

Глава 6
СВОЙСТВА ЗВЕНЬЕВ ПРОЦЕССА
ВОЗБУЖДЕНИЯ В МЕХАНОРЕЦЕПТОРАХ
В предыдущих главах были описаны некоторые общие характеристики деполяризационных локальных и распространяющихся ответов (РП и ПД) механорецепторов (главы 2 и 3), а также показана роль различных структур механорецепторпых приборов в развитии адаптации (глава 4). В данной главе будут рассмотрены свойства отдельных звещьев той цепи процессов, которая приводит к возникновению возбуждения в механорецепторах при действии на них адекватных стимулов. Анализ свойств указанных звеньев проводился в литературе различными методами, с помощью различных физических и химических факторов, таких как электрический ток, температурные воздействия, разнообразные вещества, имеющие разные точки приложения в ткани, ионы и т. д. Некоторые характеристики указанных звеньев будут затрагиваться также и в главе 11, посвященной механизмам действия механического раздражения.
В литературе накопилось чрезвычайно много работ, посвященных действию различных факторов на механорецепторы, однако подавляющее большинство такого рода исследований было выполнено при регистрации импульсной (часто даже суммарной) активности рецепторов. Лишь сравнительно небольшое число экспериментов, преимущественно осуществленных за последнее десятилетие, было выполнено на одиночных рецепторах с регистрацией РП. Естественно, что при изучении свойств звеньев процессов, протекающих в механорецепторах при возбуждении, наибольший интерес представляет исследование процессов трансформации и процессов генерации РП. Существенным является также исследование характеристик регенеративной системы генерации, хотя реакция этой системы на действие тех или иных факторов может быть более легко понята на основе хорошо известных из литературы фактов, касающихся механизмов функционирования регенеративных мембран различных нервных проводников и клеток.
Деятельность механорецепторов во многом зависит от механических характеристик их вспомогательного аппарата и окружаю- 268
щих тканей (элементов кожи, мышц, стенок сосудов и т. д.). На роль этих структур в деятельности некоторых рецепторов специально обращалось внимание при рассмотрении процессов адаптации (глава 4). Вспомогательные аппараты механорецепторов чрезвычайно разнообразны. Описанию их свойств посвящено большое число специальных, относящихся именно к данному виду механорецепторных приборов исследований (см., например, обзоры, касающиеся деятельности вестибулярного аппарата и органа слуха, в томе данного руководства, посвященном физиологии сенсорных систем: Кисляков и др., 1972; Молчанов, Радионова, 1972). Специфика^свойств вспомогательных аппаратов различных механорецепторов не позволяет рассматривать их в рамках данной книги. По этим вопросам следует обращаться к специальным работам.
При использовании физических и химических факторов для изучения процессов, протекающих в механорецепторах, крайне важно правильно оценивать точку приложения действующего агента. Наличие в литературе не всегда обоснованных построений и противоречивых фактов, относящихся к этой области исследований, определяется прежде всего многокомпонентным действием некоторых используемых агентов. Е с ли J оценивать деятельность механорецепторов по изменению' их импульсной активности под влиянием физических и химических факторов, то местом приложения действующего агента могут быть следующие структуры.
Сосуды данной рецепторной области. Так может действовать, например, подводимый через сосудистое русло адреналин. Наступает спазм сосудов, следствием чего является развитие асфиксии, а в результате этого — угнетение деятельности рецептора (Paintai, 1959; Eldred, Schnitzlein, Buchwald, 1960; Calma^ Kidd, 1962).
Вспомогательный аппарат рецептора, а также окружающие его тканевые структуры. Примером такого действия может служить влияние на мышечные веретена, кожные механорецепторы и др. различных химических веществ: ацетилхолина (Hunt, 1952), сукцинилхолина (Granit et al., 1953; Rack, Westbury, 1966; Burgess, Clark, 1969b), никотина (Verhey, Voorhoeve, 1963), адреналина (Douglas, Gray, 1953; Witzleb, 1953; Diamond, 1955), т. e. таких веществ, которые могут вызывать сокращение окружающих рецепторы или входящих в их состав контрактильных элементов.
Механочувствительный субстрат рецептора, ответственный за восприятие адекватного раздражителя, а также связанная с ним система генерации РП. Например, под влиянием температурных изменений и при действии постоянного тока меняются амплитуда и скорости нарастания РП рецепторов растяжения у ракообразных и телец Пачини (Burkhardt, 1959; Loewenstein, Ishiko, 1960; Inman, Peruzzi, 1961; Ishiko, Loewenstein, 1961; Ильинский и др., 1965; Ильинский, Акоев, 1966).
Регенеративная система генерации, обеспечивающая появление ПД. Примером такого избирательного действия на эти структуры может служить влияние тетродотоксина на тельца Пачини и рецепторы растяжения у" ракообразных (Loewenstein et al., 1963; Ozeki, Sato, 1965; Nishi, Sato, 1966, 1968). Следует указать, что очень многие эффекты, связанные с влиянием физических и химических факторов на рецепторы (Paintai, 1964, 1971. 1972), обусловлены именно преимущественным действием на регенеративную систему генерации (рис. 95).
Возрастание
устойчивости к действию химических
веществ, охлаждению и аноксии
Миелинизированное
волокно
Летучие
наркотики, вератрин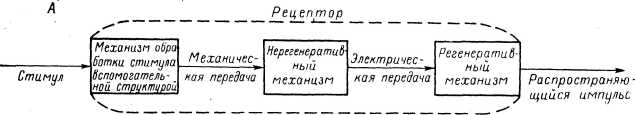



Область регенератов- Область нерегенеративной системы
Немиелинизированное
волокно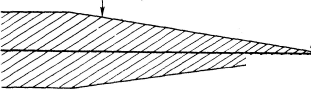

Область регенеративной Область нерегенератив- системы ной системы
f I I
Ацетилхолин, серотонин, гистамин, вератрин, летучие наркотики и др
Рис. 95. Чувствительность отдельных систем первичного рецептора к действию различных факторов среды (по: Paintai, 1972).
А — механизмы, участвующие в процессе возбуждения в первичном механорецепторе, и их относительная устойчивость к действию факторов среды. В — точки приложения действия различных химических веществ (область регенеративной системы, т. е. место генерации ПД, где диффузионные барьеры минимальны).
5. Эфферентные нервные окончания и связанные с ними постсинаптические структуры тех или иных элементов рецепторного прибора. Так влияют, например, химические агенты, имитирующие действие медиаторов, выделяющихся из эфферентных нервных окончаний: ацетилхолина в гамма-системе в случае мышечных веретен (Granit et al., 1953) или в случае волосковых вторичных механорецепторов, например в органах боковой линии (Russell, 1968, 1971b; Flock, Russell, 1973).
Очевидно, что может иметь место и комбинированное действие исследуемых факторов на различные структуры рецептора и окружающие его ткани, например при действии температуры и постоянного тока. Обычно такого рода сложные влияния наблюдаются и при использовании больших доз химического раздражителя, например адреналина (Paintai, 1964). У вторичночувствующих механорецепторов благодаря наличию рецептирующей клетки и, следовательно, синаптического контакта между этой клеткой и сенсорным нейроном имеются по сравнению с первичночувствую- щими структурами еще и другие возможные точки приложения физических и химических раздражителей. Такое положение дел требует осторожного выбора средств для анализа свойств процессов, протекающих в механорецепторах, равно как и осторожного истолкования полученных результатов. Следует учесть, что многочисленные существующие в литературе данные по действию различных физических и химических факторов на механорецепторы далеко не всегда были получены в опытах, направленных на выяснение механизмов деятельности самих механорецепторных приборов. Не всегда учитывалось и многокомпонентное действие используемыхТагентов.
Характеристики ответов механорецепторов при действии электрического тока
Физиологам давно известно, что применение электрического тока является весьма действенным методом анализа свойств различных возбудимых структур. Важные факты по действию электрического тока были получены и в опытах на рецепторных элементах, в том числе и на механорецепторах (Matthews, 1931а; Gatton, 1958; Lippold et al., 1960a; Ishiko, Loewenstein, 1961; Terzuolo, Washizu, 1962; Ильинский, Акоев, 1966; Nakajima, Onodera, 1969a, 1969b, и др.). При этом было обнаружено, что часть рецепторных структур, входящих в состав органов боковой линии рыб, обладает чрезвычайно высокой чувствительностью к действию электрического тока (специализированные органы боковой линии). Это позволило рассматривать их как электрорецепторы (см. обзорные работы: Lissmann, 1958; Lissmann, Machin, 1958; Machin, 1962; Dijkgraaf, 1963; Bennett, 1965, 1967, 1970, 1971; Murray, 1965a, 1967; Lissmann, Mullinger, 1968; Акоев и др., 1971; Протасов, 1972, и др.). Многочисленные морфологические и физиологические данные позволяют считать, что развитие электрочувствительности специализированных сенсорных структур боковой линии произошло на основе рецепторной клетки, обладающей высокой механочувствительностью (волосковой сенсорной клетки) (см. данные морфологии в главе 1). Частью рецепторов специализированных органов волоски были затем утрачены, в других же, таких как ампулы Лоренцини, они сохранились (глава!, рис. 16). Ряд таких рецепторов, как уже отмечалось ранее, обладает высокой чувствительностью к действию механических стимулов, что даже позволило исследователям (Loewenstein, I960; Murray, 1960; Szabo, 1970, 1972) рассматривать их как механорецепторы. (Интересно заметить, что, как и другие типичные механорецепторы, сенсорные элементы ампул Лоренцини возбужда
ются при действии растяжения—деформации,— а не давления — см. главу 11, рис. 121). В этой связи в данном разделе представляется целесообразным рассматривать действие электрического тока не только на «типичные» механорецепторы боковой линии (невро- маеты), но и на электрорецепторные образования (рецепторы «особых», специализированных, органов боковой линии).
Работ, посвященных действию электрического тока на активность механорецепторов, сравнительно немного. Эти исследования проводились в двух основных направлениях: изучалось влияние электрического тока на ритмическую активность, а кроме того на процессы генерации РП. Общим итогом работ первого направления явилось установление того факта, что в различных тонических механорецепторах катод постоянного тока увеличивает импульсную активность, а анод ее угнетает (Katsuki, Yoshino, 1952; Та- saki, Fernandez, 1952; Edwards, 1955; Lowenstein, 1955; Murray, 1956; Catton, 1958; Terzuolo, Washizu, 1962).
Этот вывод хорошо согласуется с представлением о том, что генераторный потенциал деполяризует отходящее от рецептора нервное волокно и вызывает тем самым возникновение в нем импульсной активности. Однако следует сразу указать, что продолжительный деполяризующий ток достаточной силы может не только способствовать усилению импульсной активности механорецепторов, но и угнетать ее. Это определяется аккомодационными свойствами регенеративной системы генерации. В естественных условиях, когда под влиянием адекватного раздражения в рецепторе возникает деполяризующий ток (лежащий в основе деполяризационного РП), именно аккомодационные свойства, как отмечалось в главе 4, являются одним из решающих факторов, определяющих его адаптационную способность. Использование электрического тока для целей анализа явлений адаптации в механорецепторах, равно как и в других рецепторах, неоднократно встречается в литературе (см. главу’*4).
Сложнее изучать действие электрического тока на генерацию РП механорецепторов. В литературе существуют лишь немногочисленные специальные работы такого рода, выполненные прежде всего на традиционных объектах исследования (тельцах Пачини, мышечных веретенах, рецепторах растяжения у ракообразных).
Действие электрического тока на первичные рецепторы
Влияние постоянного электрического тока на тельца Пачини. Было установлено (Loewenstein, Ishiko, 1960; Ильинский, Акоев, 1966), что при действии катода на нервное окончание амплитуда РП уменьшается по сравнению с нормой, а скорость нарастания РП падает. При действии анода величина РП и скорость нарастания ответа увеличивались. Ток любого направления значительно увеличивал флуктуацию РП. Действие
постоянного тока проявлялось практически сразу после его включения. Кривая зависимости амплитуды локального ответа от силы адекватного раздражения при поляризации смещалась вдоль оси амплитуды РП таким образом, что каждое значение РП в нормальных условиях как бы умножалось на некоторую величину (зависящую от силы тока): больше единицы при действии гипер- поляризующего тока и меньше единицы при действии деполяризующего тока (рис. 96).
В настоящее время имеются все основания полагать, что под влиянием надпорогового механического стимула в механочув- ствительной мембране телец Пачини происходят структурные перестройки, следствием которых тивления и ионной проводимости. В свою очередь, в результате этого меняются трансмембранные ионные токи, что и регистрируется в виде* РП. Следовательно, величина локального ответа при прочих равных условиях будет опре-
Рис. 96. Влияние постоянного тока на рецепторный потенциал телец Пачини (Ильинский, Акоев, 1966).
По оси абсцисс — сила раздражения (R, усл. ед.); по оси ординат — амплитуда РП (V, мкв). 7V— отсутствие поляризации; 4-2 >10-7 а и —2‘10~7 а — сила гипер- и деполяризующего тока. Значения амплитуды РП при гиперполяризации и деполяризации могут быть получены умножением амплитуды в отсутствие поляризации (N) на коэффициенты 1.4 и 0.7 соответственно.
является
изменение ее сопро-

Процесс, лежащий в основе структурных преобразований механорецепторной мембраны под влиянием адекватного стимула, обозначают как процесс трансформации. Ионные токи, возникающие в результате изменения свойств поверхностной мембраны (как отмечалось выше, — стр. 74), обозначают как процесс генерации РП (Ильинский, 1964а, 1966д). Таким образом, между внешним стимулом и РП существуют две группы явлений: одна, связанная с процессом трансформации, а другая — с процессом генерации. Для целей изучения этих явлений их нужно каким-то образом разделять.
Несомненно, что для преодоления сил, сохраняющих покоящуюся структуру мембраны нервного окончания, требуется при- дожить некоторое пороговое усилие, как бы мало оно ни было. Можно думать, что величина этого усилия в какой-то мере характеризует процесс трансформации. Измерить эту величину непосредственно пока не удается, однако об относительных изменениях ее можно составить представление на основании рассмотрения порога возникновения РП (/?0). Пороговая сила раздражения зависит от двух факторов: от механических свойств мембраны нервного окончания, характеризующих собой процесс трансформации, а также от механических свойств вспомогательного аппарата. Поэтому оценивать исключительно процессы трансформации по характеру изменений при том или ином воздействии нельзя, тем более что эта величина может быть крайне малой. Значительно более надежно судить о нем на основании изменения всей кривой зависимости амплитуды РП от силы раздражения. Кроме того, известную характеристику процессов трансформации можно получить на основании анализа величины 7?тах, т- е- наименьшей силы раздражения, при которой достигается максимальный локальный ответ Утах.
Амплитуда РП при постоянной силе раздражения может меняться под влиянием того или иного воздействия в результате изменения сил, сохраняющих покоящуюся структуру рецепторной мембраны, а также вследствие изменения электрохимических градиентов. Если приложить к рецептору стимул, амплитуда которого заведомо больше, чем необходимая для максимального ответа, то в этом случае изменения механической составляющей процесса трансформации (так же как и изменения в механических свойствах вспомогательного аппарата) не должны сказываться на величине локального ответа. Амплитуда ответа будет теперь зависеть исключительно от величины электрохимических градиентов. Поэтому, изучая изменения именно величины Fmax под влиянием различных воздействий, можно составить представление об этих градиентах, определяющих процессы генерации РП (Ильинский, 1966а, 1966г, 1966д).
Как следует из результатов опытов по действию постоянного тока на тельца Пачини, кривая зависимости амплитуды РП от силы раздражения смещается в область более значительных амплитуд РП при гиперполяризации и в сторону меньших значений РП при деполяризации. Так как при этом исходное значение РП, соответствующее каждой данной силе раздражения, как бы умножается на некоторую величину (постоянную для каждой силы поляризующего тока), то можно говорить о неизменности величин Ro и /?тах (рис. 96). Следовательно, можно полагать, что электрический ток не оказывает влияния на процессы трансформации в тельцах Пачини. Напротив, в опытах с поляризацией величина Утах изменяется значительно, что может говорить о зависимости генерации РП, т. е. следующего звена в цепи процессов возбуждения, от действия постоянного тока (Ильинский, Акоев, 1966).
Данные, полученные на тельцах Пачини, интересно сопоставить с результатами экспериментов на мякотных и безмякотных нервных волокнах, которые раздражались механическим стимулом (Julian, Goldman, 1962). Было установлено, что при такого рода раздражении возникала локальная деполяризация, которая могла перерастать в спайк. Развитие деполяризации сопровождалось падением сопротивления мембраны. Дополнительно к этому на волокнах было установлено, что увеличение проводимости при механическом раздражении наблюдалось даже в случае, если развитие деполяризации предупреждалось электрическими методами. Уменьшение электрического сопротивления мембраны, вызванное механическим стимулом, было много больше, чем при электрическом раздражении, приводящем к развитию деполяризации, равной по амплитуде. Такого рода факты дают основания предполагать осуществление каких-то структурных перестроек в возбудимой мембране нервного волокна, обусловленных механическим раздражением и отличных от тех структурных изменений, которые могут сопровождать первичные изменения мембранного потенциала. Таким образом, данные этих экспериментов хорошо согласуются с результатами, полученными на тельцах Пачини.
Известно, что если импульсная активность в тельцах Пачини заблокирована (например, новокаином или тетродотоксином), то возникающий в немиелинизированной части нервного окончания локальный ответ распространяется вдоль рецептора пассивно (Loewenstein, 1961а, 1965). Было показано (рис. 27), что амплитуда РП падает по экспоненциальному закону по мере удаления от точки раздражения. Так же изменялась амплитуда электрического потенциала, наносимого с помощью микроэлектрода на декапсулированное нервное окончание и измеряемого в разных точках от места нанесения. Все эти данные говорят о том, что возбуждение рецепторной мембраны в тельцах Пачини является локальным процессом. Соседние с возбужденным участком области мембраны не вовлекаю!ся локальными токами в процесс возбуждения. Итак, на основании изложенных данных можно утверждать, что элементы механорецепторной поверхности в тельцах Пачини, ответс1венные за восприятие адекватного раздражения, являются электрически невозбудимыми.
Это же подтверждается следующим. Из литературы хорошо известно, что мембраны, не возбуждающиеся электрическим током, могут генерировать реверсные локальные ответы на адекватное раздражение при соответствующих изменениях величины мембранного потенциала клетки (см., например: Eccles, 1957, 1964; Grundfest, 1959, 1961а; Bennett, 1961; Костюк, 1962). Можно было ожидать, что в тельцах Пачини также воспроизводится указанное явление, если их рецепторная мембрана электрически невозбудима. Это и было доказано в соответствующей работе (Ильинский, Акоев, 1966).
Таким образом, реакция рецепторной мембраны нервного окончания тельца Пачини на действие электрического тока оказалась принципиально такой же, как у различных электроневоз- будимых постсинаптических и рецепторных структур позвоночных и беспозвоночных животных. Такого рода аналогии касаются, например, нейронов центральной (Coombs et al., 1955) и периферической (Hagiwara et al., 1959; Nishi, Koketsu, 1960) нервной системы, мышечных волокон (Fatt, Katz, 1951; Castillo, Katz, 1954; Burke, Ginsborg, 1956; Takeuchi, Takeuchi, 1959, 1960b), фоторецепторов (Fuortes, Poggio, 1963; Walther, 1965; Millecchia, Mauro, 1969b; Baylor, Fuortes, 1970; Brown et al., 1970; Wulff, 1971), других видов механорецепторов (Kuffler, Eyzaguirre, 1955; Hagiwara et al., 1960; Jansen et al., 1971b), хеморецепторов (Ozeki, 1971) и мн. др.
Изучение действия постоянного тока на генерацию ПД телец Пачини показало, что при гиперполяризации амплитуда и скорость нарастания спайков возрастала, а при деполяризации — падала (Loewenstein, Ishiko, 1960). Критический уровень локальной деполяризации, необходимый для генерации ПД, возрастал при действии как входящего, так и выходящего токов. Порог возникновения ПД при механическом раздражении рецептора возрастал под влиянием деполяризации, снижался при относительно небольших силах гиперполяризующего тока и возрастал при его увеличении. Снижение порога генерации ПД наблюдалось лишь в диапазоне интенсивностей гиперполяризующего тока, при которых наблюдавшееся увеличение амплитуды РП превалировало над повышением критического уровня деполяризации. Тот факт, что деполяризующий ток вызывал затруднения в возникновении импульсной активности в тельцах Пачини, следует связать с особенностями регенеративной системы генерации ПД в этих рецепторах. Тельца Пачини не могут стационарно генерировать импульсные разряды при длительной деполяризации: наблюдается быстрая инактивация регенеративных механизмов. Это, как было показано в главе 4, существенно отличает фазные рецепторы (крайним вариантом которых являются тельца Пачини) от тонических рецепторных приборов, в которых длительная исходная деполяризация достаточной интенсивности обеспечивает стационарный процесс генерации спайковой активности.
Влияние электрического тока на мышечные веретена. Действие постоянного электрического тока на РП этих рецепторов изучалось в экспериментах на мышечных веретенах лягушки (Алексеев, 1973а). Результаты этого исследования оказались принципиально сходными с данными, полученными на тельцах Пачини. Под влиянием деполяризующего тока наблюдалось уменьшение амплитуды РП, а при действии гиперполяризующего тока — ее увеличение. Статическая фаза деполяризационного РП, а также гиперполяризация, возникающая при снятии механического раз
дражения, изменялись в большей степени, чем динамическая составляющая ответа, особенно при небольших величинах растяжения.
Под влиянием постоянного тока кривые зависимости амплитуды динамической и статической фаз деполяризационного РП смещались вдоль оси амплитуды РП так, что каждое значение на кривой в нормальных условиях следовало умножить на некоторую (одинаковую для данной используемой силы поляризующего тока) величину, превышающую единицу при гиперполя- ризующем воздействии или же меньше единицы при деполяризующем влиянии, т. е. величины Но и 7?юах не изменялись при поляризации. Латентный период и скорость нарастания РП заметно не изменялись под влиянием постоянного тока. При больших значениях деполяризующего тока наблюдалось извращение знака, реверсия РП. На основании полученных данных был сделан вывод, что механочувствительный субстрат мышечного веретена является электрически невозбудимым. Это заключение полностью согласуется с рассмотренными выше данными, относящимися к тельцам Пачини.
Изменение импульсной активности мышечного веретена оказалось сходным в опытах различных исследователей (Edwards 1955; Lippold et al., 1960a, 1960b; Emonet-Denand, Houk, 1969; Ito, 1970; Алексеев, 1973a): под влиянием деполяризации частота фоновой импульсации возрастала, амплитуда ПД снижалась; при действии же гиперполяризации отмечалось уменьшение частоты ПД, а амплитуда спайков увеличивалась.
В ряде работ рассматривалась роль аккомодационных особенностей регенеративной системы генерации в механизмах адаптации мышечных веретен (Lippold et al., 1960а; Emonet-Denand, Houk, 1969).
Влияние электрического тока на рецепторы растяжения у ракообразных. Исследование действия электрического тока на рецепторы растяжения проводилось в ряде работ. Основной задачей этих исследований было изучение явлений адаптации. При этом с помощью электрического тока оценивались аккомодационные характеристики регенеративной системы генерации ПД (Wendler, Burkhardt, 1961; Nakajima, 1964; Nakajima, Onodera, 1969a, 1969b). Подробнее данные работы рассматривались в главе 4. В этих исследованиях было установлено не только различие в аккомодационных характеристиках регенеративных систем генерации медленно и быстро адаптирующихся рецепторов, но и различие в свойствах разных участков аксона МРОХ (см. рис. 70).
Опыты с электрическим раздражением различных отделов рецептора растяжения, а также с антидромной стимуляцией его аксона показали (Grampp, 1966), что разветвления дендритов сенсорного нейрона, деформация которых и ведет к развитию ГП, являются, по-видимому, электрически невозбудимыми. Нерегенеративный характер электрических процессов в этой об
ласти отмечают и другие исследователи (Klie, Wellhoner, 1973), использовавшие метод фиксации потенциала.
Влияние электрического тока на другие механорецепторы» Систематического изучения действия электрического тока на РП других механорецепторов не проводилось. Отдельные заслуживающие внимания факты, полученные методиками внутриклеточ- ных’отведений и касающиеся действия этого фактора на волосковые рецепторные клетки органов чувств беспозвоночных (Alkon, Bak, 1973; Detwiler, Alkon, 1973), приводились ранее (стр. 100, 183).
* ♦ ♦
Эксперименты на тельцах Пачини, мышечных веретенах и рецепторах растяжения позволяют говорить об электрической не- возбудимости механорецептирующего субстрата. В настоящее время имеется уже довольно обширная литература, касающаяся электрической невозбудимости постсинаптических и рецепторных структур (Kuffler, 1942; Fatt, Katz, 1951; Castillo, Katz, 1954; Grundfest, 1957, 1959, 1961a, 1961b, 1965, 1971; Hagiwara, 1961; Werman, 1963; Eccles, 1964; Baylor, Fuortes, 1970, и мн. др.). Согласно концепции, развиваемой прежде всего работами Грунд- феста и его сотрудников, воспринимающие поверхности клеток (постсинаптические мембраны и рецептирующие структуры) активируются лишь специфическими для них раздражителями. Электрический же ток, который генерируется при возбуждении этого участка мембран, действует лишь на другие, электрически возбудимые части клеток. Первоначально предполагалось, что вся рецептирующая поверхность клетки является электрически невозбудимой. Однако, по-видимому, более правильно говорить в этом случае о собственно воспринимающих зонах мембраны, размеры которых могут ограничиваться даже молекулярным диапазоном. По-видимому, если восприятие внешнего раздражения в рецепторах начинается с трансформационных процессов, то именно способность мембранных участков к этим процессам можно считать как-то связанной с электрической невозбуди- мостью субстрата. Электрически невозбудимые и возбудимые микроучастки мембраны могут чередоваться (см.: Grundfest, 1971). В таких случаях, по мнению некоторых авторов (Spyropou- los, Tasaki, 1960; Eccles, 1964), кажется невозможным доказать или отвергнуть точку зрения Грундфеста и других, согласных с его воззрениями авторов.
Электрическую невозбудимость воспринимающих элементов клеток можно отнести к одному из общих их свойств. В отношении рецепторных приборов следует отметить, что начиная с процессов генерации РП явления в различных рецепторах (фото-, хемо-, механорецепторах и т. д.) если не совсем идентичны, то все же имеют много общих черт (Davis, 1961; Ильинский, 1966д, 1967; Fuortes, 1971). Процессы же взаимодействия внешнего раздражителя с рецептирующим субстратом очень специфичны и в силу этого должны существенно отличаться у разных рецепторов. Вместе с тем очевидным сходством этих различных процессов трансформации в разных рецепторах является обсуждавшаяся здесь электрическая невозбудимость рецептирующего субстрата.
Действие электрического тока на вторичные рецепторы
Влияние электрических стимулов на рецепторные структуры системы органов боковой линии. «Обычные» о р- ганы (система невромастов). Первые электрофизиологические исследования влияния электрического тока на рецепторы боковой линии неэлектрических рыб были осуществлены на японском угре (Katsuki et al., 1951а, 1951b, 1951с; Katsuki, Yoshino, 1952).1 Нервные волокна от механорецепторов боковой линии, по данным этих авторов, весьма варьировали по толщине . (от 4 до 15 мкм) и различались по функциональным особенностям. Так, тонкие волокна обладали значительной спонтанной активностью, а толстые обычно ее не имели. Тонкие волокна характеризовались более низкими порогами к действию тока, а максимальная частота их разрядов (40 в 1 сек.) была значительно ниже, чем у толстых волокон. Пороговые силы тока, оказывавшие влияние на рецепторы, были весьма значительны (30 мка и более). Обычно под влиянием постоянного входящего тока (положительный электрод на поверхности кожи, а отрицательный — в теле животного) наблюдалось возбуждение рецепторов, а при действии выходящего тока — торможение. Выключение раздражения вызывало обратные эффекты. Вместе с тем встречались волокна, активность которых изменялась противоположным образом.
Тонкие волокна генерировали тонический ответ, а толстые — фазный. Частота импульсов в ответе возрастала по мере увеличения крутизны нарастания раздражающего тока. Использование переменного тока приводило к появлению активности соответственно каждому циклу воздействия. Толстые волокна в этих условиях обладали способностью отвечать на раздражения большей частоты, чем тонкие. Некоторые функциональные особенности тонких и толстых волокон, по-видимому, были связаны с их морфологическими различиями (характером разветвлений и окончаний на рецепторных, структурах.)
Сопоставление действия электрического тока на «обычные» и «особые» органы боковой линии слабоэлектрической рыбы
1 В последние годы в литературе начали появляться работы, посвященные изучению свойств волосковых механорецепторов боковой линии с помощью внутриклеточных микроэлектродов (Flock, Russell, 1973; Flock et al., 1973a, 1973b). В этих работах начато систематическое изучение электрических характеристик волосковых клеток. Основные относящиеся к этой области факты были приведены ранее (стр. 94).
Gymnolus carapo показало (Suga, 1967a, 1967c), что при достаточной интенсивности раздражения в волокнах, отходящих от невро- мастов, можно было отметить появление импульсации, сохраняющейся на протяжении всего времени действия стимула. Однако поверхностно и глубоко лежащие невромасты обладали значительно меньшей чувствительностью к воздействию электрического стимула, чем специализированные образования.
При раздражении импульсами длительностью 0.5—1.Q мсек, (соответствующими продолжительности собственных разрядов электрического органа рыбы) «обычные» органы не реагировали даже при градиенте электрического стимула более 400 мв/см2. Разряды собственных электрических органов также не изменяли импульсной активности механорецепторов боковой линии. Сходные данные были получены и на сильноэлектрических скатах Torpedo (Акоев и др., 1975), мощные разряды электрического органа которых, вызывавшие значительные изменения активности электрорецепторов ампул Лоренцини, были неэффективны по отношению к «обычным» механорецепторам боковой линии. Эти результаты представляют интерес в связи с рассмотрением роли механо- и электрорецепторных структур боковой линии в моменты обороны и нападений (сравн.: Belbenoit, 1970а, 1970b). На различия в чувствительности к электрическому току «обычных» и специализированных органов боковой линии указывали и другие авторы (Hagiwara, Morita, 1963; Hagiwara et al., 1965).
«Особые» (специальные) органы. Первые доказательства высокой чувствительности этих структур к действию электрического тока были получены давно в экспериментах по изучению поведения (Parker, Van Heusen, 1917). Однако лишь после работ Лиссмана (Lissmann, 1951, 1958, 1961, 1963; Lissmann, Machin, 1958), показавшего способность некоторых рыб ориентироваться в пространстве и различать объекты, основываясь исключительно на электрических свойствах среды, «особые» органы боковой линии стали предметом систематических электрофизиологических исследований. К настоящему времени накопилась обширная литература по данному вопросу (см. обзоры: Dijk- graaf, 1963; Bennett, 1970, 1971; Акоев и др., 1971; Протасов, 1972).
Наиболее существенной отличительной чертой реакции специализированных органов боковой линии на действие электрического тока является их очень высокая чувствительность, намного превышающая электрочувствительность «обычных» структур. Так, пороговая интенсивность стимуляции током ямковых органов у карликовых сомиков равняется 10“9—10“12 а/мм2 (Roth, 1968; Peters, Buwalda, 1972; Андрианов, Ильинский, 1973). Сходные данные (10“10—10"11 а/мм2) были получены для рецепторов ампул Лоренцини у скатов (Murray, 1962; Акоев, Жадан и др., 1974; Андрианов и др., 1974), сенсорных структур у мормирид (Bennett, 1965) и др.
По характеру ответов рецепторы специализированных органов могут быть подразделены на фазные и тонические. К фазным относятся рецепторы бугорковых органов, а к тоническим — ампулированных (Bennett, 1967; Suga, 1967b). Узор разрядов электрорецепторов при действии электрического тока чрезвычайно разнообразен. По-видимому, по количеству способов кодирования информации электрорецепторы стоят на первом месте среди различных сенсорных структур (Акоев и др., 1971).
Детальное рассмотрение свойств специализированных (элек- трорецепторных) органов боковой линии выходит за рамки дан-
1
—
■МШИ]In
| llllllllllllllj
и iiiiimiimn и ■
'-IN!iMllillIliiBliiliiiiliilliy^. ||||MilHhllllJllt ' Пфмм III iifiMHiiiiHiii
4

Рис. 97. Действие механического раздражения на активность рецепторов ампул Лоренцини ската Dasyatis pastinaca.
1 — фоновая активность рецепторов; 2—4 — изменение активности под влиянием водной струи, направленной в область ампул; 5 — активность рецепторов при прикосновении к области выхода ампулярных каналов в коже. I, 2 — при естественных ритмических дыхательных движениях животного; 3—5 — при искусственном непрерывном пропускании воды через жаберные щели. Сверху вниз: на 1—2 — биоэлектрическая активность; движения жаберных крышек; отметка раздражения; на 3—5 — биоэлектрическая активность; отметка раздражения. Отметка времени — 0.5 сек.
ной книги. Поэтому здесь будут вкратце изложены лишь некоторые данные, касающиеся только особенностей^функционирования таких рецепторов, которые обладают также и высокой механической чувствительностью. К их числу, как уже отмечалось, следует прежде всего отнести рецепторы ампул Лоренцини, имеющихся у пластиножаберных рыб (акул, скатов) (см. главу 1, рис. 16). Механическая чувствительность этих образований хорошо выявляется в различных условиях эксперимента (рис. 97). Интересно, что ответы рецепторов ампул Лоренцини на механическое воздействие четко выявляются даже при нанесении раздражения в области выхода ампулярных каналов, т. е. на большом расстоянии (10—15 см и более) от самих рецепторов (Акоев и др.,
1975). Кроме того, получены данные (Красникова, 1974), говорящие о том, что в желе ампул Лоренцини при действии давления возникают так называемые потенциалы смещения (см. стр. 335), происхождение которых обусловлено полиэлектролитным характером кислых мукополисахаридов (гликозаминогликанов), содержащихся в большом количестве в ампулярном желе (см. также: Doyle, 1963, 1967, 1968; Denizot, 1969, 1970). Возникновение подобных потенциалов позволяет предположить их причастность к механизмам возбуждения в ампулярных механорецепторных структурах.
Рецепторы ампул Лоренцини обычно даже в отсутствие внешнего раздражения находятся в состоянии активности, носящей постоянный характер у рыб при искусственном непрерывном пропускании аэрированной морской воды через жаберные щели. У животных с естественными ритмическими дыхательными движениями пачки импульсов активности четко коррелируют с темпом дыхательных движений (рис. 98) и обусловлены раздражением рецепторов токами, возникающими при мышечных сокращениях (Dijkgraaf, Kalmijn, 1966; Katsuki, Hashimoto, 1969; Акоев, Ильинский, 1972, 1973; Акоев и др., 1975). Аналогичного рода наблюдения были сделаны и на других электрорецепторах (Peters, Buwalda, 1972). Механорецепторы «обычных» органов боковой линии в этих же условиях генерируют спонтанную активность, которая обычно не модулируется в ритме с дыхательными движениями. Общие движения тела животного вызывают появление активности у рецепторов обоих типов (сравн.: Roberts, 1972, и Акоев и др., 1975). Механизмы же их активности, по-видимому, различны.
Электрические стимулы оказывают четко выраженное действие на рецепторы ампул Лоренцини. Обычно катодические стимулы обладают возбуждающим действием, а анодические — тормозящим (Murray, 1962, 1965а, 1965Ь, 1967; Акоев, Ильинский, 1972, 1973; Obara, Bennett, 1972; Акоев, Жадан и др., 1974; Акоев и др., 1975). Следует указать, что для рецепторов многих других специальных органов анодические стимулы являются возбуждающими, а катодические — тормозящими (Bennett, 1965; Hagiwara et al., 1965, и др.).
Ответ рецепторов на раздражение носит фазно-тонический характер. Даже при достаточно больших силах стимулов наблюдается явление полной адаптации рецепторов к действию постоянного тока в пределах нескольких минут (Акоев, Жадан и др., 1974) (см. также рис. 98). В момент выключения тока наблюдаются эффекты, противоположные тем, которые имеют место в момент его включения. Величина ответа при включении и выключении раздражения зависит от силы и длительности воздействия (Murray, 1962, 1965а; Акоев, Ильинский, 1972; Obara, Bennett, 1972). При кратких стимулах умеренной интенсивности эффект воздействия зависит от соотнесения момента раздражения с периоди-
чески возникающей фоновой активностью рецепторов (рис. 98). При сильных воздействиях (как и при разряде электрического органа у сильноэлектрических скатов) фоновая ритмика резко нарушается. Характерной особенностью ответов рецепторов ам-
Рис.
98. Действие электрического раздражения
на активность рецепторов ампул Лоренцини
ската Dasyatis pastinaca при естественных
ритмических дыхательных движениях (Л)
и при искусственном непрерывном
пропускании воды по жаберным щелям
(Б).

на
действие тока является большой скрытый
даже при значительных силах тока
составлял часто равнялся 20—40 мсек,
(Murray, 1965а;
пул Лоренцини период, который 10—15 мсек., а Акоев, Ильинский, 1972; Акоев, Жадан и др,, 1974). Работа рецепторов ампул Лоренцини неравно подверглась обследованию
с помощью внутриклеточных микроэлектродов (Obara, Bennett, 1972; см. также: Obara, Oomura, 1973).
В последнее время удалось показать (Броун и др., 1972; Андрианов и др., 1974; Акоев и др., 1975), что рецепторы ампул Лоренцини могут улавливать изменения магнитного поля (рис. 99). Постоянное магнитное поле было неэф-
IIIIIIIIIIUIIIIIIIIIIIIIIIIIIlillMiBmiiHiiiiiiiiiiiiiiHii
Рис.
99. Активность рецепторов ампул Лоренцини
ската Raja clavata при магнитной (1—4)
и электрической (5)
стимуляции.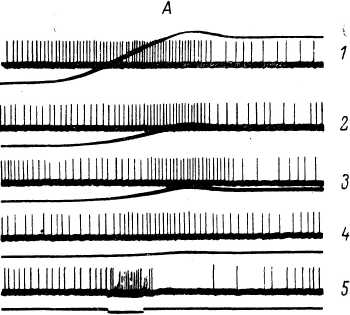
![]()
![]()
![]()

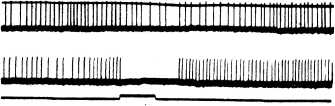

Плавная кривая — изменение интенсивности магнитного поля (северный полюс постоянного магнита на вентральной стороне тела; А — нарастание, Б — ослабление напряженности поля); скорость изменения поля: на! — 70, на 2 — 17, на з — 8 и на 4 — 3 гс/сек. На 5 — католическая (А) и анодическая (Б) электрическая стимуляция при интенсивностях 0.3 на/мм2. Отметка^времени — 500 мсек.; калибровка — 30 гс.
фективно. Величина реакции зависела от скорости изменения магнитного поля (порог равнялся 0.8 гс/сек.). Чувствительность рецепторов к изменению магнитного поля находилась в прямой зависимости от их чувствительности к электрическому току и от длины ампулярных каналов. Характер реакции рецепторов на магнитный стимул зависел как от направления вектора магнитной индукции, так и от расположения ампулярных каналов на теле животного. Авторы приходят к заключению о возможности восприятия рыбами магнитного поля Земли с помощью рецепторов ампул Лоренцини.
Температурная зависимость биоэлектрической активности механорецепторов
Изучение влияния температуры на те или иные возбудимые образования может дать важные сведения о процессах, лежащих в основе их нормальной деятельности (см., например: Самойлов, 1924). Это обстоятельство, а также тот факт, что температура, по-видимому, может в какой-то степени оцениваться организмом не только с помощью специфических терморецепторов, но и при участии механорецепторов (Минут-Сорохтина, 1972; Минут-Сорохтина, Глебова, 1974), послужили причиной весьма интенсивного изучения действия температурного фактора на различные механорецепторные приборы. Такого рода исследования были проведены в опытах с регистрацией как импульсной активности механорецепторов, так и РП. В последнем случае опыты проводились на одиночных рецепторах холоднокровных и теплокровных животных, таких как рецепторы растяжения у ракообразных (Florey, 1956; Burkhardt, 1959; Grampp, 1966), мышечные веретена лягушки (Matthews, 1931а; Ottoson, 1965b) и кошки (Eldred, Lindsley, Buchwald, 1960; Lippold et al., 1960b), тельца Пачини (Inman, Peruzzi, 1961; Ishiko, Loewenstein, 1961; Loewenstein, 1961b; Sato, 1961; Sato, Ozeki, 1963; Ильинский и др., 1965; Ильинский, 1966а).
Механорецепторы кожи
Кожные механорецепторы в большей степени, чем другие механочувствительные приборы, испытывают на себе действие температурного фактора. Поэтому они изучались в указанном плане достаточно подробно. Эти работы привели даже к возникновению представления о том, что действие температуры воспринимается организмом с помощью кожных механорецепторов. По мнению сторонников так называемой сосудистой теории терморецепции (Nafe, 1929, 1942; Kenshalo, Nafe, 1962; Минут- Сорохтина, 1965, 1967, 1968, 1972; Минут-Сорохтина, Глебова, 1974), холодовыми рецепторами кожи являются механорецепторы: собственно механорецепторы кожи, а также механорецепторы кожных и подкожных сосудов. В последнем случае стимуляция механорецепторов происходит вследствие сокращения гладких мышц сосудов, развивающегося в ответ на охлаждение. Тепловые рецепторы — это механорецепторы, которые при нагревании активируются в результате расслабления гладкой мускулатуры сосудов или, может быть, каким-то иным образом. Мы не будем вдаваться здесь в более детальное рассмотрение этой концепции (см. подробнее обзор: Минут-Сорохтина, 1972), которая в своей крайней форме фактически отрицает наличие специфических терморецепторов кожи и вряд ли может быть признана целиком (о морфологии и физиологии специфических терморецепторов см. обзоры: Hensel, 1973, 1974а). Вместе с тем следует согласиться, что температура может существенно изменять активность различных механорецепторных приборов (в первую очередь медленно адаптирующихся,) тем самым сигнализируя и с их помощью о своем действии на организм.
Некоторые нервные окончания кожи, так называемые механо- холодовые рецепторы, обладают достаточно большой чувствительностью как к механическим, так и к температурным стимулам. Такого рода рецепторы, связанные с миелиновыми афферентными волокнами, были обнаружены у птиц (Necker, 1974b) и млекопитающих:!^ кошки (Hensel, Zotterman, 1951а; Witt, Hensel, 1959; Iggo, 1968), обезьяны (Iggo, 1969) и человека (Hensel, Во-
man, 1960). Все эти рецепторы, по-видимому, являются на самом деле медленно адаптирующимися механорецепторами (у обезьяны пороговое механическое воздействие по параметру смещения составляло 5—20 мкм). Волокна этих рецепторов проводят импульсы значительно быстрее (скорость от 30 до 80 м/сек.), чем волокна специфических терморецепторов. Рисунок их разряда также отличался от ответа специфических холодовых рецепторов на действие охлаждения (сравн.: Hensel, Zotterman, 1951b; Hensel et al., 1960).
Действие температуры в наибольшей степени сказывается на ответах медленно адаптирующихся кожных механорецепторов II типа, которые; по-видимому, и изучались в известных работах (Witt, Hensel, 1959; Hunt, McIntyre, 1960a), но не были однозначно идентифицированы. Стационарная активность, вызванная в этих рецепторах действием на кожу постоянного давления, существенно изменялась под влиянием температуры: максимальная частота импульсации наблюдалась в пределах температур от 20 до 40°. При снижении температуры рецепторы отвечали увеличением частоты импульсации и, напротив, фоновая активность временно угнеталась при повышении температуры. Ответы рецепторов I типа в целом изменялись сходным образом, однако, в силу большей адаптационной способности, изменения импульсации были менее выражены. Максимум частоты находился в диапазоне 30—40°, а частота импульсов в стационарном разряде не превышала 20/сек. (Iggo, 1966а; Iggo, Muir, 1969).
При резком снижении температуры быстро адаптирующиеся механорецепторы могут отвечать несколькими импульсами. Это чаще наблюдается у наиболее чувствительных и наиболее медленно адаптирующихся, например у механорецепторов типа D из числа связанных с волосяными фолликулами (Iggo, 1966а). Интересно заметить, что у других чрезвычайно быстро адаптирующихся механорецепторов кожи — телец Пачини — даже резкое изменение температуры (от 60° до 0 со скоростью 38°/сек.) не вызывает появления импульсной активности (Loewenstein, 1961b).
В коже были обнаружены также рецепторы, связанные с не- миелинизированными С-волокнами, которые тоже отвечали как на действие умеренного механического раздражения, так и на влияние охлаждения (Douglas et al., 1960). Более того, некоторые С-волокна могли генерировать разряд при действии практически всех испытывавшихся видов стимулов: механических, термических (охлаждения и согревания), химических (Iriuchijima, Zotterman, 1960; Witt, 1963). Функция таких рецепторов остается неясной.
Изучение действия температуры проводилось также на механорецепторах кожи холоднокровных животных — рыб (Spath, 1967), земноводных (лягушка) (Catton, 1961) и рептилий (крокодил) (Necker, 1974а), В опытах на лягушках импульсная активность механорецепторов наблюдал ась^даже'пртГтемпературе 5°. Пороги стимуляции, необхрдимрй для появления ПД, нарастали над
при самых низких (5—12°), 1ак и при высоких температурах (28—35°). Быстрое охлаждение или нагревание вызывало появление нерегулярного механорецепторного разряда. У крокодила многие механорецепторы возбуждались при резком охлаждении. Величина ответа линейно возрастала при стимулирующем снижении температуры в диапазоне от 20 до 0°. При повышении температуры интенсивность ответа на механическое раздражение возрастала. Для многих механорецепторов максимум ответа на раздражение лежал в области 30°, а при дальнейшем нагревании частота ПД понижалась. Сходное изменение активности механорецепторов наблюдалось и у рыб.
В ряде работ проводилось изучение действия температуры на механорецепторы внутренних органов и сердечно-сосудистой системы. При этом большое внимание было уделено влиянию температурного фактора на афферентные нервные волокна (см. обзоры: Paintai, 1963а, 1972). Оказалось, что все мякотные волокна блокируются при одной и той же низкой температуре (около 7.5°) вне зависимости от диаметра (Paintal, 1965а; Franz, Iggo, 1968). Немиелинизированные волокна блокировались при более низкой температуре (порядка 5°), чем мякотные волокна (Paintal, 1967; Franz, Iggo, 1968; Sato et al., 1968; Fidone, Sato, 1969). Вместе с тем мякотные волокна, связанные с рецепторами, генерировавшими более высокочастотный разряд, изменяли свою активность в большей степени, чем волокна, частота импульсации которых была ниже. Например, снижение температуры сказывается в большей степени на «более высокочастотных» механорецепторах типа А предсердия, чем на рецепторах типа В, обладающих более низкой частотой импульсации (Paintal, 1953с, 1965b, 1966Ь).
Снижение частоты импульсации при понижении температуры отмечалось также и в опытах с каротидными (Diamond, 1955) и аортальными (Angell James, 1968, 1971с) барорецепторами. Температурный коэффициент (<?10) в случае каротидных механорецепторов равнялся примерно 2. Эта величина была такой же, как и для мышечных веретен лягушки (Ottoson, 1965b). Порог возбуждения сосудистых механорецепторов сдвигался при изменении температуры, но не слишком резко. По одним данным (Diamond, 1955), порог несколько увеличивался при снижении температуры, а по другим (Angell James, 1971с), он слегка уменьшался.
В экспериментах на тельцах Пачини, которые в этом плане изучались наиболее подробно (Inman, Peruzzi, 1961; Ishiko, Loewenstein, 1961; Ильинский и др., 1965; Ильинский, 1966а) было показано, что при понижении температуры амплитуда Pli закономерно снижается. При этом уменьшалась не только величина РП, генерируемого при данной одной и той же силе раздражения, но и величина максимального РП. Вся кривая зависимости амплитуды РП от силы раздражения по мере снижения температуры приближалась к оси силы стимуляции, деформируясь и смещаясь в область меньших амплитуд РП и больших сил стимуляции (рис. 100). При температуре ниже 13—16° кривая переставала выходить на уровень насыщения, и анализ данных в этой температурной области был затруднительным. Выпадали из общей картины также результаты, полученные при температуре выше 41°.
Как следует из рис. 100, величина 7?тах (см. стр. 274) (можно думать и 7?0) меняется при температурном воздействии. Очевидно, что изменение кривой зависимости амплитуды РП от силы раздражения под влиянием температурного фактора носит совершенно иной характер, чем в случае действия постоянного электрического тока (сравн. рис. 96). Наблюдаемые изменения позволяют говорить о чувствительности процессов трансформации, протекающих в данном механорецепторе при механическом раздражении, к действию температуры.
Энергия активации, рассчитанная по изменениям амплитуды максимального РП, составляет величину порядка 3—8 ккал./моль. Несколько большее значение энергии активации (9—И ккал./моль) оказалось характерным для изменений силы раздражения (•Ятах)> при которой регистрировался максимальный РП (Ильинский и др., 1965). Этот параметр, как отмечалось выше, в известной мере связан с процессами трансформации в рецепторной мембране. Еще больше (13—16 ккал./моль) была энергия активации, определяемая по изменению параметра Унар — скорости нарастания РП (Inman, Peruzzi, 1961; Ishiko, Loewenstein, 1961; Ильинский и др., 1965). Скорость нарастания РП, связанная с величиной генераторного тока, также характеризует некоторые стороны трансформационного процесса. Таким образом, можно говорить о том, что трансформационные явления в тельцах Пачини связаны с процессами, обладающими достаточно большой энергией активации. Вместе с тем они не обязательно должны быть обусловлены какими-либо химическими преобразованиями, а могут быть удовлетворительно описаны и в рамках чисто физических явлений, развертывающихся на рецепторной мембране. Достаточно указать, что даже такой относительно простой процесс, как диффузия воды через монослой жирной кислоты, характеризуется энергией активации порядка 14.5 ккал./моль (Archer, LaMer, 1955).
По мере снижения температуры длительность фазы нарастания РП увеличивалась, а фазы спадения РП уменьшалась (рис. 100). Однако эти изменения были сравнительно небольшими, т. е. температурная зависимость указанных параметров оказалась низкой: коэффициент ()10 для длительности фазы нарастания РП равнялся, например, 1.58, а фазы спадения — 1.10, т. е. был много меньше, чем для Рнар>, <2ю которого доходило до 2.6 (Inman, Peruzzi, 1961). При этом важно отметить, что изменения длительности восходящей и нисходящей фаз РП были сходными, но противоположно направленными. В итоге общая продолжи-
*
34.7
Рис.
100. Зависимость активности тельца Пачини
от температуры (А
—
по: Ishiko, Loewenstein, 1961а; Б
— по:
Ильинский и др., 1965).
А
—
влияние температуры (цифры
над кривыми)
на РП (а)
и ПД (б) (схематизировано). Отметка
времени — 1 мсек.; кали бровка: на а —
25, на б — 100 мкв. Б
—
зависимость амплитуды РП от силы
раздражения при различной температуре
(ука зана на кривых). По
оси абсцисс —
сила раздражения (R,
усл. ед.); по
оси ординат —
амплитуда РП (V,
усл. ед.).
19
Физиология механорецепторов
тельность РП не менялась или лишь слегка увеличивалась. Характер температурного изменения нисходящей фазы РП позволяет считать, что она обусловлена пассивными электрическими свойствами мембран. Это обстоятельство, а также отсутствие зависимости общей продолжительности РП от температуры позволяют утверждать, что развитие РП в тельцах Пачини едва ли связано с кинетикой какой-либо химической медиаторной реакции.
Некоторые авторы считают, что при механическом воздействии на рецепторный прибор из окружающих нервных окончаний структур внутренней колбы рецептора может выделяться медиатор, действующий на нервное окончание (подробнее эта медиаторная гипотеза будет рассмотрена в главе 8). В таком случае следовало бы ожидать более существенное замедление временного течения РП по мере охлаждения, как это и имеет место в случае мионевральной передачи (Eccles et al., 1941). Таким образом, эксперименты с температурным воздействием делают более вероятным предположение, что развитие РП обусловлено чисто физическими процессами, связанными с диффузией ионов через растянутую мембрану, хотя и не окончательно отвергают возможность, что появление РП обусловлено также и какими-то химическими превращениями на мембране немедиаторного происхождения, итогом которых является изменение ее проницаемости.
Сравнительное изучение изменений on- и off-РП под влиянием температуры показало (Ильинский и др., 1965), что оба ответа менялись при изменениях темпераауры одинаково. Это хорошо согласуется с исследованиями, показавшими, что ответ на выключение в тельцах Пачини обусловлен механическими свойствами вспомогательных структур рецептора, а не какими-либо особенностями рецепторной мембраны (Ильинский, 1966в; Loewenstein, Skalak, 1966).
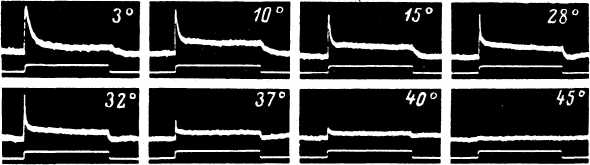
Действие ъемпературы на ПД телец Пачини было отличным от ее влияния на РП этих рецепторов, но сходно с влиянием температуры на ПД других мякотных нервных волокон млекопитающих (см.: Ishiko, Loewenstein, 1961). Снижение температуры приводило к возрастанию длительности ПД. При этом длительность восходящей и нисходящей фаз ПД изменялась одинаково. Скорость нарастания и скорость спадения монофазного ПД при этом уменьшалась (рис. 100). При охлаждении могла весьма возрастать и следовая гиперполяризация. Латентное время ПД сильно удлинялось при понижении температуры, а порог возникновения спайка возрастал. В основе этого лежало как изменение возбудимости регенеративной системы генерации, так и изменения РП (его амплитуды и скорости нарастания). При охлаждении абсолютная величина флуктуаций порога возникновения ПД возрастала, а относительная (рассчитанная в единицах порога для данной температуры) — значительно уменьшалась. Аналогичным образом в ходе охлаждения менялись пороги возбуждения нервного волокна телец Пачини при прямом электрическом раздражении (Ильинский и др., 1965). Изменение параметров РП и (или) уменьшение возбудимости регенеративной системы при охлаждении обнаруживается также в виде следующего феномена. Если при достаточно высокой температуре в ответ на одиночный краткий толчок рецептор генерировал два спайка, возникавших из одного РП, то после понижения температуры возникал лишь один ПД (Inman, Peruzzi, 1961).
Как показали эксперименты с вибрационным воздействием (Sato, 1961), порогово-частотная кривая при охлаждении смещается в область более низких частот (при 37° минимальный амплитудный порог к вибрационным воздействиям наблюдается в области 300 гц, а при 24° — около 150—200 гц) (рис. 41). Амплитуда ПД при начальном охлаждении несколько возрастала: максимум, по данным разных авторов, наблюдался либо около 23° (Ishiko, Loewenstein, 1961), либо около 35° (Inman, Peruzzi, 1961), либо в бодее широкой зоне — 30—35° (Ильинский и др., 1965). При дальнейшем охлаждении ПД уменьшались особенно сильно при t < 20—25°. При температурах 18—12° ПД исчезал. Как явствует из этих данных, ПД телец Пачини переставал возникать при температурах, более высоких, чем те, при которых возникают блоки проведения в миелинизированных афферентных нервных волокнах. В последнем случае выключение проведения имело место в районе 7.2—7.6° (Paintai, 1965а; Franz, Iggo, 1968). Более высокая чувствительность механорецепторов к охлаждению по сравнению с нервными проводниками несомненно связана с появлением при охлаждении трудностей для возникновения ПД в регенеративной системе генерации, обусловленных изменениями ее свойсгв, а также изменениями регенеративной системы генерации (снижением амплитуды и скорости нарастания РП). Следует напомнить, что система генерации ПД в тельцах Пачини обладает известными особенностями, что находи! отражение в ее очень быстрой аккомодационной способности (J. А. В. Gray, Matthews, 1951b). О возможном значении аккомодационных свойств регенеративной системы генерации в эффектах действия температуры на тельца Пачини говорит, например, тот отмеченный ранее факт, что с помощью лишь температурных изменений невозможно вызвать возбуждение данных рецепторов (Loewenstein, 1961b), в то время как другие, более медленно адаптирующиеся тканевые механорецепторы, в той или иной степени могут возбуждаться при действии этого фактора среды. В заключение следует отметить, что, как показали специальные эксперименты (Ishiko, Loewenstein, 1961), действие температуры было одинаковым как на активные, так и на декапсулированные тельца Пачини, т. е. все наблюдавшиеся явления никак не были связаны со вспомогательными структурами этих рецепторов.
Мышечные рецепторы
В работе, проведенной на мышечных веретенах кошки (Lippold et al., 1960b), было установлено, что при температуре ниже 32° нервные окончания дают регулярные неадаптирующиеся афферентные разряды даже в состоянии полного расслабления мышц. Частота этих разрядов зависела от величины температуры, а не от скорости ее изменения. Наибольшая активность отмечалась примерно при охлаждении до 28°. Затем активность снижалась и полностью прекращалась в области 15°. Возникновение импульсной активности, по мнению авторов, не было обусловлено локальным сокращением мышечных структур, а явилось результатом изменений свойств самих рецепторных элементов. Авторы считают, что такого рода ответ мышечных веретен является результатом специфической «холодовой» деполяризации рецепторной мембраны. Действие различных других деполяризующих факторов, таких как натяжение рецепторов, удаление ионов кальция, увеличение концентрации ионов калия, приложение катода постоянного тока суммировалось с холодовой деполяризацией и увеличивало эффект холода. Напротив, гиперполяризующие воздействия уменьшали или блокировали температурные влияния.
Несколько отличные данные были получены другой группой исследователей (Eldred, Lindsley, Buchwald, 1960), которые обнаружили при охлаждении угнетение активности растянутых мышечных веретен. Причина такого расхождения в результатах остается неразъясненной. По-видимому, все-таки следовало бы более тщательно исследовать действие холода на интрафузаль- ную мускулатуру веретен, с тем чтобы выяснить, не вызывается ли возбуждение рецепторов сокращением этих элементов в ответ на охлаждение, особенно учитывая данные (Buchtal, Zahn, 1951) о возможности спонтанных сокращений отдельных интрафузальных волокон лягушки при температуре 20°. У теплокровных животных исследовалось также влияние температуры на различные характеристики импульсной активности веретен (см., например: Poppele, Bowman, 1970).
У холоднокровных животных изучалось влияние температуры на рецепторы растяжения у рака и на мышечные веретена у лягушки. Полученные данные оказались весьма сходными между собой и, естественно, несколько отличались от результатов экспериментов на теплокровных животных. В опытах на мышечном веретене лягушки было установлено (Matthews, 1931а), что по мере увеличения температуры (в диапазоне 10—26°) частота разряда на стимул одной и той же величины возрастает, рефрактерный период после ПД уменьшается, в то же время общая длительность импульсного ответа уменьшается, т. е. скорость адаптации растет. Тем не менее общее число импульсов в ответе увеличивается. Принципиально сходные, но более детальные данные по влиянию
температуры на импульсную активность мышечного веретена лягушки были получены в экспериментах (Ottoson, 1965b), в которых исследовались не только особенности импульсной активности, но и свойства РП (рис. 101). Оказалось, что число импульсов при данной силе растяжения возрастало при увеличении тем-
А
Б В
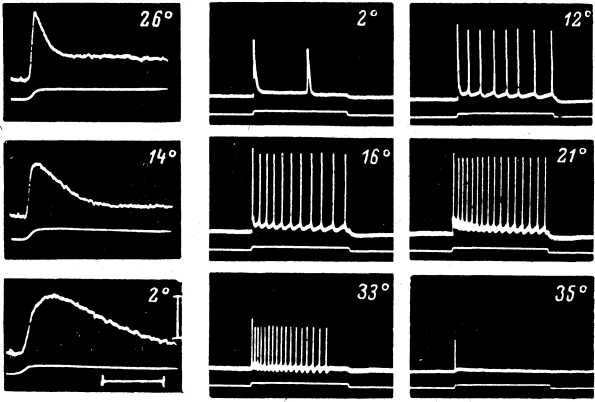
Рис. 101. Ответы изолированного мышечного веретена при различной температуре (по: Ottoson, 1965b).
А — изменение РП при нагревании рецептора; Б — изменение динамического компонента РП при охлаждении (на А и Б ПД блокированы лигнокаином); В — импульсная активность рецептора. Сверху вниз: биоэлектрическая активность; механическое раздражение. Длительность раздражения на А и Б — 300 мсек. На Б: отметка времени — 20 мсек.; калибровка — 0.5 мв.
пературы от 3 до 30°, а затем (выше 30°) быстро падало. Реакция исчезала полностью при температуре выше 35°. Амплитуда ПД при нагревании снижалась. Вначале (от 2 до 22°) она изменялась незначительно, а затем (от 22 до 36°) весьма резко.
Действие температуры на динамический и статический компоненты РП несколько разнилось (рис. 101, А—Б). От 3 до 25° динамический компонент реакции оставался почти неизменным ((2io близко к единице), в то время как статическая составляющая несколько падала (<2ю составляет, примерно, 1.2). Выше 25° амплитуда обоих компонентов прогрессивно снижалась, особенно резко после 30° (<2ю равнялось, соответственно, 1.80 и 2.45). При температуре около 40° РП полностью исчезал. Длительные нарастания и спадения динамической фазы ответа экспоненциально уменьшались при увеличении температуры от 3 до 35°, а время разрушения статической фазы после снятия раздражения возрастало при этом почти линейно. Ни быстрое охлаждение, ни быстрое нагревание сами не вызывали появления импульсной активности. Следует отметить, что изменение параметра РП происходило весьма неравномерно, что нашло свое отражении в виде разных величин коэффициента (2ю Для одного и того же параметра в разных диапазонах температуры.
В обстоятельной работе (Burkhardt, 1959) было установлено, что по мере увеличения температуры в диапазоне от 3 до 33° увеличивается частота импульсации тонического рецептора растяжения у рака. При этом эффект температуры был тем больше, чем значительнее ,было исходное натяжение рецептора и соответственно, его исходная спайковая активность. В момент повышения температуры часто1а импульсации кратковременно, но резко падала, а затем постепенно возрастала до значения, превышающего исходный уровень. При снижении температуры наблюдалась обратная картина. При температуре свыше 30° спайки начинали группироваться в пачки, частота разрядов в которых, а также продолжительность пачки импульсов были тем больше, чем выше была температура. При температуре выше 33° активность рецептора внезапно исчезала. При увеличении температуры наблюдалось сильное ускорение процесса адаптации рецептора к адекватному раздражению. Следует отметить, что какого- либо максимума активности при определенной температуре, которая могла бы рассматриваться как оптимальная для рецепторов растяжения, не было, что существенно отличает их от терморецепторных приборов (сравн., однако: Florey, 1956).
С помощью внутриклеточного отведения потенциалов было установлено (Burkhardt, 1959), что повышение температуры вызывает уменьшение РП. Однако наряду с этим эффектом наблюдалось снижение порога возникновения ПД. Суммарный эффект влйяния повышения температуры на ритмическую активность зависел от взаимодействия этих противоположно направленных эффектов влияний: в первый момент сказывалось действие температуры на РП, а затем начинал преобладать эффект изменения порога генерации ПД. При снижении температуры направленность реакции была противоположной. Возможно, что в определенных пределах изменения РП и порога возникновения ПД могут взаимно компенсироваться и тем самым нивелировать температурные изменения, что может иметь определенное значение для пойкило- термных животных,
Имеются данные (Grampp, 1966), согласно которым увеличение температуры приводит к возрастанию максимальной скорости деполяризации и реполяризации мембраны и к снижению амплитуды ПД. Температурные изменения скорос1и деполяризации и реполяризации сомы и дендритов клетки были выражены меньше, чем в аксоне, а амплитуда ПД аксона падала медленнее, чем в сомо-дендритной области. Это, по мнению автора, говорит о неодинаковом распределении по поверхности мембраны рецепторного нейрона регенеративных элементов.
Волосковые рецепторы органов чувств
Исследование свойств РП волосковых механорецепторов органов чувств у позвоночных, как отмечалось выше, только начинается. И влияния температурного фактора на РП в Э1их сенсорных приборах пока не изучалось.
В ряде работ испытывалось температурное воздействие на активность специализированных органов боковой линии — ампул Лоренцини, рецепторы которых, как отмечалось, обладают значительной механической чувствительностью (Hensel, 1955, 1974b). Выявилась очень высокая чувствительность рецепторов этих образований к такого рода влияниям, что первоначально даже позволило говорить о терморецепторной функции данных сенсорных приборов.
♦ ♦ ♦
Изложенные материалы показывают, что температурный фактор действует на различные звенья процессов, протекающих в механорецепторах. В этой связи многие данные, полученные в такого рода исследованиях, не могут быть истолкованы однозначно. В частности это касается такого важного вопроса, как природа процесса 1рансформации в механорецепторах. Результаты, полученные на тельцах Пачини с использованием температурного воздействия, с одной стороны, позволяют утверждать, что медиаторный механизм не имеет места в цепи явлений, связанных с появлением РП в ответ на механическое раздражение этих механорецепторов. С другой стороны, имеющиеся данные не дают возможности однозначно решить вопрос о том, связаны ли трансформационные явления в этих (и в других) тканевых механорецепторах с преимущественно химическими или физическими явлениями.
Исследование механорецепторов с помощью других физических факторов
Ультразвуковое воздействие. В последние годы было проведено несколько работ, посвященных изучению действия ультразвука (УЗ) на механорецепторы (Макаров и др., 1964; Гаврилов и др., 1972, 1973, 1974).
В работе Макарова и др. (1964) изучалось влияние УЗ на мышечное веретено лягушки. При длительных воздействиях УЗ спонтанная импульсация рецептора могла либо уменьшаться (при больших интенсивностях), либо увеличиваться (при малых интенсивностях). Воздействие УЗ в виде короткого стимула могло вызвать появление ПД покоящегося рецептора. Механизм действия УЗ не исследовался.
Основной целью работы Гаврилова с соавторами было выяснение принципиальной возможности возбуждения с помощью фокусированного УЗ различных периферических нервных структур, расположенных как на поверхности тела, так и в его глубине. Хотя основная часть материалов этих исследований была получена в экспериментах на людях (оценка эффектов проводилась на основе субъективных ощущений), тем не менее некоторые из установленных фактов имеют прямое отношение к рассматриваемым нами проблемам. Оказалось, что фокусированный УЗ, действовавший в течение коротких интервалов времени при достаточной мощности, способен вызвать у человека все виды ощущений (в том числе и тактильные), которые могут наблюдаться при естественных воздействиях на кожу и ткани конечностей. Анализ действия УЗ привел авторов к заключению, что из всех теоретически возможных факторов, которые сопутствуют распространению УЗ в среде, важнейшими являются смещение среды в фокальной области и изменения температуры, обусловленные поглощением УЗ энергии в тканях.
Фактором, прежде всего ответственным за возникновение различных ощущений, является амплитуда колебаний структур тканей в фокусе УЗ. Оказалось, что одни и те же пороговые ощущения (в том числе и тактильные) при воздействии фокусированным УЗ различной частоты соответствуют приблизительно одинаковым амплитудам смещения. Остальные параметры (радиационное давление, кавитация и др.) при этом сильно меняются, иногда даже на несколько порядков. Не вдаваясь в более детальное рассмотрение указанных работ, следует обратить внимание на тот важный факт, что именно амплитуда смещений, а не величина давления является существенной для возбуждения нервных структур. Этот факт полностью согласуется с другими имеющимися в литературе данными (см. стр. 455), относящимися к механизму действия механического воздействия на различные нервные структуры (в том числе и на механорецепторы).
При действии фокусированного УЗ на изолированные из брыжейки кишечника кошки одиночные тельца Пачини можно было зарегистрировать возникновение ПД. Порог возникновения спайка при частоте 480 кгц в величинах амплитуды колебательного смещения равнялся 0.03—0.05 мкм (Гаврилов и др., 1974), что хорошо согласуется с данными о порогах стимуляции телец Пачини, полученными иными методами (J. А. В. Gray, 1959b; Loewenstein, 1965; Ильинский, 1966а). Как уже отмечалось, порог тактильного ощущения, по-видимому, определяется порогом возбуждения наиболее чувствительных механорецепторов кожи — телец Пачини (Lindblom, Lund, 1966). Сопоставление пороговых интенсивностей УЗ, вызывавших тактильные ощущения у человека, с пороговыми значениями интенсивностей фокусированного УЗ, вызывавшими появление ПД в изолированных тельцах Пачини кошки, показало их хорошее соответствие (Гаврилов и др., 1974).
Фокусированный УЗ мог вызывать возбуждение и волосковых механорецепторов органа слуха. Пороговыми при частоте 480 гц были интенсивности порядка 0.01 —0.1 вт/см2 (соответственно 0.004—0.01 мкм по амплитуде колебательного смещения).
Действие ионизирующей радиации. В литературе имеется ряд работ, посвященных анализу действия этого фактора на различные элементы нервной системы, в том числе и на механорецепторы (органа слуха, вестибулярного аппарата и др.) (см., например: Лебединский, Нахильницкая, 1960; Ливанов, 1962; Haley, Snider, 1962, 1964; Cleave, 1963, и др., —а также труды специального симпозиума «Effect of ionising radiation on the nervous system», 1962). Однако сенсорные элементы в целом исследовались в этом направлении меньше (Гейнисман, Жирмунская, 1952; Африка- нова, 1956; Ливанов, Делицына, 1956; Делицына, 1957, 1958, 1960; Ливанов, 1957; Лебедев, 1958, и др.) и, как правило, лишь с оценкой суммарной активности большого количества элементов, что, естественно, затрудняет трактовку полученных результатов. На одиночных же механорецепторах проводились лишь единичные работы (Ильинский, Комаров, 1963; Talbott et al., 1969).
В экспериментах на одиночных тельцах Пачини было показано (Ильинский, Комаров, 1963), что облучение пучком [3-частиц вызывало повышение возбудимости рецептора к действию механического раздражения. Понижение порога генерации ПД и уменьшение скрытого периода ответа происходили плавно в ходе облучения по Мере увеличения дозы (максимальная доза достигала 930 рад за 70 мин. облучения). После облучения возбудимость в первые 10 мин. сохранялась повышенной. Под влиянием облучения возникновение возбуждения (в отсутствие механического раздражения) в механорецепторах не наблюдалось. Было показано, что в применявшихся дозах облучение не изменяло свойства афферентных нервных проводников.
Авторы делают заключение о преимущественном влиянии радиации на нерегенеративную систему генерации в механорецепторе. С указанным выводом хорошо согласуются данные, полученные при изучении других нерегенеративных структур — постсинаптических мембран мотонейрона кошки (Sato, Austin, 1964). В упоминаемых опытах было показано, что под влиянием ионизирующей радиации (доза 600—15000 рад) наблюдалось увеличение натриевой проводимости, ВПСП возрастал. Деятельность натриевого насоса при этом не менялась, и потенциал покоя клетки был постоянным. Амплитуда ТПСП также не менялась, т. е. можно было думать, что проницаемость для ионов, обусловливающих развитие ТПСП, оставалась прежней.
В работе на других механорецепторах — рецепторах движения в тонком кишечнике кошек (Talbott et al., 1969) —были получены данные, принципиально сходные с результатами экспериментов на тельцах Пачини.
Действие ультрафиолетового облучения. В литературе имеются данные по влиянию ультрафиолетового света на рецептор растяжения у ракообразных (Giese, Furshpan, 1954). Под влиянием такого рода воздействия фоновая частота импульсации МРО первоначально возрастала, а затем снижалась и полностью пропадала. Механизм действия этого вида энергии остается невыясненным.
Чувствительность механорецепторов к действию химических веществ
Изучение действия различных веществ на механо- рецепторы проводилось в очень большом числе работ (наиболее полные обзоры: Aviado, Schmidt, 1955; Heymans, 1955; J. А. В. Gray, Diamond, 1957; Heymans, Neil, 1958; J. A. B. Gray, 1959b; Черниговский, 1960; Edwards, 1960; Douglas, Ritchie, 1962; Paintal, 1963a, 1964, 1971, 1972; С. M. Smith, 1963, 1967; Matthews, 1964, 1972; Zipf, 1966; Ильинский, 1967; Fex, 1973, и др.). Исследователи пытались с помощью фармакологических средств понять некоторые особенности звеньев механизма активации механорецепторов и свойства их волокон, что представляет для целей данной работы наибольший интерес. В некоторых работах химические агенты использовались для целей идентификации механорецепторов (например, подразделения мышечных веретен и сухожильных органов Гольджи —см.: Granit et al., 1953; G. M. Smith, 1963). Наконец, в ряде работ авторов больше интересовал чисто фармакологический аспект исследуемых вопросов. Следует заметить, что фармакологический подход широко используется при изучении механизмов эфферентной регуляции деятельности рецепторов (мышечных веретен, волосковых механорецепторов органов чувств позвоночных, рецепторов растяжения у ракообразных). Эта группа работ рассматривалась в главе 5. В некоторых работах использование ряда химических воздействий осуществлялось с целью изучения метаболических процессов, протекающих в нервных клетках. Удобным объектом для изучения изолированных нейронов являются рецопторы растяжения у ракообразных благодаря размерам сомы клетки и своей доступности (см., например: Rossini et al., 1966). Отдельные исследования такого характера, имеющие отношение к рассматриваемым в данной работе проблемам, будут затронуты в главе 8.
В подавляющем большинстве случаев из-за методических трудностей при изучении действия химических агентов рассматривалась лишь импульсная активность рецепторных приборов. Рецепторные же потенциалы и их изменение под влиянием различных веществ исследовались, как и во многих других случаях, на традиционных объектах: тельцах Пачини, мышечных веретенах, рецепторах растяжения у ракообразных.
Согласно существующим в настоящее время воззрениям (Pain- tal, 1964, 1971, 1972), основной точкой приложения действия различных веществ на механорецепторы (прежде всего первичные) является их регенеративная система генерации. Механочувстви- тельный субстрат рецепторов, по-видимому, в значительно меньшей степени изменяет свои свойства под влиянием этих раздражителей (см. рис. 95). (Конечно общая картина реакции может быть весьма осложнена при действии химического фактора на сосуды и окружающие ткани).
Как уже отмечалось, в механорецепторах поверхность, генерирующая рецепторный потенциал, и регенеративная система генерации (точнее, область, являющаяся наиболее благоприятной для возникновения ПД), как правило, пространственно разделены. Очевидно, что в такой ситуации исключительно большое значение имеют диффузионные барьеры рецептора, которые могут быть весьма различными в разных его частях. Это хорошо видно на примере телец Пачини. В данном случае немиелинизированная часть нервного окончания заключена в многослойную капсулу, наружные оболочки которой очень плохо проницаемы для жидкостей (J. А. В. Gray, Sato, 1955).2 Понятно, что при такой структуре химический раздражитель, действующий извне (например, при диффузии по межклеточному пространству в коже, стенках внутренних органов или в стенках сосудов), сможет действовать на расположенные в этих местах рецепторы исключительно посредством проникновения в область нервного волокна, точнее в область 2—3-го перехватов Ранвье, которые находятся вне капсулы рецептора (Quilliam, Sato, 1955; Ильинский, 1966г; Волкова, 1972а). Лишь при декапсулировании тельца химический агент может оказывать действие и на более глубоко расположенные структуры рецептора, что достигается в условиях эксперимента. В случае же подведения химических веществ к тельцу Пачини через сосудистое русло преимущественной точ-. кой приложения химических агентов оказывается область первого перехвата Ранвье, или самое начало безмякотного нервного окончания, т. е. то место, где чаще всего ветвятся подходящие к рецептору капилляры (Diamond et al., 1958b; Волкова, Черепнов, 1971; см. также: Pallie et al., 1970). Различия в диффузионных барьерах у разных структур рецептора имеют место и в других
2 Благодаря этому обстоятельству жидкость, находящаяся внутри капсулы, не вытекает наружу и тельце не спадается. Удаление наружных оболочек приводит к быстрому выделению жидкого содержимого капсулы рецептора, что позволяет, в частности, изучать его состав (см. главу 7, стр. 374). сенсорных механочувствительных приборах (Paintai, 1964, 1971).XIV По-видимому, отсутствие капилляров в непосредственной близости к рецептирующим элементам, например около наружных волосковых клеток кортиева органа (см. стр. 342) или около немиели- низированных окончаний нервных волокон в тельцах Пачини и в мышечных веретенах (см. главу 1), должно препятствовать быстрому проникновению к рецепторам из крови различных веществ, изменяющих их возбудимость или даже способных вызвать их возбуждение «неадекватным» способом. Следует заметить, что вопрос о том, почему у рецепторов — структур, обладающих таким интенсивным обменом (см. главу 8), — капилляры не подходят непосредственно к основному рецептирующему элементу системы,XV является весьма интересным, но практически совсем не разработанным.
Здесь не представляется возможным рассматривать влияние всех химических агентов на механорецепторы. Целесообразно ограничиться изложением лишь наиболее интересных данных, позволяющих понять некоторые существенные особенности звеньев механизма возбуждения этих сенсорных приборов. О влиянии некоторых веществ (катехоламинов, гамма-аминомасляной кислоты и стрихнина) уже упоминалось выше (см. соответственно: стр. 213, 255, 257).
Влияние ацетилхолина. Действие экзогенного ацетилхолина и различных холиномиметиков представляет специальный интерес в связи с гипотезой о принадлежности тканевых механорецеп- торов^к вторичночувствующим сенсорным приборам (подробнее об этой медиаторной гипотезе см. главу 8). Одним Гиз основных критериев, которыми эту гипотезу обосновывают ее сторонники, является возбуждающее или облегчающее действие экзогенного
ацетилхолина, которое отмечалось авторами при подведении этого агента к механорецепторам (например: Brown, Gray, 1948; Jarrett, 1956). Согласно медиаторной гипотезе, ацетилхолин является наиболее вероятным кандидатом в медиаторы, который может выделяться из клеточных элементов, окружающих нервные окончания тканевых (прежде всего, инкапсулированных) механорецепторов.
Влияние ацетилхолина и других веществ со сходным действием на различные механорецепторы 5 изучалось очень многими авторами, например в опытах на рецепторах кожи (Brown, Gray, 1948; Habgood, 1950; Douglas, Gray, 1953; Jarrett, 1956; Douglas, Ritchie, 1959; Fjallbrant, Iggo, 1961; K. R. Smith, Creech, 1967), мышечных веретенах (Гершуни, Нарикашвили 1942; Hunt, 1952; Granit et al., 1953; Henatsch, Schulte, 1958; Ottoson, 1961; С. M. Smith, Eldred, 1961; Verhey, Voorhoeve, 1963; Burgess, Clark, 1969b), интероцепторах (Brown, Gray, 1948; Landgren et al., 1953, 1954; Paintai, 1954a, 1954b; Widdicombe, 1954b; Diamond, 1955; Джелиев, Черниговский, 1959; Witzleb, 1959; Акоев и др., 19746), волосковых рецепторах органов чувств позвоночных (Russell, 1968, 1971b; Bobbin, Konishi, 1971), механорецепторах беспозвоночных (Wiersma et al., 1953; Florey, 1954a, 1957a; Ташмухамедов, 1961; Washizu, 1965; McLennan, York, 1966). Обычно в такого рода экспериментах регистрировалась лишь импульсная активность в афферентных волокнах, а иногда о реакции механорецепторов судили не по электрофизиологическому показателю, а по изменению, например, кровяного давления. Естественно, что точка приложения раздражителя в таких опытах могла быть определена лишь ориентироводно на основании ряда косвенных данных. После подведения к механорецепторам ацетилхолина и других сходных веществ часто наблюдается появление импульсации или увеличение фоновой активности. Повышается также чувствительность нервных окончаний к адекватному раздражению. При увеличении дозы введенного ацетилхолина до 1 -10“4 г некоторые механорецепторы (кожи, языка) вслед за начальной фазой повышения активности впадают в состояние угнетения и не отвечают ни на механическое, ни на повторное химическое раздражение (Landgren et al., 1953; Jarrett, 1956). Состояние угнетения постепенно проходит. Сходное действие ацетилхолина наблюдалось и в опытах на терморецепторах кожи и на вкусовых рецепторах языка (Bing, Skouby, 1950; Dodt et al., 1953; Landgren et al., 1953). В других механорецепто-
Несомненно также, что эффекты химических веществ типа ацетилхолина, никотина и др. на различные неидентифицированные рецепторы, среди которых, как можно полагать, были и механорецепторы, воспроизводились во многих экспериментах В. Н. Черниговского и его сотрудников, когда применялась необходимая для доказательства рефлекторных механизмов методика перфузии различных органов и структур, изолированных в гуморальном отношении от организма (см.: Черниговский, 1943в, 1960). pax (каротидного синуса, кишечника) состояние длительного угнетения после активации не наблюдается.
В отдельных случаях ацетилхолин может действовать на механорецепторы опосредованно через контрактильные элементы окружающих тканей и вспомогательного аппарата. Так, по-видимому, он действует на мышечные веретена (Hunt, 1952; Ottoson, 1961, и др.). На сами же нервные окончания в этом случае он влияет в меньшей степени или же не влияет совсем. Он может действовать, по-видимому, и на постсинаптические системы эфферентных синапсов в волосковых механорецепторах органов чувств (Russell, 1971b). В опытах на рецепторах растяжения у рака (Wiersma et al., 1953; Florey, 1954a, 1957a; Washizu, 1965), барорецепторах каротидного синуса у кошек (Diamond, 1955), механорецепторах кожи (Jarrett, 1956) и внутренних органов у кошек (Джелиев, Черниговский, 1959) отмечалось прямое возбуждающее действие ацетилхолина на механорецептивные сенсорные приборы.
Экспериментальная проверка идеи о возможном участии ацетилхолина в актах возбуждения механорецепторов проводилась рядом авторов. Полученные данные во всех случаях привели исследователей к заключению, что ацетилхолин не причастен к нормальным этапам трансформационных процессов и генерации РП в первичных механорецепторах (обзоры: J. А. В. Gray, 1959а; Ильинский, 1967). Это доказывается тем, что после введения постсинаптических блокаторов холинергической передачи, таких как тубокурарин, гексометоний и никотин, примененный в больших дозах, возбуждающее действие ацетилхолина на механорецепторы кожи и барорецепторы каротидного синуса отсутствовало, в то время как ответ на механический стимул не претерпевал никаких изменений (Brown, Gray, 1948; Douglas, Gray, 1953; Diamond, 1955; Jarrett, 1956). Атропин также не оказывал никакого влияния на эффекты адекватной стимуляции (Jarrett, 1956). Возбуждающее действие ацетилхолина или снижение порогов механического раздражения при подведении ацетилхолина авторы связывают с деполяризующим действием этого вещества на нервные волокна (регенеративную систему генерации). На основании анализа большого количества фактов, относящихся к разным механорецепторам, к такому же заключению пришел и Пайнтал (Paintai, 1964). Он же считает, что никакие медиаторы не участвуют в актах возбуждения тканевых механорецепторов при действии на них адекватных стимулов (Paintai, 1964, 1972).
, Недавно подробное исследование действия ацетилхолина и блокаторов холинергической передачи было осуществлено в опытах на декапсулированных тельцах Пачини при регистрации не только ПД, но и РП (Акоев и др., 1.9746). В этих экспериментах было установлено, что концентрации ацетилхолина 1«10“6 г/мл и ниже не оказывают на тельце Пачини заметного влияния. Даже более концентрированные растворы ацетилхолина (1 -10~3 г/мл) не вызывали появления импульсной активности в рецепторе, хотя и уменьшали пороговые величины механических стимулов. (В этих случаях под влиянием ацетилхолина снижался критический уровень РП, необходимый для генерации ПД). Небольшие концентрации ацетилхолина не влияли на РП, а большие (1*10~3 г/мл и выше) уменьшали амплитуду ответа. Атропин, гексаметоний и d-тубокурарин даже при длительном действии не блокировали импульсную активность рецептора, вызванную механическим раздражением. Более того, d-тубокурарин снижал критическую величину деполяризации и увеличивал количество ПД в ответе, но уменьшал при этом и амплитуду РП. Возможно, что это было обусловлено его собственным деполяризующим действием. Ионы магния, которые, как известно, уже в небольших дозах блокируют выделение ацетилхолина из пресинаптических окончаний (Castillo, Engbaek, 1954), даже при больших концентрациях (до 20 ммолей/л) не угнетали ответы телец Пачини при адекватной стимуляции. Все эти данные однозначно свидетельствуют против какого бы то ни было участия холинергических звеньев в механизмах возбуждения телец Пачини при механическом раздражении.
Таким образом, облегчающие, а в некоторых случаях возбуждающие влияния экзогенного ацетилхолина на механорёцепторы, наблюдавшиеся в проанализированных экспериментах, служат еще одним проявлением весьма широкой распространенности холинореактивных систем в организме (см. обзор: Левтов, 1967). Экзогенный ацетилхолин, взаимодействуя с этими системами нейрональных мембран, вызывает эффекты деполяризационного характера. Однако эти эффекты не имеют прямого отношения к первичному трансформационному процессу, развивающемуся в механорецепторах при их адекватном раздражении.
Влияние тетродотоксина. Известно, что тетродотоксин весьма избирательно блокирует натрий-переносящую систему (см.: Nakamura et al., 1965; Као, 1966; Narahashi, 1972). Благодаря этому тетродотоксин эффективно угнетает импульсную активность, т. е. регенеративную систему генерации. Генерация РП в механорецепторах также в основном определяется натриевыми токами (см. стр. 308). Поэтому важно выяснить особенности изменения РП под влиянием тетродотоксина. Такого рода эксперименты были выполнены на тельцах Пачини у кошки, рецепторах растяжения у ракообразных, мышечных веретенах у лягушки. Однако результаты различных исследователей не полностью согласуются друг с другом.
Так, по одним данным (Loewenstein et al., 1963), указанный агент избирательно действует лишь на натриевые каналы регенеративной системы генерации телец Пачини и рецепторов растяжения (рис. 102). По другим данным (Ozeki, Sato, 1965; Nishi, Sato, 1966), тетродотоксин может постепенно (через 30—60 мин.) угнетать на 40% и деполяризационный РП, т. е. нерегенеративную систему генерации, хотя он не влияет на сопротивление покоящегося нервного окончания телец Пачини, а также на потенциал покоя и на калиевую проводимость. Наконец, существуют данные (Albuquerque, Grampp, 1968) о том, что тетродотоксин не влияет на амплитуду стационарной фазы РП, однако применение его несколько повышает (примерно на 5 мв) потенциал покоя, значительно увеличивает (примерно на 50%) мембранное сопротивление покоящегося рецептора растяжения и сильно угнетает (на 65%) гиперполяризацию, следующую за периодом импульсной активности. В целом авторы данной работы полагают; что тетродотоксин лишь незначительно угнетает натриевые каналы нерегенеративной системы, повышает калиевую проводимость и, возможно, угнетает деятельность электрогенного натриевого насоса, которая, как известно (Nakajima, Takahashi, 1966), в основном и определяет следовую гиперполяризацию в рецепторах растяжения. Отсутствие влияния Или очень небольшое влияние тетродотоксина на РП и генераторный ток в МРОХ было отмечено другими исследователями, в том числе и в опытах с фиксацией потенциала (Wellhoner, 1968, 1970; Klie, Wellhoner, 1973).
Важно указать, что в отличие от угнетающего действия тетродотоксина на следовую гиперполяризацию мёханорецепторов не было замечено его влияния на первичную гиперполяризацию (Nishi, Sato, 1968), т. е. на гиперполяризационный РП (опыты на тельцах Пачини; см. главу 10, рис. 115).
Деполяризационный РП мышечного веретена практически не изменяется тетродотоксином, который, следовательно, не влияет на натриевую проводимость нёрегенеративной системы (Albuquerque et al., 1969). В этой же работе обнаружено, что повышение содержания в среде кальция уменьшает и задерживает угнетающее действие тетродотоксина на импульсную активность рецептора, в то время как отсутствие кальция действовало в обратном направлении.
Тетродотоксин был также неэффективным и при подведении его к механорецепторным волосковым клеткам органов чувств (Katsuki et al., 1966). В этих опытах выявлялись только свойства нерегенеративных систем этих рецепторов. Наличие существенного диффузионного барьера не позволяло данному веществу оказать воздействие на регенеративные компоненты афферентных нервных проводников.
Неэффективность или значительно меньшая эффективность влияния тетродотоксина на нерегенеративную систему в механорецепторах позволяет предположить, что все натриевые ворота (или их большая часть) локализованы на внутренней поверхности рецепторных мембран механочувствительного субстрата, куда нет доступа для данного вещества. Опыты с тетродотоксином четко показывают существование больших различий в свойствах нерегенеративной и регенеративной систем генерации в механорецепторах.
Рис.
102. Влияние тетродотоксина на рецепторный
потенциал и потенциал действия (по:
Loewenstein et al., 1963).

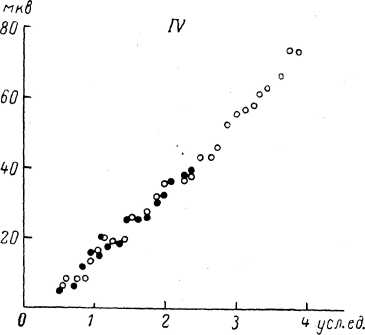

j _ ответы медленно адаптирующегося рецептора растяжения у ракообразных до (а, б) и после (в, г) аппликации тетродотоксина (5 • 10-в г/мл). Сверху вниз: биоэлектрическая активность (внутриклеточная запись от сомы нейрона); то же (запись от аксона на расстоянии около 2 мм от сомы нейрона); механическое растяжение рецептора. Отметка времени — 2 сек.; калибровка — 10 мв. II — ответ декапсулированного тельца Пачини на пороговое механическое раздражение до (б) и после (е) приложения тетродотоксина (5-Ю-4 г/мл). Сверху вниз: биоэлектрическая активность (отведение вблизи немиелинизированного участка нервного окончания); механическое раздражение. Отметка времени — 1 мсек.; калибровка — 15 мкв. Ill, IV — зависимость амплитуды РП от интенсивности раздражения рецептора растяжения (III) и тельца Пачини (IV) до (черные кружки) и после (белые кружки) приложения тетродотоксина. По оси абсцисс — величина ^растяжения (на III — мм, на IV — усл. ед.); по оси ординат — амплитуда РП (на III — мв; на IV — мкв).
20 Физиология механорецепторов
Влияние анестетиков и наркотиков. Общеизвестное угнетающее действие различных анестетиков на нервные структуры, в том числе и на рецепторы, ряд авторов связывает с их проникновением в липидную часть мембран и с возникающим при этом ухудшением проницаемости мембран к ионам натрия и калия (см. обзор: Аничков, 1974). Анестетики, как и многие снотворно-наркотические средства, называют поэтому «стабилизаторами» мембран.
Анестетики резко угнетают механизмы генерации ПД в нервных волокнах (обзор: Ходоров, 1969). На одиночных механорецепторах действие анестетиков было четко выявлено в опытах на” мышечных веретенах (Katz, 1950b), тельцах Пачини (J. А. В. Gray, Sato, 1953; Diamond et al., 1958b; Ильинский, 1963; Sato, Ozeki, 1963), рецепторах растяжения у ракообразных (Eyzaguirre, Kuffler, 1955a). Действие местных анестетйков на тельца Пачини было даже предметом специальных опытов (Nishi, 1968b). Оказалось, что в небольших концентрациях (0.1 —0.3% для мышечного веретена, 0.01—0.75% для телец Пачини при их перфузии) новокаин избирательно блокирует импульсную активность рецепторов, сохраняя в то же время РП неизмененным. Большие дозы анестетика угнетают и РП. Можно думать, что новокаин повышает порог возникновения ПД в претерминальном участке нервного волокна. Увеличение концентрации ионов кальция ослабляло действие новокаина. Ионы натрия не оказывали заметного влияния на эффект (Nishi, 1968b}. Избирательное действие новокаина сейчас широко используется всеми исследователями, анализирующими локальные реакции рецепторных приборов (обзоры: J. А. В. Gray, 1959b; Paintai, 1964; Ильинский, 1967).
Действие наркотиков (эфира, трихлорэтана, хлороформа и др.) изучалось в опытах на легочных рецепторах растяжения (Whit- teridge, Bulbring, 1944; Paintai, 1957; Whitteridge, 1958), каротидных барорецепторах (Robertson et al., 1956) и на ряде других механорецепторов (обзоры: Paintai, 1964, 1971, 1972; Zipf, 1966). Оказалось, что в отличие от местных анестетиков снотворнонаркотические препараты в небольших дозах повышают импуль- сацию, вызванную механическим раздражением, и лишь при увеличении дозы уменьшают или блокируют ее. Интересно, что в период возрастания величины реакции на адекватное раздражение ее порог оставался неизменным. Возможно (Paintai, 1964), что этот эффект обусловлен увеличением скорости восстановления нерегенеративной системы генерации после возбуждения.
Влияние вератридина и аконитина. Эти и другие алкалоиды со сходным действием называют «лабилизаторами» мембран (обзор: Аничков, 1974). Они оказывают значительный возбуждающий эффект на механорецепторы, в частности на механорецепторы сердечно-сосудистой системы (обзоры: Paintai, 1964, 1972), что и определяет широкое использование этих алкалоидов ^в исследованиях. Стимулирующий эффект данных соединений на^возбудимые мембраны обусловлен активацией под их влиянием системы натриевой проводимости (обзор: Ulbricht, 1969).
Анализ действия вератридина и аконитина на механорецепторы, проведенный на рецепторах растяжения у ракообразных, показал (Wellhoner, 1968, 1970), что указанные вещества действуют избирательно на натриевые каналы регенеративной системы, обусловливающей развитие ПД, и не влияют на ионную проницаемость, обеспечивающую развитие генераторного потенциала (РП). В соответствии с этим тетродотоксин, избирательно угнетающий натриевую проводимость регенеративных элементов мембраны, полностью устраняет действие вератридина и аконитина на рецепторы.
Влияние оубаина и других ингибиторов активного транспорта натрия. Оубаин (строфантин G) подавляет активный транспорт натрия (Caldwell, Keynes, 1959). Под влиянием этого вещества исчезает как посттетаническая гиперполяризация, так и вторичные гиперполяризационные сдвиги потенциала в мышечном веретене лягушки (Алексеев, Макаров, 1970; Пошина, 1973). Важно, что ингибиторы энергетического метаболизма 2,4-динитрофенол, азид, цианид также оказывали угнетающее влияние на следовую гиперполяризацию, наступающую после раздражения веретена (Алексеев, Макаров, 1970). Можно думать поэтому, что в основе следовых гиперполяризационных сдвигов, так же как это было показано для рецептора растяжения у ракообразных (см.: Nakajima, Takahashi, 1966), лежит работа электрогенного натриевого насоса, обеспечиваемая энергетическим метаболизмом.
Влияние ионов. В настоящее время изучению ионных механизмов генерации активности в различных рецепторных образованиях уделяется все большее внимание (см. обзоры: J. А. В. Gray, 1959а, 1959Ь; Лебедева, 1965; Ильинский, 1967, 1972а; Fulpius, Baumann, 1969; Ottoson, Shepherd, 1971a; Fuortes, O’Bryan, 1972; Sickel, 1972). Тем не менее, в отличие от синаптических структур рецепторы, и в частности механорецепторы, в этом плане изучены еще явно недостаточно. Более того, при проведении требуемых исследований механорецепторных элементов экспериментаторы сталкиваются с дополнительными трудностями: вспомогательные структуры этих рецепторов существенно затрудняют диффузию ионов и различных веществ в пространстве между нервным окончанием и окружающей его средой, например у телец Пачини и мышечных веретен (J. А. В. Gray, Sato, 1955; Calma, Kidd, 1962; Kidd, 1964; Ильинский, Красникова, 1972, и др.).
I В литературе имеется много работ, посвященных действию ионов на импульсную активность механорецепторов (Matthews, 1931а; Feng, 1933; Talaat, 1933; Dun, Finley, 1938; Fitzgerald, 1940; Ильинский, Лебедева, 1963; Nishi, 1968a; Nier, 1974, ими. др.). Число исследований, которые были осуществлены на одиночных рецепторах с регистрацией их РП, до недавних пор было сравнительно невелико, но существенно возросло за последние годы (Diamond et al., 1958b; Edwards et al., 1963; Ottoson, 1964, 1965a; Calma, 1965; Obara, 1968; Obara, Grundfest, 1968; M. Sato et al., 1968; Albuquerque et al., 1969; Ильинский, Акоев и др., 1971; Husmark, Ottoson, 1971a, 1971c, 1971d; Алексеев, 1972; Акоев, Эльман, 1974; Ильинский, Красникова и др., 1974). Как обычно, основные опыты проводились на тельцах Пачини, мышечных веретенах, рецепторах растяжения у ракообразных.
Ионы натрия и лития. На тельцах Пачини было установлено, что ионы натрия переносят основную часть заряда при генерации РП. При перфузии через капилляры брыжейки кошки безнатриевых растворов амплитуда РП у интактных телец Пачини снижалась на 90% (Diamond et al., 1958b). На 87—90% падала амплитуда РП и при непрерывной внешней перфузии декапсулированных телец Пачини безнатриевым раствором (Ильинский, Акоев и др., 1971; Акоев, Эльман, 1974; Ильинский, Красникова и др., 1974). По другим данным (М. Sato et al., 1968), это падение амплитуды РП было меньше и не превышало 75%. Можно думать, что менее выраженные изменения объяснялись неполным вымыванием ионов натрия в связи с особенностями применявшейся методики.
В опытах различных авторов показано, что в безнатриевых растворах импульсная активность телец Пачини блокировалась очень быстро. Изменения РП наступали позже. После замены без- натриевого раствора нормальным ПД вновь восстанавливался, если время нахождения рецептора в среде без ионов натрия было не очень велико. Учитывая возможную роль начальной части миелинизированного волокна в генерации ПД (стр. 120) и особенности ее васкуляризации (стр. 22), можно полагать, что именно эта область является первоначальной точкой приложения угнетающего влияния безнатриевого раствора.
В экспериментах на мышечных веретенах лягушки было показано (Ottoson, 1964; Calma, 1965; Albuquerque et al., 1969), что амплитуда РП в безнатриевом растворе может снижаться на 70—80%. Параллельно со снижением величины РП падала и скорость нарастания РП. В целом уменьшение статической фазы РП происходило в большей степени, чем ослабление динамического компонента (Husmark, Ottoson, 1971с). При замене безнатриевых растворов на обычные РП восстанавливался. Если РП, хоть и ослабленные, наблюдались при довольно продолжительном (например, 30-минутном) действии безнатриевого раствора (значение существующего диффузионного барьера), то импульсная активность менялась очень быстро. Спонтанные разряды ПД быстро урежались и полностью исчезали. Уменьшалась и вызванная активность, которая к 3—10 мин. воздействия полностью блокировалась. РП в этот период еще оставался неизменным. Блокирование импульсной активности и постепенное уменьшение ГП в безнатриевых растворах отмечалось и в опытах на рецепторах растяжения у ракообразных (Obara, Grundfest, 1968).
В безнатриевых растворах величина тока (опыты с фиксацией напряжения), вызванного растяжением, падала до 10% от исходного значения (Klie, Wellhoner, 1973). В безнатриевых растворах, содержащих трис (гидроксиметиламинометан), амплитуда ГП рецепторов растяжения была весьма значительной, составляя 30—5О°/о от величины в контроле (ОЬага, 1968).
На основании изложенных фактов можно заключить, что ионы натрия переносят основную часть зарядов при генерации как ПД, так и РП. Натриевые каналы рецепторной механочувстви- тельной мембраны оказываются более стойкими к изъятию натрия из внешней среды, чем каналы регенеративных структур рецептора и отходящего нервного волокна. Это подтверждается также и другими опытами (см. выше о действии тетродотоксина, стр. 303).
Известно, что ионы лития, весьма сходные по своим свойствам с ионами натрия, могут их заменять в процессах генерации электрической активности нервными и мышечными элементами (Overton, 1902; Gallego, Lorente de N6, 1951; Huxley, Stam- pfli, 1951, и др.). Эксперименты с литием могут дать сведения о специфичности натриевых каналов в мембране нервного окончания.
Эксперименты с мышечным веретеном не дали однозначных результатов. Одни авторы (Albuquerque et al., 1969) обнаружили, что литий очень эффективно заменяет натрий, так что амплитуда деполяризационного РП при данной замене не менялась. По другим данным (Алексеев, Макаров, 1969), амплитуда генераторного потенциала уменьшалась. Исчезала и посттетаническая гиперполяризация.
В экспериментах с тельцами Пачини выяснилось, что ион лития может замещать ион натрия лишь частично. В литиевом растворе амплитуда РП заметно (на 30%) падала. После перехода от литиевого раствора к обычному РП восстанавливался полностью (М. Sato et al., 1968). В литиевом растворе пороги возникновения ПД при механическом раздражении прогрессивно возрастали и ответы исчезали полностью, хотя и медленнее, чем в без- натриевом растворе, в который литий не добавлялся (Nishi, 1968а). Изменения в генерации ПД, по-видимому, были связаны как со снижением амплитуды РП, так и с длительной деполяризацией мембраны нервного окончания, которую, как известно (Keynes, Swan, 1959; Bohm, Straub, 1962, и др.), ионы лития могут вызывать, накапливаясь внутри клетки.
В опытах с рецепторами растяжения также было установлено, что ионы лития лишь частично заменяют ионы натрия: амплитуда РП в литиевых растворах снижалась по сравнению с нормой (Obara, Grundfest, 1968), частота импульсации рецептора уменьшалась, а при полной замене натрия на литий она блокировалась (Giacobini, Stepita-Klauco, 1970). Этот эффект также можно было объяснить накоплением лития в рецепторе и его влиянием на деятельность натриевого насоса. Каналы, обеспечивающие движение литиевых ионов через мембрану в клетку, оказались чувствительными к влиянию находящихся в среде ионов калия (Gia- cobini et al., 1970). В целом следует заключить, что ионы лития, по-видимому, не могут полностью заменить ионы натрия для механизмов, обеспечивающих перенос заряда в процессе генерации электрической активности механорецепторов.
Ионы калия. Исследование действия этих ионов на механорецепторы представляется важным не только потому, что они могут участвовать в генерации РП, но и потому, что, как показали многочисленные исследования (подробнее см. главу 7), в среде, окружающей высокоспециализированные механорецепторы (например, волосковые клетки органа слуха), содержится значительное количество именно ионов калия.
Действие ионов калия исследовалось в опытах на различных механорецепторах. В этих экспериментах было установлено, что ионы калия могут весьма значительно менять возбудимость механорецепторов к действию адекватного раздражения. Эффект существенно зависел от концентрации калия, и определялся суммой изменений, наблюдаемых как в нерегенеративной, так и в регенеративной системах генерации.
Анализ изменений возбудимости, проведенный на тельцах Пачини, показал (Акоев, Эльман, 1974; Ильинский, Красникова и др., 1974), что некоторое (примерно до 12—13 ммолей/л) увеличение концентрации калия приводило к двум эффектам. Наблюдалось небольшое уменьшение амплитуды деполяризационного РП, возникающего в ответ на механическое раздражение, а также происходило снижение критического уровня деполяризации, необходимой для генерации ПД. Второй эффект превалировал над первым, и возбудимость рецептора в целом возрастала (рис. 103). Дальнейшее увеличение содержания ионов калия приводило к возрастанию порогов генерации ПД и полному блоку импульсной активности (например, при концентрации 22.4 ммолей/л, см. также: Nishi, 1968а). Возрастание порогов и блокирование генерации ПД явилось, по-видимому, результатом инактивации натриевых каналов, вызванной нарастающей деполяризацией структур рецептора, аналогичной той, которая имеет место при действии калия в других возбудимых образованиях (Huxley, Stampfli, 1951; Ritchie, Straub, 1957; Stampfli, 1959, и др.; см. также обзоры: Cole, 1968; Ходоров, 1969). Уменьшение амплитуды ГП происходило прежде всего вследствие снижения величины мембранного потенциала. Известно, что величина мембранного потенциала и амплитуда РП у механорецепторов связаны линейной зависимостью (см. например: Terzuolo, Washizu, 1962). Уменьшение мембранного потенциала, снижение сопротивления мембраны и уменьшение РП при увеличении наружной концентрации ионов калия были показаны на рецепторе растяжения у ракообразных (Edwards et al., 1963).
Прогрессивное снижение амплитуды РП при увеличении содержания ионов калия в среде отмечалось и в опытах на мышечных веретенах (Husmark, Ottoson, 1971с). При этом уменьшения величин статической составляющей ответа были более значительными, чем динамического компонента. Важно заметить, что несмотря на падение амплитуды ГП у мышечных веретен в некотором диапазоне концентраций калия (примерно до 12 ммолей/л) пороги возникновения ПД при действии механического раздражения снижались (Kidd et а]., 1971) аналогично рассмотренным выше данным по тельцам Пачини. Уменьшение порогов свидетельствует о превалировании повышения возбудимости регенеративной системы генерации над уменьшениями ГП и в случае мышечных веретен.
Рис. 103. Зависимость возбудимости тельца Пачини от концентрации ионов калия.
По оси абсцисс — концентрация КС1 (ммоль/л); по оси ординат — сила порогового для генерации ПД механического раздражения (усл. ед.). Стрелка — концентрация К+ (22.4 ммоля/л), при которой импульсные ответы рецептора отсутствуют, по данным работы Ниши (Nishi, 1968а).
В ходе ритмического раздражения телец Пачини стимулами умеренной частоты отмечается снижение порогов (Ильинский, 1966а). Высказывалось предположение, что это явление вызвано накоплением ионов калия около регенеративных элементов рецептора. Функциональное возрастание концентрации калия в среде при работе, по-видимому, может иметь место и в случае мышечных веретен (Kidd et al., 1971). Эффекты добавления ионов калия на механорецепторы возрастали при исходном уменьшении содержания кальция в среде (Matthews, 1931а; Fitzgerald, 1940; Ильинский, Лебедева, 1963; Edwards et al., 1963; Calma, 1965; Nier, 1974).
Уменьшение содержания калия в среде вызывало уменьшение амплитуды ГП мышечных веретен (Husmark, Ottoson, 1971а, 1971с) и телец Пачини (Акоев, Эльман, 1974). В ответах мышечных веретен на действие механического стимула под влиянием удаления калия в большей степени менялась динамическая составляющая ответа, которая могла полностью исчезнуть. Эти данные говорят об участии ионных механизмов в явлениях ранней адаптации ГП мышечного веретена (Husmark, Ottoson, 1971а, 1971с).6 Удаление ионов калия из среды временно повышало возбудимость мышечных веретен, однако затем пороги возрастали, и импульсная активность полностью блокировалась (Husmark, Ottoson, 1971а, 1971d).
Исследование действия ионов калия проводилось и на волосковых рецепторах канальных, ямковых и ампулированных органов боковой линии акул (Katsuki, Hashimoto, 1969; Katsuki et al., 1970, 1971; Hashimoto, Katsuki, 1972; Onoda, Katsuki, 1972; обзор: Katsuki, 1973). Наиболее четкие данные были получены на рецепторах ямковых органов, которые у акул имеют все морфологические характеристики обычных механорецепторных клеток (каждая клетка имеет одну полярно расположенную кино- цилию и пучок стереоцилий; все рецепторы отдельного ямкового органа имеют одинаковую ориентацию), но обладают достаточно высокой чувствительностью как к механическим, так и к электрическим раздражающим стимулам. Эти эксперименты показали, что механическая и электрическая чувствительность рецепторов, тестируемая по величине импульсного ответа, значительно увеличивается при возрастании концентрации калия в окружающей среде по сравнению с содержанием его в морской воде.7
В экспериментах на органе слуха морских свинок было показано, что при замене богатой калием эндолимфы на растворы, содержащие небольшое количество калия, кохлеарные ответы (микрофонные и суммационные потенциалы, а также импульсная активность) уменьшаются, равно как и величина эндокохлеарного потенциала (Konishi et al., 1966). Эти данные, как и результаты экспериментов на рецепторах боковой линии, позволяют говорить о важной роли ионов калия в механизмах деятельности волосковых механорецепторов органов чувств (подробнее этот вопрос рассматривается в главе 7).
Результаты экспериментов по действию ионов калия показывают, что эти ионы могут оказывать достаточно существенное влияние на возбудимость механорецепторов и в силу этого могут быть своего рода регуляторами деятельности сенсорных приборов.
Ионы кальция и магния. Исследование влияния этих ионов на деятельность механорецепторов проводилось в экспериментах на мезентериальных тельцах Пачини (Ильинский, Лебедева, 1963; Nishi, 1968а; Акоев и др., 19746), мышечных
В этой связи следует упомянуть также данные, полученные на рецепторе растяжения у ракообразных (Ottoson, 1973) и в экспериментах на мышечных веретенах у земноводных с применением тетраэтил аммония (Алексеев, 1972), который угнетает калиевую проницаемость возбудимых мембран (Schmidt, Stampfli, 1966; Koppenhofer, Vogel, 1969).
О влиянии различных ионов на специализированные образования органов боковой линии см. также другие работы (Loewenstein, Ishiko, 1962; Roth, 1971; Nier, Hensel, 1973).
веретенах (Matthews, 1931а; Calma, 1965; Ottoson, 1965a; Albuquerque et al., 1969), вибриссах (Fitzgerald, 1940; Nier, 1974), тактильных рецепторах (Talaat, 1933; К. R. Smith, Creech, 1967), рецепторах растяжения (Edwards et al., 1963; Washizu, 1965; Obara, 1968). Уменьшение содержания ионов кальция в среде повышало возбудимость механорецепторов. Порог генерации ПД снижался, и даже могла появиться фоновая импульсация рецепторов в отсутствие механического раздражения (Ильинский, Лебедева, 1963; Ottoson, 1965а). В соответствии с этим увеличение концентрации кальция в среде приводило к снижению возбудимости механорецепторов.
Такие влияния ионов кальция определялись изменениями системы генерации ПД, которые превалировали над изменениями нерегенеративных структур, связанных с генерацией РП. Уменьшение содержания кальция приводило к снижению амплитуды деполяризационного РП мышечного веретена и рецептора растяжения у ракообразных. На мышечном веретене было установлено, что отсутствие ионов кальция вызывает снижение амплитуды ГП на 60—80% и даже более (Calma, 1965; Ottoson, 1965а). Этот эффект был особенно хорошо выражен при обработке рецептора безнатриевым раствором или новокаином. Полного восстановления ГП после угнетающего действия бескальциевого раствора не наблюдалось. При удалении ионов кальция сопротивление мембраны рецептора растяжения снижалось (Edwards et al., 1963; Obara, 1968).
При умеренном повышении концентрации кальция амплитуда ГП менялась незначительно (могла даже кратковременно несколько возрасти). Однако при значительном увеличении содержания кальция (в 5—15 раз и более) величина ГП мышечного веретена (Ottoson, 1965а) и рецептора растяжения (Obara, 1968) прогрессивно падала. Увеличение концентрации ионов кальция ускоряло развитие процесса адаптации механорецепторов (Fitzgerald, 1940), а уменьшение содержания кальция в среде замедляло развитие этих явлений (Talaat, 1933), что было, несомненно, связано с действием кальция на регенеративную систему генерации.
Ионы магния уменьшали или предотвращали развитие действия бескальциевых растворов (Calma, 1965). Под влиянием ионов магния амплитуда ГП телец Пачини возрастала (Акоев и др., 19746).
Ионы хлора. Влияние этих ионов исследовалось в экспериментах на рецепторах растяжения у ракообразных (Edwards et al., 1963; Obara, 1968). Согласно полученным данным, можно заключить, что ионы хлора не принимают участия в процессах генерации ГП.8
Следует заметить, что эксперименты на рецепторах растяжения говорят об отсутствии у этих сенсорных образований (в отлцчие, например, от инкапсулированных механорецепторов позвоночных) сколько-нибудь
* * *
В данной главе были представлены исследования свойств отдельных элементов механорецепторных приборов, проводившиеся с использованием физических факторов среды (электрический ток, температура и др.), а также с применением ряда химических веществ. Эти материалы показали, что наибольшей резистентностью обладают вспомогательные структуры. Они практически не изменяют своих свойств иод влиянием использовавшихся физических воздействий. Они также оказались значительно менее чувствительными по сравнению с нерегенеративными и регенеративными элементами механорецепторных приборов к действию изучавшихся химических факторов. Лишь в тех случаях, когда некоторые из химических агентов, введенные в достаточно высоких дозах, могли вызывать сокращение контрактильных элементов вспомогательных структур, равно как и окружающих^тканей, наблюдались изменения ответной реакции механорецепторов на действие механического раздражения или даже обнаруживалось возникновение возбуждения рецепторов в отсутствие адекватного стимула.
Приведенные материалы выявили также, что вспомогательные структуры и окружающие ткани могут создавать диффузионный барьер, затрудняющий проникновение химических агентов из жидкостей тела к рецептирующим элементам, причем в разной степени относительно различных элементов механоре- цепторного прибора. Регенеративные структуры генерации ПД рецепторов оказываются более доступными для действия химических веществ, чем нерегенеративные структуры, ответственные за возникновение РП. Это обстоятельство, а также особенности функционирования регенеративных структур (на возникновение ПД сказываются изменения в свойствах как нерегенеративной, так и регенеративной систем генерации) делают данное звено в цепи процессов наиболее чувствительным при действии на механорецепторы различных факторов. Поэтому в конечном счете именно за счет реагирования регенеративной системы происходят практически все изменения в ответных реакциях механорецепторов при действии на них физических и химических факторов. Следует отметить также, что в целом деятельность регенеративной системы генерации механорецепторов не обнаруживает каких- либо специфических черт, отличных от аналогичных процессов, присущих мембранам обычных нервных волокон.
Наибольшим сходством в механизмах деятельности нерегенеративных и регенеративных систем генерации является то, что в обоих случаях развитие деполяризационных процессов связано с перемещением ионов натрия, однако «ворота» натриевых кана-
существенных диффузионных барьеров. Это дает основание полагать, что вокруг данных механорецепторов отсутствует специфическая микросреда (см. главу 7).
лов в нерегенеративных и регенеративных структурах локализуются, по-видимому, на разных поверхностях возбудимых мембран. Очевидно, именно данное обстоятельство обусловливает изложенные выше особенности действия разных химических веществ (например, тетродотоксина и анестетиков) на эти структуры.
Было показано также, что важной особенностью структур, связанных с явлениями трансформации в механорецепторах, является их электрическая невозбудимость. Напротив, процессы, обусловливающие генерацию РП, так же как и возникновение ПД, демонстрируют отчетливую зависимость от действия поляризующего электрического тока. Из представленных данных следует также, что трансформационные процессы и явления генерации биоэлектрической активности в их отношении к температурному фактору не демонстрируют четко выраженных особенностей, позволяющих однозначно охарактеризовать механизмы, лежащие в их основе.
Следует констатировать, что свойства отдельных звеньев процессов возбуждения в механорецепторах и прежде всего трансформационных явлений, а также процессов генерации РП изучены еще недостаточно. В частности, в настоящее время не представляется возможным ответить на вопрос: имеется ли какое-либо различие в трансформационных процессах у механорецепторов, относящихся к различным типам (например, таких как в органах чувств и в тканях у позвоночных, т. е. обладающих специализированными — реснитчатыми — структурами или же лишенных их). Не удается пока однозначно резюмировать в обсуждающемся здесь аспекте многие представленные выше описательные данные о действии ионизирующего излучения, ультразвука и других изучавшихся факторов. Зато достаточно существенные выводы представляется возможным сделать на основе изложенных фактов об изменении возбудимости механорецепторов под влиянием изменений концентрации ионов. Для этого вывода, конечно, необходима прежде всего детальная оценка естественных концентраций веществ в среде, непосредственно окружающей механо- рецепторные структуры, и именно эти вопросы будут анализироваться в главе 7.
Анализ действия физических и химических факторов на тканевые механорецепторы позвоночных позволяет утверждать, что местом первичных трансформационных процессов у этих чувствительных образований являются сами окончания сенсорных нейронов, а не окружающие их клеточные элементы. Этот вопрос более подробно будет рассматриваться в главе 8 (стр. 388).