрелки. Однотипность же ориентации оси конотрункуса в сердце плода и ново рожденного объясняется столь же обязательной обратной ротацией конотрункуса (по часовой стрелке) в процессе рассасывания бульбовентрикулярной складки и вовлече ния аорты в состав ЛЖ . Кстати, перед нелевая ротация бульбуса обусловливает и со прикосновение (и сращение) до этого не сливающихся краев конусной перегородки и бульбовентрикулярной складки, а обратная ротация - удлинение и завитие легочного ствола вокруг аорты, что определяется в дефинитивном сердце.
Более того, на наш взгляд, именно ротация конотрункуса позволяет объяснить по следовательность соединения аорты с левой и правой ВА, что трудно понять из материа лов предыдущих исследователей. К тому же мы полагаем, что именно ротация позволя ет объяснить формирование вариантов с единственной ВА, кстати говоря, чаще - левой.
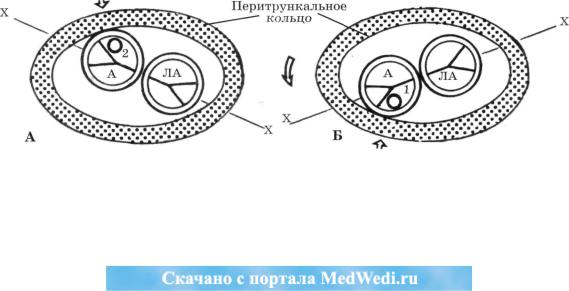
Итак, мы уже отмечали, что в результате изменения конфигурации неразделенно го артериального отверстия создаются условия для сближения с ним перитрункального кольца. Первым стенку аорты пенетрирует изогнутый задний сегмент (будущий ле вый ствол венечно-артериальной системы) вокругартериального кольца, а по мере ро тации конотрункуса против часовой стрелки - передний (будущий правый ствол ве нечно-артериальной системы) (рис. 15).
Рис. 15. Присоединение венечно-артериального сплетения ко 2-му лицевому синусу аорты по мере переднего перемещения бульварной части сердца (А) и к 1-му лицевому синусу аорты по мере ротации конуса против часовой стрелки (Б).
Изменение оси артериальных клапанов (Х-Х) на разных этапах формирования сердца. А - аорта, ЛА - легочная арте рия, 1 и 2 - 1-й и 2-й лицевые синусы аорты.

После присоединения заднего сегмента венечно-артериального сплетения к аорте обо собленная до этого единая венечно-артериальная система преобразуется в часть систем ной циркуляции и до соединения ее переднего сегмента с аортой представляет собой пред вестник однокоронарной (левой) венечной системы. Как свидетельствует анализ наших данных, для двойственности присоединения перитрункального кольца к аорте необхо дим разворот оси конотрункуса чуть кпереди от линии, параллельной прямой, проведен ной вдоль заднего края атриовентрикулярных клапанов. Но и на этом этапе системы пра вой и левой В А не разобщены (сохраняется целостность перитрункального кольца).
Процесс разобщения общего венечно-артериального сплетения на систему обособ ленных правой и левой ВА осуществляется позже и определяется развитием инфундибулума и обратной ротацией конотрункуса по часовой стрелке.
Первый разрыв перитрункального кольца обусловлен ротацией конотрункуса по ча совой стрелке. При этом в результате заднего смещения заднего артериального клапана в процессе рассасывания бульбовентрикулярной складки, образования митрально-полу лунного фиброзного контакта и вовлечения этого клапана в состав ЛЖ разрывается сли вающаяся часть (ретробульбарный отдел перитрункального кольца) сплетения. При ро тации конуса по часовой стрелке таким клапаном является аортальный, и в левый же лудочек инкорпорируется аортальный клапан. В этих случаях образуется спектр сердец от ОАЛА ПЖ до нормы, и В А располагаются сзади только легочного клапана (рис. 16). При ротации конуса против часовой стрелки общий венечно-артериальный круг разры вается легочным клапаном, и последний включается в состав ЛЖ (см. рис. 16). В таких случаях образуется спектр сердец от ОАЛА ПЖ до анатомически корригированной мальпозиции аорты и легочной артерии (АКМАЛА). В этих случаях ВА располагаются сзади только от аорты. Персистенция этой части перитрункального кольца возможна только при сохранении мышечной прослойки между одним из полулунных клапанов и митральным клапаном, что имеет место в сердцах с отхождением аорты и легочной ар терии от ПЖ (ОАЛА ПЖ) или ОАЛА ЛЖ (рис. 16, 17). Отсюда высокая вероятность су ществования единственной (левой) ВА в сердцах с ОАЛА ПЖ или ЛЖ .
Перитрункальное и вокругбульбарные кольца полностью и сливающиеся с ними фрагменты вокругпредсердного и бульбовентрикулярных колец образуют так называ емый круг (круги) Вьессена.
То, что обычно принято считать конусной артерией (ветвью), представляет собой ос татки фетального круга Вьессена (рис. 18, 19) с разорванными вследствие роста подле гочного инфундибулума (см. рис. 9,А и 10) вокругбульбарными кольцами (при рота ции конуса по часовой стрелке) и подаортальным кольцом (при ротации конуса против часовой стрелки). Разрыв перитрункального и вокругбульбарных колец завершает формирование двух обособленных систем (правой и левой) венечного кровообращения.
Таким образом, мы разделяем мнение S. Van Praagh и соавт. (1982) о том, что круг Вьессена представляет собой часть первичного фетального венечно-артериального сплетения. Мы отмечали уже, что по мере разворота конотрункуса круг Вьессена рвет ся за одним из артериальных клапанов. По мере же развития инфундибулума перед него артериального сосуда круг Вьессена рвется и спереди артериальных клапанов и перестает существовать как круг1 1 (см. рис. 9). Поэтому в дефинитивном сердце пе редняя часть круга Вьессена чаще обнаруживается в виде небольшой конусной ветви правой ВА (ПВА) или самостоятельной конусной артерии (КА). Иногда можно просле дить несколько веточек КА, отходящих от ПВА (остатки трех вокругбульбарных кру гов; см. рис. 9). При недоразвитии же подлегочного инфундибулума (например при тетраде Фалло) любая из веточек (коммуникаций между ПВА и ПМЖВ, представляю щих собой фетальные вокругбульбарные кольца круга Вьессена) может сохраниться. Согласно данным литературы, К А может отходить от ПВА (см. рис. 18, Б), от ЛВА
11 С этого же момента прекращает свое существование перитрункальное кольцо, и образуются две обособ ленные системы ВА - правая и левая.
40
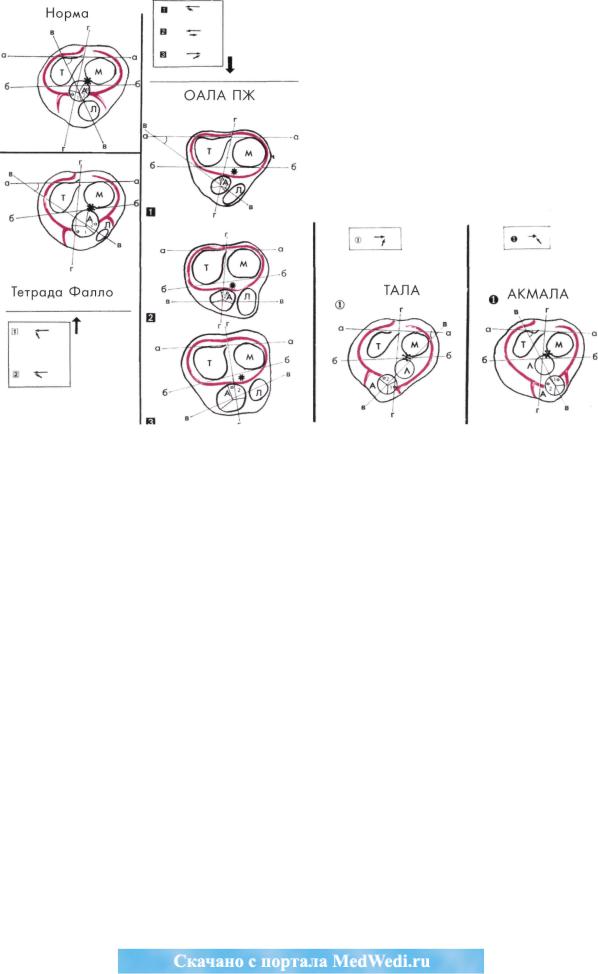
Рис. 16. Изменения в строении венечно-артериальной системы, обусловленные стороной рота ции конуса.
В исходной позиции бульбовентрикулярная складка сохранена (показано звездочкой), а артериальные клапаны отделе ны от атриовентрикулярных. При этом ось артериальных клапанов может быть сближена с трехстворчатым (Т) клапа ном Q, митральным (М) клапаном 0 и параллельна линии, проведенной по заднему краю атриовентрикулярных кла панов 0 • Независимо от небольших ротационных изменений в положении конуса, характерного для сердец с ОАЛА ПЖ, в большинстве сердец с этим пороком сохраняется ретробульбарный сегмент перитрункального кольца. Последнее соединяется со 2-м лицевым синусом аорты, образуя единственную (левую) венечную артерию.
При ротации конуса по часовой стрелке образуется спектр сердец от тетрады Фалло до нормы. В этих сердцах ретро бульбарный сегмент перитрункального кольца разрывается аортой. Разорванные фрагменты пенетрируют лицевые си нусы аорты.
При ротации конуса против часовой стрелки образуется спектр сердец от ТАЛА до АКМАЛА. Ретробульбарный сегмент перитрункального кольца в этой группе разрывается легочной артерией. Разорванные фрагменты пенетрируют лицевые синусы аорты. В обеих последних группах адсорбирована бульбовентрикулярная складка, сформирован митрально-полу лунный фиброзный контакт (показан звездочкой) и формируется двухкоронарная система кровообращения сердца.
А - аорта, Л - легочная артерия, а-а - линия, проведенная по заднему краю атриовентрикулярных клапанов, б-б - ли ния, проведенная по переднему краю атриовентрикулярных клапанов, в-в - линия, проведенная по серединам полу лунных клапанов, г-г - линия, проведенная по проекции задней межжелудочковой перегородки. ОАЛА ПЖ - отхождение аорты и легочной артерии от правого желудочка, ТАЛА - транспозиция аорты и легочной артерии, АКМАЛА - анатомически корригированная мальпозиция аорты и легочной артерии. Q, 0, 0 — варианты ОАЛА ПЖ .
(см. рис. 19) и самостоятельно как от первого, так и от второго лицевых синусов аор ты (см. рис. 18,Б) (Н. В. Архангельская, 1959; С. Meng et al., 1965; W. McAlpine, 1975; B.Edwards et al., 1981; A. Gittenberger-de Groot et al., 1983). Самостоятельное отхождение КА от 1-го лицевого синуса аорты мы наблюдали в 20 случаях, от 2-го - в одном.
Как уже отмечалось, в первичной фетальной венечно-артериальной системе вокругбульбарные и предсердно-желудочковые кольца сливаются. При соединении этих колец с аортой проксимальнее места слияния в зрелом сердце обнаруживается отхождение ВА одним устьем от одного лицевого синуса (см. рис. 18,Б). Если же вокругбульбарные кольца внедряются в аорту до места их слияния, в зрелом сердце обнаруживается само стоятельное отхождение КА одним или несколькими устьями (см. рис. 18, В).
Таким образом, в норме ретробульбарный сегмент перитрункального кольца от сутствует. Тут следует, однако, учитывать, что этот сегмент первоначально пред-

Рис. 17. Механизм формирования единственной венечной артерии (ЕВА) при отхождении аор ты и легочной артерии от правого желудочка (ОАЛА ПЖ).
При данном пороке бульбовентрикулярная складка сохранена (А - показана стрелками), аортальный (А) и митраль ный (М) клапаны разобщены. Поэтому ретробульбарный сегмент перитрункального кольца сохранен (Б). При отсутст вии ротации конуса передний фрагмент перитрункального кольца не сближается с 1-м лицевым синусом аорты (1). Ве- нечно-артериальное сплетение соединено только со 2-м лицевым синусом аорты (2). Поэтому при ОАЛА ПЖ чаще все го имеется единственная левая ВА.
ЛА - легочная артерия, Т - трехстворчатый клапан.
Рис. 18. Дефинитивные венечные артерии бульбуса.
Конусная артерия (КА) представляет собой дериват одного из кругов Вьессена, сформированный как результат его разрыва вследствие роста подлегочного инфундибулума
(А). При слиянии вокругбульбарных и предсердно-желу- дочковых колец до их соединения с аортой, КА отходит от правой венечной артерии (Б); если эти кольца соединяют ся с аортой без предварительного слияния друг с другом, КА отходит от аорты самостоятельно (В).
Обозначения те же, что и на рис. 10.
ставляет собой многоканальное соеди нение. Даже полное рассасывание этого сегмента в сердцах с полулунно-мит ральным контактом не означает одно временного исчезновения фрагмента вокругпредсердного круга: по мере расса сывания бульбовентрикулярной склад ки этот ствол сплетения смещается кверху, к стенке предсердия, служа ос новой формирования предсердных вет вей (ветвей синусного узла) от правой или левой ВА. Более того, в исключи тельных случаях возможно и сохране ние перитрункального кольца. В таких случаях имеет место отхождение огиба ющей ветви (ОВ) от первого лицевого си нуса аорты или от правой ВА.
Завершая анализ формирования двухвенечного кровообращения, пред ставляется логичным отметить и разли чия в соединении ВА при пороках коно трункуса. Как известно, при всем много образии вариантов строения ВА, крово снабжение миокарда не страдает ни при
одном |
ВПС |
(J.Meyer |
et al., 1975; |
R.Hurwitz |
et |
al., 1980; |
D.Velimirovich |
et al., |
1989). |
Наши |
исследования |
(И. И. Беришвили и др., 1994) полностью подтверждают эти предположения. Как показал анализ нашего материала, в
норме во всех сердцах независимо от различий в ротационных особенностях конотрункуса сохранены проксимальные сегменты всех (кроме перитрункального кольца) межсегментных колец венечно-
42

Рис. 19. М е х а н и з м ы ф о р м и р о в а н и я р а з л и ч н ы х вариантов строения конусной артерии .
Изолированная крупная конусная артерия может быть образована вследствие сохранения правого фрагмента низшего из кругов Вьессена (1), фрагментов одного из этих кругов (2-4) или сохранения длинного левого фрагмента круга Вьессена (5). В зависимости от числа сохранившихся кругов Вьессена (1,2) отмечаются различия в кровоснабжении вывод ного отдела правого желудочка. «Ярусность» конусной артерии - следствие сохранения правого (1) или левого (2) фрагментов первичных кругов Вьессена.
артериального сплетения1 2 . Как уже отмечалось, по мере перестановок во взаимоот ношениях между кольцами и их соединения в ретробульбарной области создаются предпосылки для приобретения этими кольцами коммуникационных функций. В норме они соединены с перитрункальным кольцом. Перитрункальное кольцо в зави симости от ротационных особенностей конотрункуса и рассасывания бульбовентрикулярной складки сохраняет одни и редуцирует другие сегменты. Сохранение тех или иных фрагментов вокругартериального кольца зависит от переднезадних пере мещений артериальных сосудов вследствие рассасывания бульбовентрикулярной складки (и соответственно ротации конотрункуса) и развития инфундибулума пе реднего артериального сосуда. Различия в строении ВА при пороках конотрункуса заключаются только в подсоединении к различным фрагментам перитрункального кольца, сохраняющимся при различных векторах разворота конотрункуса.
В группе сердец с ротацией конотрункуса по часовой стрелке редуцируются фраг менты за аортальным клапаном и впереди легочного клапана. В сердцах с ротацией ко-
12 В литературе имеется несколько сообщений (одно на основании данных ангиографического исследова ния) об отсутствии ОВ левой ВА (Г. И. Цукерман и др., 1976; Е. Mievis et al., 1979; R. Bestetti et al., 1985) или ПМЖВ (D. Ferguson et al., 1985).

нотрункуса против часовой стрелки, наоборот, облитерируются фрагменты за легоч ным клапаном и впереди аортального клапана. Кроме того, редуцируются и дублирую щие фрагменты перитрункального кольца (что исключает возможность бесполезного циркулирования крови по перитрункальному кольцу) и сохраняются фрагменты, обес печивающие кратчайшее соединение с ветвями-коммуникантами (ОВ ПВА, КА, ОВ ЛВА, ПМЖВ). При этом меняются источник кровоснабжения этих ветвей (лицевой си нус и ствол ВА, кровоснабжающий эти ветви) и нормальная парность их отхождения.
Как следует из самого определения «перитрункальное кольцо», это кольцо распо лагается вокруг артериальных стволов. Отсюда нормальный эмбриогенез ВА предпо лагает следование первичных венечно-артериальных сосудов вокруг аорты и легочно го ствола (но не между ними).
Кроме того, как свидетельствуют материалы, приведенные выше, основу существу ющего строения и ветвления ВА составляют первичные кольца, веерообразно расходя щиеся от места их слияния в ретробульбарной области. Отсюда понятно, что в норме ветви ВА не могут перекрещиваться1 3 . В норме поэтому же исключено соединение дистальных фрагментов веерообразно расходящихся колец.
В норме проксимальные сегменты ветвей-коммуникант через посредство капил лярного сплетения обеспечивают бесперебойное кровоснабжение всех участков сердца и не имеют сообщения с камерами сердца. Дистальные же сегменты этих ветвей реду цируются, составляя основу коллатерального кровообращения. Редуцированные в нор ме участки перитрункального кольца могут сохраняться только при редких вариантах нормы, а также при пороках, анатомические особенности которых не провоцируют их редукцию (тетрада Фалло, например) либо требуют их существования (аномалия от хождения ЛВА от ЛА).
Таким образом, в процессе эмбриогенеза сердца и венечных артерий складывается, как правило, двухкоронарная венечно-артериальная система. При этом обе ВА отходят от лицевых синусов аорты. В норме сохранившиеся фрагменты перитрункального кольца соединяются с проксимальными сегментами всех остальных межсегментных колец первичного венечно-артериального сплетения, причем в различных секторах разворота оси конотрункуса с разными кольцами и не обязательно (хотя чаще) в одина ковом количестве.
В норме в отдельных секторах разворота оси конотрункуса возможно формирова ние однокоронарной системы венечного кровообращения. В норме в сердцах с однокоронарной системой венечного кровообращения все ветви-коммуниканты соединены с системной циркуляцией.
Нормальный эмбриогенез венечно-артериального сплетения и постнатальное раз витие ВА предусматривают возможность самостоятельного отхождения всех главных ветвей от лицевых синусов аорты. При этом все они обязательно соединены (в пределах лицевых синусов аорты) с артериальной циркуляцией.
В норме терминальные ветви ВА не сообщаются с камерами сердца.
Отхождение ВА не от лицевых синусов аорты (эктопическое отхождение от восхо дящей аорты или от ЛС, например), следование ВА между клапанами артериальных со судов или наперекрест друг к другу, формирование однокоронарной системы ВА в не соответствующих секторах разворота оси конотрункуса, отсутствие одной из ветвейкоммуникант или формирование сообщений между ВА и камерами сердца являются отклонениями от нормального формирования сердца и ВА. Поэтому их следует квали фицировать как аномалии.
Таким образом, окончательное формирование двухкоронарного кровообращения подразумевает целый ряд преобразований, сопровождающих формирование сердца.
Присоединение венечно-артериального сплетения к системной циркуляции осу ществляется его перитрункальным кольцом путем пенетрирования эластичной стенки аорты снаружи вовнутрь.
13 В литературе, по данным ангиографии, такие случаи приведены как казуистические (B.Chaitman et al, 1975; L. Muldermans
44

Процесс присоединения перитрункального кольца к артериальному стволу неодно моментен.
На начальных этапах венечно-артериальное сплетение присоединяется только в од ной точке (в области будущего второго лицевого синуса аорты), создавая основу однокоронарного (левого) венечного кровообращения. Двойственное прикрепление венечно-арте- риального сплетения к артериальному стволу (в норме к аортальной его части) осуществ ляется впоследствии, по мере первичной ротации конотрункуса против часовой стрелки.
Растянутость во времени процесса соединения венечно-артериального сплетения и аорты обусловлена меняющимися пространственными взаимоотношениями между ними в процессе формирования сердца.
Первичное соединение сплетения с аортой обусловлено его тесным прилежанием ко второму лицевому синусу аорты на начальных этапах развития этой связи. Двойст венное же, дефинитивное, их соединение завершается по мере присоединения пери трункального кольца к первому лицевому синусу аорты вследствие их сближения по мере переднего перемещения конотрункуса.
Формирование окончательной дефинитивной системы ВА обусловлено нескольки ми процессами и факторами:
1)процессом рассасывания бульбовентрикулярной складки и задним перемеще нием заднего артериального клапана вследствие ротации конотрункуса и вовле чением заднего артериального сосуда в состав левого желудочка;
2)ростом инфундибулума и передним перемещением переднего артериального со суда;
3)рассасыванием перитрункального кольца за задним и впереди переднего арте риального сосуда в сердцах с рассосавшейся бульбовентрикулярной складкой;
4)рассасыванием дублирущих сегментов перитрункального кольца. Особенностью формирования сердец с нерассосавшейся бульбовентрикулярной
складкой является сохранение ретробульбарного фрагмента перитрункального спле тения. При этом в исходной позиции, как правило, исчезают нередкие дублирущие сегменты перитрункального кольца, а при ротации конотрункуса - боковые.
Таким образом, в процессе формирования дефинитивной двухкоронарной (или однокоронарной) системы венечного кровообращения наибольшим изменениям подвер жено перитрункальное кольцо. Проксимальные сегменты остальных колец венечноартериального сплетения в норме независимо от ротационных нарушений в строении конотрункуса изменениям не подвержены: они, как правило, сохранены и соединены нередуцированными фрагментами перитрункального кольца. Дистальные же фраг менты этих колец по мере роста камер сердца редуцируются, составляя потенциаль ный остов коллатерального кровообращения, причем, как показывает опыт, фрагмен ты колец, редуцированные в процессе эмбриогенеза сердца позже, при необходимости функционирования коллатералей, как правило, восстанавливаются первыми (пери трункальное кольцо, дистальные сегменты вокругбульбарных и бульбовентрикулярных колец). Отсюда эффективность коллатералей, в основе формирования которых ле жит восстановление передних (предсердно-правожелудочковых) фрагментов вокругартериальных, вокругбульбарных и бульбовентрикулярных колец.
Как следует из приведенных данных, говорить о фиксированной васкуляризации различных отделов сердца как в норме, так и в сердцах с ВПС неправомочно. И дело здесь не «в значительной функциональной перестройке коронарного русла... при па тологии сердца», как предполагают некоторые исследователи (Д. Г. Иоселиани, 1979), а в особенностях формирования сердца и венечно-артериальной системы.
При сопряженности формирования сердца и ВА, при сложности и многообразии их преобразований в течение этого процесса, а также в силу эмбриональной «заложен ности» дублирования кровоснабжения коммуникантных колец (родоначальников де финитивных ветвей венечно-артериального дерева второго и третьего порядка) пер вичного венечно-артериального сплетения из его интеграционного комплекса понят но, что изменения могут касаться обеих групп ВА.

Характеризуя возможные отклонения в формировании дефинитивной системы ВА, представляется необходимым подчеркнуть, что принципиально следует разли чать две основные группы отклонений: 1) связанные собственно с особенностями фор мирования ВА-системы и 2) связанные с позиционными нарушениями дефинитивной установки конотрункуса.
Изменения первой группы вызваны различиями в области разрыва дистального сегмента коммуникационных колец вследствие роста желудочкового комплекса и во влечением заднего артериального клапана в состав левого желудочка и связанным с этим разрывом перитрункального кольца в ретробульбарной области. Другую под группу в этой группе образуют отклонения, обусловленные сохранением многостволь ное™ перитрункального кольца или характером его присоединения к аорте.
В результате различий в области разрыва интеграционного и коммуникационных колец формируются различные типы доминантного кровоснабжения желудочков и предсердий. Эти же различия ответственны и за различия в формировании васкуляри зации синусного и атриовентрикулярных узлов. Таким образом, в основе различий ва скуляризации основных водителей ритма и отделов сердца лежат различия в месте разрыва первичных колец фетального венечно-артериального сплетения.
Одновременно отклонения в интеграционном комплексе (сохранение многостволь ное™ его ретробульбарного сегмента, соединение с аортой в точках, расположенных до слияния этих колец) могут в норме привести к отхождению ВА не от соответствую щих синусов аорты (транслокация устьев с сохранением редуцированных в норме стволов интеграционного комплекса) либо к отхождению ВА несколькими устьями или же совмещению обеих особенностей.
Как уже отмечалось, формирование второй группы отклонений (вариантов) связа но с различным разворотом конотрункуса. Так как в зависимости от угла разворота оси конотрункуса формируются постоянные варианты соединения интеграционного коль ца с коммуникационными, в соответствующих секторах разворота оси конотрункуса эти варианты строения венечно-артериального дерева также являются нормальными.
Нам представляется, что приведенные материалы имеют двоякое значение: 1) по зволяют формировать наши представления о нормальности венечно-артериального де рева и 2) имеют непосредственно прикладное значение.
Понимание того, что ОВ правой ВА, ОВ левой ВА, ПМЖВ, ДВ, заднебоковая ветвь ОВ и ЗМЖВ представляют собой равноценные фрагменты колец первичного венечноартериального сплетения позволяет отвлечься от принципа соподчиненности стволов и ветвей ВА и их деления на ветви первого, второго и других порядков, широко ис
пользуемого |
в ортодоксальной |
анатомии (Б.В.Огнев и др., 1954; В.В.Кованое и |
Т. Н.Аникина, |
1974; С. С. Михайлов, |
1987). Правильная оценка этого факта определя |
ет анатомическую необходимость полной реваскуляризации (всех вышеперечислен ных ветвей) миокарда при ИБС. Понимание того, что указанные артериальные стволы представляют собой нередуцированные сегменты веерообразно расходящихся (и по этому несоединенных в фетальной жизни, а значит и в постнатальной) колец раскры вает отсутствие возможности существования эффективной субэпикардиальнои связи (коллатералей или иных соединений) между ними. Поэтому проксимальные сужения в этих ветвях требуют обязательной реваскуляризации.
Как известно, адекватная реваскуляризация миокарда в настоящее время является
основополагающим принципом |
аортокоронарного шунтирования (В.И.Бураковский |
и др., 1985; В. С. Работников и др., |
1986; Г. В. Кнышев и др., 1986; В. И. Урсуленко и др., |
1991; Л. А. Бокерия и др., 1997-2000; В. Barratt-Boyes et al., 1984; D. Cosgrove et al, 1984; J. Kirklin et al, 1985; G. Lawrie et al., 1987; R. Zeff et al., 1988; P. Davis et al., 1989). Одна ко формирование такой точки зрения было основано на материалах изучения дисталь ного русла ВА и на оценке результатов аортокоронарного шунтирования. Хотя была раз личная ценность основных стволов ВА в васкуляризации миокарда (по локализации в них патологического процесса, W.Roberts, 1976), эмбриологическая и анатомическая сущность этих различий до настоящего времени не раскрыта. Концепция полной ревас-
46