

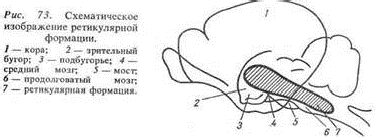
Вопрос47. Ретикулярная формация ствола головного мозга
Ретикулярная формация (лат. rete - сеть) представляет собой совокупность клеток, клеточных скоплений и нервных волокон, расположенных на всем протяжении ствола мозга (продолговатый мозг, мост, средний и промежуточный мозг) и в центральных отделах спинного мозга . Ретикулярная формация получает информацию от всех органов чувств , внутренних и других органов , оценивает ее, фильтрует и передает в лимбическую систему и кору большого мозга. Она регулирует уровень возбудимости и тонуса различных отделов центральной нервной системы, включая кору большого мозга, играет важную роль в сознании, мышлении, памяти, восприятии, эмоциях, сне, бодрствовании, вегетативных функциях, целенаправленных движениях, а также в механизмах формирования целостных реакций организма. Ретикулярная формация прежде всего выполняет функцию фильтра, который позволяет важным для организма сенсорным сигналам активировать кору мозга, но не пропускает привычные для него или повторяющиеся сигналы.
Ретикулярная формация представляет собой важный пункт на пути восходящей неспецифической соматосенсорной системы . Соматовисцеральные афференты идут в составе спиноретикулярного тракта ( переднебоковой канатик ), а также, возможно, в составе проприоспинальных (полисинаптических) путей и соответствующих путей от ядра спинального тройничного тракта . К ретикулярной формации приходят также пути от всех других афферентных черепномозговых нервов , т.е. практически от всех органов чувств. Дополнительная афферентация поступает от многих других отделов головного мозга - от моторных областей коры и сенсорных областей коры , от таламуса и гипоталамуса . Имеется также множество эфферентных связей - нисходящие к спинному мозгу и восходящие через неспецифические таламические ядра к коре головного мозга , гипоталамусу и лимбической системе . Большинство нейронов образует синапсы с двумя - тремя афферентами разного происхождения, такая полисенсорная конвергенция характерна для нейронов ретикулярной формации. Другими их свойствами являются большие рецептивные поля поверхности тела, часто билатеральные, длительный латентный период ответа на периферическую стимуляцию (вследствие мультисинаптического проведения), слабая воспроизводимость реакции (стохастические колебания числа потенциалов действия при повторной стимуляции). Все эти свойства противоположны свойствам лемнисковых нейронов в специфических ядрах соматосенсорной системы ( рис.9-7 и рис. 5-13 ).
Функции ретикулярной формации изучены не полностью. Считается, что она участвует в следующих процессах:
1. в регуляции уровня сознания путем воздействия на активность корковых нейронов , например, участие в цикле сон / бодрствование ,
2. в придании аффективно-эмоциональной окраски сенсорным стимулам, в том числе болевым сигналам , идущим по переднебоковому канатику , путем проведения афферентной информации к лимбической системе ,
3. в вегетативных регулирующих функциях, в том числе во многих жизненно важных рефлексах ( циркуляторных рефлексах и дыхательных рефлексах , рефлекторных актах глотания , кашля , чихания ), при которых должны взаимно координироваться разные афферентные и эфферентные системы,
4. в целенаправленных движениях в качестве важного компонента двигательных центров ствола мозга .

Вопрс48. Сравнительная характеристика кабельного и сальтаторного видов проведения возбуждения
Нервная ткань обладает таким физиологическим свойством как проводимость, т. е. способностью проводить возбуждение по ходу нервного волокна в виде потенциала действия. Выделяют два вида проведения возбуждения в зависимости от строения нервного волокна. Различают два вида нервных волокон: мякотные (миелиновые) и безмякотные (немиелиновые). В безмякотных нервных волокнах наблюдается непрерывное распространение возбуждения, в основе которого лежат локальные или круговые токи. Как было сказано раньше, возбужденный электроотрицательный участок нервного волокна становится раздражителем для ближайшего невозбужденного электроположительного участка, который возбуждается (возбужденный участок как бы разряжается в сторону невозбужденного, следствием чего и являются появление локальных или круговых токов).
Миелин, прокрывающий нервное волокно, располагается сегментами, т. е. прерывисто. Миелин - хороший изолятор и, если бы он сплошным слоем покрывал нервное волокно, то возбуждение не распространялось бы. Миелиновая оболочка образуется клетками неврилеммы или шванновскими клетками. Плазматическая мембрана одной шванновской клетки обвертывает спирально в несколько слоев участок аксона, длиной в несколько сотых микрон. Между участками аксона, покрытого миелиновой оболочкой, остаются немиеленизированные зоны. Эти зоны называются перехватами Ранвье.
В волокнах, покрытых миелиновой оболочкой (мякотные волокна) возбуждение распространяется скачкообразно (сальтаторно), т. е. по перехватам Ранвье. Как было показано японским физиологом Тасаки, это создает своеобразную систему надежности для распространения возбуждения (разности потенциалов между возбужденным и невозбужденным участками волокна хватает на 5-6 перехватов Ранвье). В случае, если появится повреждение на небольшом участке волокна вследствие большого электрического поля распространение потенциала действия не нарушается. Как известно, начальная часть аксона в нервной клетке не покрыта миелиновой оболочкой. Именно в этом участке волокна и формируется потенциал действия. Возникает разность потенциалов между возбужденным и невозбужденным участком первого перехвата Ранвье, который под влиянием этого электрического поля возбуждается. Затем разность потенциалов формируется между возбужденным первым перехватом Ранвье и следующим, который перезаряжается и возбуждение приобретает распространяющий характер. Таким образом, в основе распространения возбуждения по мякотному волокну, как и безмякотному, лежат также местные (круговые, вихревые) токи. В перехватах Ранвье, находящихся друг от друга на расстоянии 2 мм, обнаружена большая плотность натриевых каналов - до 1200 на 1 мкм2, что значительно облегчает проведение возбуждения по нервному волокну. Прерывистое распространение возбуждения имеет некоторые преимущества по сравнению с непрерывным. Во-первых, скорость распространения возбуждения в волокнах, покрытых миелином, в 8-10 раз быстрее, чем в безмякотных. Во-вторых, на распространение возбуждения прерывистого типа затрачивается меньше энергии, оно более экономично, что, по всей вероятности, связано с большой плотностью натриевых каналов в перехватах Ранвье.
При распространении возбуждения по нервному волокну следует учитывать чисто физические или кабельные свойства проводника (нерв можно представить как кабель, помещенный в морскую воду). К кабельным свойствам относится, в частности, диаметр (поперечное сечение) проводника - чем толще нервное волокно (или больше поперечное сечение), тем меньше сопротивление. Следовательно, тем быстрее будет распространение возбуждения в виде импульса. Большое значение при возбуждении имеет также емкость и сопротивление мембраны. Так, если входное сопротивление мембраны больше, то и возбудимость в этом месте уменьшается. К кабельным свойствам относится также и электротон, оказывающий большое влияние на проводимость: чем выраженнее катэлектротон, тем быстрее проводится потенциал действия. Анэлектротонические изменения, напротив, ухудшают проведение возбуждения по нервной ткани.
В зависимости от скорости проведения возбуждения все нервные волокна делятся на три группы: А, В и С. Нервные волокна группы А - это высокоскоростные волокна, исключительно мякотного типа. В зависти от сечения нервного волокна скорость проведения возбуждения их колеблется в пределах 20-120 м/с. Различают А- волокна - самые скоростные - 70-120 м/с (диаметр волокна 12-20 мкм - a-волокна, их средняя скорость проведения возбуждения составляет 70-120 м/с; диаметр 8-12 мкм - b-волокна, проводящие возбуждение со скоростью 40-70 м/с; диаметр волокна 4-8 мкм - g-волокна, проводящие возбуждение со скоростью 20-40 м/с). Таким образом, чем толще проводник, тем больше скорость проведения возбуждения. Нервные волокна группы В представляют собой в основном безмякотные волокна, скорость распространения возбуждения которых составляет 6-20 м/с. Нервные волокна группы С представлены исключительно безмякотными волокнами вегетативной природы, скорость проведения возбуждения их составляет 0,5-6 м/с.
В физиологии имеется три закона распространения возбуждения.
Закон целостности нерва (закон непрерывности). Нерв проводит возбуждение только в том случае, если он сохраняет свою гистологическую и функциональную целостность. Любые отклонения этих показателей приводят к нарушению его проводимости. Действие местных анестетиков (новокаин) основано на том, что молекулы новокаина блокируют натриевые каналы, в результате чего прекращается натриевый ток и ткань теряет способность возбуждаться. Возбуждение при раздражении болевых рецепторов доходит до места, где действует новокаин и блокируется, вследствие чего болевые импульсы не достигают болевого центра.
Закон двустороннего проведения возбуждения. Нервное волокно способно проводить возбуждение от рецепторов к центрам и наоборот, от центров к периферическим образованиям. Такая закономерность была показана в классических исследования Кюне и Бабухина. Так, опыт Кюне заключался в следующем: если нарушить целостность мышцы между двумя ее участками, которые иннервируются двумя разветвлениями одного аксона, то электрическое раздражение любого из ответвлений аксона приводит к сокращению обеих частей мышцы.
Закон изолированного распространения возбуждения. Известно, что потенциал действия в волокнах, покрытых миелином, не перебрасывается с одного нервного волокна на другое благодаря хорошим изоляционным свойствам миелина. Такое изолированное проведение возбуждения обеспечивает мелкие и точные профессиональные сокращения мышц (игра на пианино, работа часового мастера и др.). Сразу после рождения достаточная миелинизация нервных волокон отсутствует и на любое раздражение новорожденные в большинстве случаев отвечают не локальными, а диффузными сокращениями большой группы мышц. Подобная же ответная реакция наблюдается во всех гладких мышцах, которые иннервируются безмякотными нервными волокнами, не обладающими изоляционнымитсвойствами.