

Genetika_Razvitija
.pdfГлава 3. Раннее развитие млекопитающих
______________________________________________________________________________
недоступной для другим спермиев. Блестящая оболочка растворяется протеолитическими ферментами. Ее устойчивость к этим ферментам существенно возрастает после оплодотворения, что, вероятно, обусловлено действием содержимого кортикальных гранул, попавшего в перивителиновое пространство. Предполагают, что овопероксидаза кортикальных гранул модифицирует ZP3 и, помимо этого, связываясь с остатками тирозина, модифицирует и ZP2, что и приводит к затвердеванию блестящей оболочки.
Благодаря наличию видоспецифических рецепторов, блестящая оболочка препятствует связыванию с ней чужеродных спермиев.
Блестящая оболочка не исчезает после оплодотворения и сохраняется вокруг зародыша на протяжении всего доимплантационного периода (зародыши мышей, крыс, хомячков) или в течение значительной части этого периода (зародыши кроликов, овец, свиней и др.). Благодаря блестящей оболочке бластомеры дробящегося зародыша располагаются компактно и упорядоченно, ориентируясь в ограниченном трехмерном пространстве, существующем внутри блестящей оболочки.
Это имеет немаловажное значение для взаимодействий бластомеров и образования между ними максимального количества контактов, что способствует нормальной компактизации и поляризации бластомеров. Если удалить блестящую оболочку, то дробление бластомеров будет продолжаться, однако во многих
случаях они располагаются в виде цепочки, и компактизация полностью нарушается либо резко запаздывает. Кроме того, блестящая оболочка мешает случайному слипанию соседних зародышей, то есть предотвращает образование спонтанных аггрегационных химер, а также прилипанию зародышей к клеткам яйцевода или матки. Необходимо иметь в виду, что на начальных стадиях дробления, вплоть до стадии бластоцисты (наружный слой которой представлен трофэктодермой), бластомеры обладают высокой адгезивностью. Если трансплантировать зародыш, лишенный блестящей оболочки, в яйцевод, то при этом неминуемо происходит адгезия бластомеров с эпителиальными клетками стенки яйцевода и зародыш гибнет.
Фолликулярные клетки, примыкающие непосредственно к блестящей оболочке, формируют лученостный венец (corona radiata), переходящий без резкой границы в яйценосный бугорок (сumulus oophorus), он образован более диффузно распределенными фолликулярными клетками, которые расположены на большем расстоянии от яйца. Клетки лучистого венца и яйценосного бугорка погружены в матрикс, состоящий преимущественно из гиалуроновой кислоты и гликопротеинов.
Гиалуровая кислота (называемая также гиалуронатом или гиалуронаном) относится к глюкозаминогликанам, которые представляют собой длинные неразветвленные полисахаридные
___________________________________________________________________________________
3-25
Глава 3. Раннее развитие млекопитающих
______________________________________________________________________________
цепи, состоящие из повторяющихся |
что локальное увеличение синтеза |
|||||||||||
дисахаридных |
звеньев. |
|
Ýòè öåïè |
гиалуроновой |
|
кислоты, |
||||||
недостаточно гибки, |
чтобы склады- |
притягивающей воду и тем самым |
||||||||||
ваться в компактные глобулярные |
вызывающей набухание матрикса, |
|||||||||||
структуры, и кроме того в высокой сте- |
облегчает |
миграцию клеток при |
||||||||||
пени гидрофильны, поэтому они |
морфогенезе и регенерации. |
|
||||||||||
стремятся принять конформацию |
Благодаря |
наличию |
вокруг |
|||||||||
очень рыхлого, неупорядоченного |
овулировавшего ооцита лученосного |
|||||||||||
клубка, который занимает |
огромный |
венца и кумулюса, |
возрастает общая |
|||||||||
äëÿ |
своей массы объем, и образуют |
масса клеток, |
è |
ýòî, à |
также |
|||||||
гели даже в очень низких |
физические свойства матрикса, в |
|||||||||||
концентрациям. Благодаря высокой |
который погружены все эти клетки, |
|||||||||||
плотности отрицательных зарядов, их |
способствует захвату овулировавших |
|||||||||||
молекулы притягивают множество |
яйцеклеток фимбриями фаллопиевой |
|||||||||||
таких осмотически активных ионов, |
трубы. Кроме того, кумулюс играет |
|||||||||||
êàê Nà+, что ведет к всасыванию в |
существенную роль в транспорте |
|||||||||||
матрикс большого количества воды. |
яйцеклеток по ампулярной части |
|||||||||||
Ýòî |
создает |
давление |
набухания |
яйцевода и |
нужен для того, |
чтобы |
||||||
(тургор), |
позволяющее |
|
матриксу |
реснички |
эпителиальных |
клеток |
||||||
противостоять сжимающим силам, |
фаллопиевой трубы могли участвовать |
|||||||||||
обеспечивая ткани механическую |
в перемещении яйцеклеток. Когда |
|||||||||||
опору, и, в то же время, не препятствуя |
яйцеклетки |
достигают |
места |
|||||||||
быстрой диффузии водорастворимых |
оплодотворения, они обычно теряют |
|||||||||||
молекул |
è |
|
миграции |
клеток. |
значительную часть клеток кумулюса, |
|||||||
Гиалуроновая кислота состоит из |
причем это происходит под влиянием |
|||||||||||
повторяющейся |
последовательности |
гиалуронидазы спермиев или в ре- |
||||||||||
несульфатированных дисахаридных |
зультате механического действия |
|||||||||||
единиц |
è |
может |
содержать до |
ресничек |
эпителия фаллопиевой |
|||||||
нескольких тысяч сахарных остатков. |
трубы. |
|
|
|
|
|||||||
Îíà |
встречается |
â |
различных |
Процесс утраты яйцеклетками |
||||||||
количествах во всех тканях и |
кумулюса называется денудацией (от |
|||||||||||
жидкостях тела взрослых животных. |
англ. denudation - обнажение). Процесс |
|||||||||||
Но особенно много ее у ранних |
денудации видоспецифичен, у одних |
|||||||||||
зародышей. Гиалуроновая кислота |
видов млекопитающих происходит |
|||||||||||
выполняет особую функцию там, где |
раньше, чем у других, и это не коррели- |
|||||||||||
происходит |
|
миграция |
клеток, |
рует с тем временем, когда зародыши |
||||||||
например, |
|
â |
|
процессах |
попадают в матку. |
Кумулюс может |
||||||
эмбрионального развития и при |
сохраняться 10-16 ч и растворяется |
|||||||||||
заживлении ран. В периоды клеточной |
только после оплодотворения, а у |
|||||||||||
миграции она |
образуется |
|
в больших |
яйцеклеток крыс и кроликов исчезает |
||||||||
количествах, |
а после прекращения |
через 4-6 ч после овуляции, причем вне |
||||||||||
миграции ее избыток разрушается |
зависимости от оплодотворения. У |
|||||||||||
ферментом гиалуронидазой. Полагают, |
яйцеклеток собак и котов кумулюс |
|||||||||||
___________________________________________________________________________________
3-26
Глава 3. Раннее развитие млекопитающих
______________________________________________________________________________
частично дезинтегрируется после пенетрации спермиями, но 1-3 слоя фолликулярных клеток остаются связанными с блестящей оболочкой на протяжении первых делений дробления.
Слой цитоплазмы, располагающийся непосредственно под плазматической мембраной яйца, называется кортикальным слоем или кортексом. Цитоплазма этого слоя имеет более упорядоченное строение, чем цитоплазма более глубоких слоев,
цитоскелета, что в свою очередь приводит к мозаичной организации плазматической мембраны. Примерно от одной десятой до одной пятой общей поверхности яйцеклетки мыши представлено «лысым» районом, в котором нет микроворсинок. В субкортикальном районе этой области яйцеклетки располагается густая сеть микрофиламентов, а глубже находится мейотическое веретено метафазы II. У других млекопитающих «лысый» район также соответствует той области
èв ней содержатся высокие цитоплазмы, где располагается
концентрации глобулярных молекул актина. При оплодотворении молекулы актина полимеризуются, образуя длинные тяжи микрофиламентов.
Микрофиламенты необходимы для осуществления клеточного деления; кроме того, они участвуют в образовании выростов, увеличивающих клеточную поверхность, - микроворсинок, которые способствуют проникновению спермия в клетку. Кортикальная цитоплазма гораздо более вязкая, чем расположенная глубже основная масса цитоплазмы, и она не смещается при центрифугировании. В кортексе заключены кортикальные гранулы. Эти мембранные структуры гомологичны акросомному пузырьку спермия, так как они содержат протеолитические ферменты и формируются аппаратом Гольджи. Однако, если каждый спермий имеет один акросомный пузырек, то яйцо, например, у морского ежа содержит приблизительно 15 тыс. кортикальных гранул.
Для овулировавших яйцеклеток млекопитающих характерна своеобразная организация
мейотическое веретено.
Лишь у немногих видов (собаки, лисицы) овулируют незрелые яйцеклетки во время поздней профазы I мейоза, в таких яйцеклетках хорошо заметно ядро (зародышевый пузырек). Эти яйцеклетки созревают после пенетрации спермиями, однако деконденсация головки спермия осуществляется лишь после завершения яйцеклеткой первого мейотического деления. У преобладающего большинства млекопитающих овулируют яйцеклетки, заблокированные на метафазе второго мейотического деления.
3.1.11. Процесс оплодотворения у млекопитающих.
Яйцеклетки млекопитающих оплодотворяются в ампулярной части яйцевода, куда доходит небольшое количество спермиев, хотя при эякуляции в половой тракт самки попадает многие миллионы спермиев. Например, у мыши из 50 млн. спермиев эякулята в ампуле яйцевода
___________________________________________________________________________________
3-27
Глава 3. Раннее развитие млекопитающих
______________________________________________________________________________
обнаруживается меньше 100, а у |
Снижение содержания холестерина |
||||||||||||||
человека из 200 млн. спермиев ампулы |
дестабилизирует |
|
мембрану |
||||||||||||
фаллопиевой трубы достигает около |
акросомного пузырька, и это является |
||||||||||||||
200. Продолжительность времени, |
â |
необходимым |
условием |
слияния |
|||||||||||
течение которого овулировавшие яй- |
мембран при акросомной реакции. |
|
|||||||||||||
цеклетки способны оплодотвориться, |
Известно, |
÷òî |
|
важнейшее |
|||||||||||
различна у разных видов млекопитаю- |
проявление |
капацитации |
- |
||||||||||||
щих, но обычно не превышает 24 час. |
приобретение спермием способности |
||||||||||||||
Примерно за это же время спермии |
узнавать прозрачную оболочку яйца. |
||||||||||||||
утрачивают |
оплодотворяющую |
Íà |
поверхности |
спермия, |
íå |
||||||||||
способность, находясь в половом |
прошедшего капацитацию, находятся |
||||||||||||||
тракте самки. Это означает, что |
углеводы («coating factors»), которые |
||||||||||||||
оплодотворение |
происходит |
â |
препятствуют |
его прикреплению к |
|||||||||||
небольшой промежуток времени, |
ÿéöó. |
Эти углеводы представляют |
|||||||||||||
поэтому |
|
ó |
большинства |
собой, по-видимому, полимер из |
|||||||||||
млекопитающих спаривание (коитус) |
повторяющихся остатков галактозы и |
||||||||||||||
происходит непосредственно перед |
N-ацетилглюкозамина. Они блокируют |
||||||||||||||
овуляцией. |
|
|
|
|
|
|
|
рецепторы спермия, |
узнающие |
||||||
Ó |
|
|
млекопитающих |
прозрачную |
оболочку |
ÿéöà. |
|||||||||
оплодотворение внутреннее, и процесс |
Рецепторами |
спермия |
являются |
||||||||||||
оплодотворения |
приспособлен |
ê |
молекулы |
фермента |
|
N- |
|||||||||
внутренней среде организма, поэтому |
à ö å ò è ë ã ë þ ê î ç à ì è í |
||||||||||||||
половые пути самки принимают |
галактозилтрансферазы (или кратко |
||||||||||||||
активное |
участие в |
|
процессе |
- гликозилтрансферазы). Молекулы |
|||||||||||
оплодотворения. |
Сразу |
после |
гликозилтрансферазы встроены |
â |
|||||||||||
эякуляции спермии еще не способны к |
плазматическую мембрану |
спермия |
|||||||||||||
оплодотворению, так как вначале они |
непосредственно над акросомой, |
||||||||||||||
должны подвергнуться процессу, |
причем их активные центры обращены |
||||||||||||||
который присущ только мле- |
наружу. Гликозилтрансфераза узнает |
||||||||||||||
копитающим. |
Приобретение |
N-ацетилглюкозаминовые остатки |
|||||||||||||
спермиями |
|
млекопитающих |
белков |
прозрачной |
оболочки |
||||||||||
оплодотворяющей |
способности при |
(возможно, гликопротеина ZP3). При |
|||||||||||||
их контакте |
с клетками и жидким |
капацитации |
блокировавшие |
åå |
|||||||||||
содержимым |
яйцеводов |
è |
матки |
углеводы |
|
убираются, |
è |
||||||||
называется капацитацией. При |
гликозилтрансфераза на поверхности |
||||||||||||||
капацитации |
происходят изменения |
спермия обнажается. В норме этот |
|||||||||||||
структуры |
|
липидов |
клеточной |
фермент катализирует присоединение |
|||||||||||
мембраны спермия. Это выражается в |
активированных остатков галактозы к |
||||||||||||||
изменении соотношения холестерина и |
углеводной цепи, оканчивающейся N- |
||||||||||||||
фосфолипидов мембраны. Полагают, |
ацетилглюкозамином. |
Однако |
â |
||||||||||||
что молекулы альбумина, имеющиеся |
половых путях самки активированные |
||||||||||||||
в половых путях самки, |
способны |
остатки |
галактозы |
отсутствуют. |
|||||||||||
отнимать |
холестерин |
ó |
спермия. |
Поэтому гликозилтрансфераза спермия |
|||||||||||
___________________________________________________________________________________
3-28
Глава 3. Раннее развитие млекопитающих
______________________________________________________________________________
просто присоединяется к N- |
акросомного |
пузырька, который |
|||||||||||||||
ацетилглюкозаминовым остаткам |
происходит в результате кальций- |
||||||||||||||||
прозрачной оболочки яйца и таким |
зависимого |
слияния |
наружной |
||||||||||||||
образом, |
ïî |
|
òèïó |
ключ-замок |
акросомной мембраны и прилежащей |
||||||||||||
прикрепляет |
спермии |
ê |
ÿéöó. |
к ней плазматической мембраны |
|||||||||||||
Гликозилтрансфераза обнаружена в |
спермия в фронтальной половине |
||||||||||||||||
спермиях мыши, морской свинки и |
головки. |
Это по сути дела реакция |
|||||||||||||||
человека. |
|
|
|
|
|
|
|
экзоцитоза, заключающаяся в том, что |
|||||||||
Ó |
хорошо |
изученных |
пузырек приближается к клеточной |
||||||||||||||
млекопитающих - мыши и хомячка - |
поверхности |
|
è, |
сливаясь |
ñ |
||||||||||||
акросомная реакция |
происходит |
плазматической мембраной, выделяет |
|||||||||||||||
после |
прикрепления |
спермия |
ê |
свое содержимое наружу. Такие |
|||||||||||||
прозрачной оболочке. У некоторых |
реакции зкзоцитоза наблюдаются при |
||||||||||||||||
видов акросомная реакция происходит, |
выделении |
|
инсулина |
клетками |
|||||||||||||
когда спермий находится на некотором |
поджелудочной |
железы |
è |
ïðè |
|||||||||||||
расстоянии от яйца. В этих случаях |
выделении |
|
нейромедиаторов из |
||||||||||||||
акросомная |
реакция |
приводит |
ê |
синаптических окончаний. Во время |
|||||||||||||
выделению ферментов, лизирующих |
акросомной реакции плазматическая |
||||||||||||||||
связи |
между |
фолликулярными |
мембрана, |
|
|
|
покрывающая |
||||||||||
клетками, |
окружающими |
ÿéöî. |
постакросомальную область головки |
||||||||||||||
Ферменты, |
|
|
выделенные |
èç |
(ее экваториальный сегмент), |
приоб- |
|||||||||||
акросомного пузырька, |
называют |
ретает |
свойства, |
|
делающие |
||||||||||||
лизинами. |
|
Îíè |
включают |
возможным |
слияние этой |
части |
|||||||||||
гиалуронидазу, |
расщепляющую |
плазматической мембраны спермия с |
|||||||||||||||
гиалуроновую кислоту матрикса, и |
плазматической |
|
мембраной |
||||||||||||||
фермент, рассеивающий |
клетки |
яйцеклетки. |
Возможно, |
÷òî íà |
|||||||||||||
лучистого венца. Кроме того, спермии |
дестабилизацию мембраны спермия |
||||||||||||||||
всех млекопитающих содержат лизин, |
влияет изменение ее липидного |
ñîñ- |
|||||||||||||||
разрушающий прозрачную оболочку в |
тава в процессе капацитации. |
|
|
||||||||||||||
месте |
прикрепления. Этот лизин, |
Спермии быстро проходят через |
|||||||||||||||
называемый |
|
акрозином, |
ïî |
блестящую оболочку, |
и, например, у |
||||||||||||
аминокислотному составу напоминает |
хомячков это занимает лишь 5-10 мин |
||||||||||||||||
протеолитический фермент трипсин. |
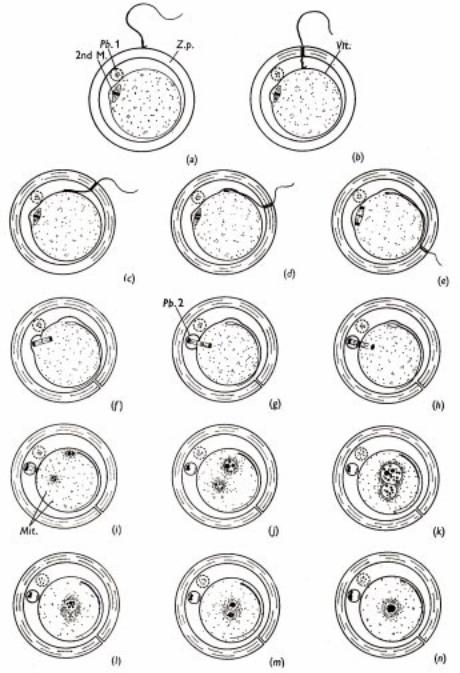
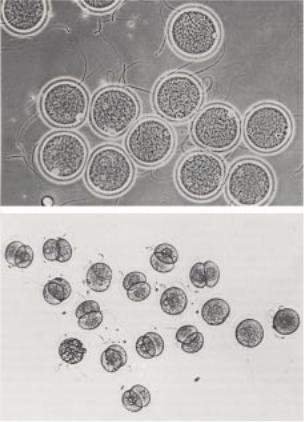
(ðèñ.3.1 è 3.2). |
Это прохождение |
|||||||||||||||
Полагают, что в спермиях находится |
осуществляется благодаря совместным |
||||||||||||||||
проакрозин, который активируется под |
действиям |
ферментов |
(вероятно |
||||||||||||||
воздействием |
|
гликопротеина |
èç |
акрозина), |
связанных с |
внутренней |
|||||||||||
половых путей самки. Таким образом, |
акросомной мембраной, и активных |
||||||||||||||||
как и в процессе капацитации, в |
движений хвоста спермия. Пройдя |
||||||||||||||||
инициации |
акросомной |
реакции |
через блестящую оболочку спермий |
||||||||||||||
половые пути самки играют активную |
попадает |
|
â |
|
перивителлиновое |
||||||||||||
ðîëü. |
|
|
|
|
|
|
|
|
пространство. |
|
Åñëè |
áëîê |
|||||
Морфологическим выражением |
полиспермии функционирует на |
||||||||||||||||
акросомной реакции является разрыв |
уровне блестящей оболочки, |
òî â |
|||||||||||||||
___________________________________________________________________________________
3-29
Глава 3. Раннее развитие млекопитающих
______________________________________________________________________________
перивителлиновое |
пространство |
спермия в цитоплазме яйца влияет на |
||||||||||||
попадает |
|
îäèí |
спермий. Если |
дальнейшую судьбу этого района. |
||||||||||
полиспермия блокируется на уровне |
Полагают, что бластомер, который |
|||||||||||||
плазматической мембраны яйцеклетки, |
получает этот участок цитоплазмы |
|||||||||||||
то в перивителлиновом пространстве |
яйца, быстрее вступает во второе де- |
|||||||||||||
можно |
найти |
много |
спермиев |
ление дробления, чем сестринский |
||||||||||
(например, |
ïðè |
оплодотворении |
бластомер (рис.3.1). |
|
|
|
|
|||||||
яйцеклеток кролика), хотя только один |
После |
проникновения |
â |
|||||||||||
из них сольется |
ñ |
яйцеклеткой. |
яйцеклетку |
головка спермия быстро |
||||||||||
Удаление |
блестящей |
оболочки |
теряет оболочки и хроматин начинает |
|||||||||||
приводит к исчезновению межвидовых |
деконденсироваться. |
|
Возможно |
|||||||||||
барьеров при оплодотворении, |
õîòÿ |
вследствие |
|
|
|
контакта |
||||||||
прикрепление спермия к прозрачной |
деконденсирующегося хроматина |
|||||||||||||
оболочке лишь относительно, а не аб- |
спермия с цитоплазмой яйца про- |
|||||||||||||
солютно |
видоспецифично |
(ïðè |
исходят |
локальные |
|
изменения |
||||||||
внутреннем оплодотворении видовая |
субкортикального слоя, |
в который |
||||||||||||
специфичность не является настолько |
попадает |
спермий. |
|
Молекулы |
||||||||||
важной проблемой, как при наружном |
глобулярного актина полимеризуются, |
|||||||||||||
оплодотворении). |
|
|
|
|
образуя скопление микрофиламентов. |
|||||||||
У млекопитающих в отличие от |
Плазматическая мембрана яйцеклетки |
|||||||||||||
других животных спермий сливается с |
выпячивается, |
образуя «бугорок» и |
||||||||||||
плазматической |
|
мембраной |
теряя микроворсинки, появляется так |
|||||||||||
яйцеклетки |
экваториальным |
называемый конус оплодотворения. |
||||||||||||
постакросомальным районом головки. |
Ýòîò |
участок |
очень |
напоминает |
||||||||||
Процесс слияния происходит при |
«лысый» |
район |
|
оолеммы, |
||||||||||
взаимодействии |
|
|
|
этого |
покрывающий субкортикальный слой |
|||||||||
специализированного |
района |
ÿéöà, |
в котором |
располагается |
||||||||||
мембраны спермия и микроворсинок |
мейотическое веретено метафазы II. |
|||||||||||||
плазматической мембраны яйца и |
Подчеркнем, что спермий пенетрирует |
|||||||||||||
занимает |
|
несколько |
минут. У |
яйцеклетку |
в любом районе, |
кроме |
||||||||
млекопитающих |
|
внутренняя |
«лысого», то есть спермий не попадает |
|||||||||||
акросомная мембрана, покрывающая |
в область |
ÿéöà, |
ãäå |
|
находится |
|||||||||
после акросомной реакции переднюю |
мейотическое |
веретено. Таким |
||||||||||||
часть головки спермия, по-видимому, |
образом, и хроматин головки спермия, |
|||||||||||||
обладает пониженной текучестью, |
и конденсированные мейотические |
|||||||||||||
поэтому она не сливается с оолеммой, |
хромосомы |
ÿéöà |
индуцируют |
|||||||||||
а как бы фагоцитируется яйцеклеткой. |
формирование в яйцеклетке районов |
|||||||||||||
У большинства млекопитающих |
цитоплазмы, |
|
|
|
богатых |
|||||||||
плазматическая мембрана хвоста |
микрофиламентами, |
è |
участков |
|||||||||||
сливается |
с оолеммой. |
È |
õîòÿ |
мембраны, лишенных микроворсинок. |
||||||||||
аксиальные филаменты и вносимая |
В норме только один спермий |
|||||||||||||
спермием |
|
|
центриоль |
сливается с яйцеклеткой и участвует в |
||||||||||
дезинтегрируются, |
наличие хвоста |
эмбриональном развитии (рис.3.1 и |
||||||||||||
___________________________________________________________________________________
3-30
Глава 3. Раннее развитие млекопитающих
______________________________________________________________________________
Рис.3.1. Схема оплодотворения яйцеклетки крысы. Хромосомные наборы становятся постепенно видимыми в районе пронуклеусов. Хромосомы видны как две группы и находятся на стадии профазы первого митоза, что означает окончание оплодотворения. Pb.1 - первое полярное тельце; 2ndM. - метафаза 2-ого мейотического деления. Vit - вителлиновая оболочка; Z.p. - zona pellucida; Pb.2 - второе полярное тельце.
3.2). Полиспермные зародыши по- |
мембраны яйца в течении десятой доли |
||||
гибают. В |
процессе эволюции |
секунды после прикрепления к ней |
|||
животные выработали разные способы |
первого спермия, и медленный блок |
||||
защиты от полиспермии. |
У таких |
полиспермии, |
вызываемый |
||
животных, |
как морские ежи и |
экзоцитозом кортикальных гранул. |
|||
амфибии, имеется два механизма: |
У млекопитающих |
ëèøü |
|||
быстрый |
áëîê, |
который |
небольшое |
число |
спермиев |
осуществляется путем деполяризации |
оказывается в состоянии достигнуть до |
||||
___________________________________________________________________________________
3-31
Глава 3. Раннее развитие млекопитающих
______________________________________________________________________________
претерпевшей овуляцию яйцеклетки, |
различать первичные, вторичные и |
|||||||||||||
поэтому необходимость в быстрой |
третичные эффекты активации. |
|
||||||||||||
блокаде полиспермии несущественна: |
|
Первичный эффект: вытеснение |
||||||||||||
у млекопитающих не происходит |
Ñà2+ из плазматической мембраны яй- |
|||||||||||||
быстрой |
|
деполяризации |
цеклетки. |
|
|
|
|
|
||||||
плазматической мембраны. С другой |
|
Вторичный эффект: резкое и |
||||||||||||
стороны, |
|
|
|
ферменты, |
кратковременное |
повышение |
||||||||
высвобождающиеся при кортикальной |
концентрации Са2+ в цитозоле, которое |
|||||||||||||
реакции млекопитающих, изменяют |
распространяется от места внедрения |
|||||||||||||
структуру |
прозрачной |
оболочки, |
спермия по всей яйцеклетке в виде |
|||||||||||
обеспечивая медленную блокаду |
кольцевой волны. Одновременно |
|||||||||||||
полиспермии. |
Ýòîò |
процесс |
происходит экзоцитоз все новых |
|||||||||||
сопровождается изменением свойств |
кортикальных гранул. Вслед за пиком |
|||||||||||||
рецепторов прозрачной оболочки, |
ñ |
концентрации ионов кальция более |
||||||||||||
которыми связываются спермии, и так |
медленно, |
но неуклонно повышается |
||||||||||||
и называется - реакция прозрачной |
ðÍ. |
|
|
|
|
|
|
|||||||
оболочки. |
|
Полагают, |
÷òî |
|
Третичный |
|
эффект: |
|||||||
кортикальные гранулы в яйцах мыши |
инактивация |
|
белка |
|
CSF |
|||||||||
содержат фермент, который отщепляет |
(цитостатического фактора), который, |
|||||||||||||
концевые остатки сахара на ZРЗ, тем |
êàê |
полагают, |
ответственнен за |
|||||||||||
самым отделяя |
прикрепившиеся |
ê |
задержку ооцитов на стадии метафаза |
|||||||||||
прозрачной оболочке спермий |
è |
II |
мейоза и поддерживает такое |
|||||||||||
предотвращая |
прикрепление |
ê íåé |
состояние ооцитов до активации. |
|
||||||||||
новых спермиев. |
|
|
|
|
3.1.12.Молекулярные механизмы |
|||||||||
Зрелая яйцеклетка является ме- |
||||||||||||||
таболически инертной. Спермий |
активации яйцеклетки. |
|
|
|||||||||||
запускает |
программу |
развития, |
|
Спермий играет роль сигнального |
||||||||||
заложенную в яйце. Связывание |
|
|||||||||||||
лиганда, который связывается с белко- |
||||||||||||||
спермия с поверхностью яйцеклетки |
||||||||||||||
индуцирует |
повышение |
|
åå |
вым рецептором плазматической |
||||||||||
метаболической активности, |
синтез |
мембраны, запуская следующую цепь |
||||||||||||
ДНК и последующее дробление. |
событий. Активированный рецептор |
|||||||||||||
Однако сам спермий служит лишь |
активирует |
G-белок |
(GÒÐ- |
|||||||||||
устройством для запуска уже |
çàëî- |
связывающий мембранный белок, |
||||||||||||
женной в яйцеклетке программы. |
который опосредует передачу в клетку |
|||||||||||||
Яйцеклетку можно активировать и |
внеклеточного сигнала). G-белок |
|||||||||||||
искусственно, с помощью множества |
активирует фермент фосфоинозитид- |
|||||||||||||
неспецифических |
физических |
è |
специфическую фосфолипазу С. В |
|||||||||||
химических воздействий. Например, |
мембране |
клетки |
содержатся |
|||||||||||
äëÿ ÿéöà |
лягушки |
эффективным |
инозитолфосфолипиды, они являются |
|||||||||||
стимулом может быть укол иглой, для |
субстратами |
фосфолипазы |
Ñ. |
|||||||||||
яйца мыши присутствие в среде 7% |
Инозитолфосфолипиды состоят из |
|||||||||||||
этанола. У млекопитающих принято |
сахарофосфатного остатка, который |
|||||||||||||
___________________________________________________________________________________
3-32
Глава 3. Раннее развитие млекопитающих
______________________________________________________________________________
à
á
Рис.3.2. Результат оплодотворения in vitro яйцеклетки китайского хомячка. А – группа из 12-и яйцеклеток инкубируется в капле среды, в которую добавлены сперматозоиды из эпидидимуса. 11 яйцеклеток находятся на разных стадиях оплодотворения. Рядом с ними находятся хвосты сперматозоидов, все 11 яиц имеют два пронуклеуса и два полярных тельца. Б – группа яиц через 24 часа после оплодотворения in vivo. 11 из 17-и яиц были оплодотворены и претерпели одно деление дробления. Видны ядра в каждом бластомере. Одно яйцо фрагментировано.
находится на внутренней стороне плазматической мембраны и двух цепей жирных кислот, пронизывающих липидный монослой плазматической мембраны. Показано, что гидролиз инозитолфосфолипидов участвует в передаче сигнала более чем от 25 различных рецепторов клеточной поверхности. Активированная фосфолипаза С менее, чем за секунду г и д р о л и з у е т фосфатидилинозитолбисфосфат
(коротко PIP2) на диацилглицерол и инозитолтрисфосфат (коротко Ins P3). Ins P3 (сахарофосфатный остаток) инициирует высвобождение Са2+ из внутриклеточных хранилищ в цнтозоль; диацилглицерол (остаток жирных кислот) активирует протеинкиназу С.
Концентрация свободных ионов кальция в цитозоли клеток очень низка (порядка 10-7 М), тогда как концентрация их во внеклеточной жидкости (более 10-3 М). Ионы кальция хранятся в специальных Сазапасающих органеллах в ассоциации с Са2+-связывающим белком кальсеквестрином (его емкость около 50 ионов Са2+ на молекулу). 1nз P3 связывается с рецептором на цитоплазматической поверхности Сазапасающей органеллы, что открывает кальциевые каналы в ее мембране. Диффузия Ins P3 приводит к распространению выхода кальция по всему кортикальному слою яйца. Выход кальция приводит к экзоцитозу кортикальных гранул, что обеспечивает блок полиспермии. Другим кальций-зависимым эффектом является активация HAД+-киназы, что приводит к стимуляции биосинтеза липидов. В конечном итоге создаются новые компоненты плазматической мембраны, необходимые в процессе дробления.
Поскольку концентрация ионов Са2+ в цитозоли после оплодотворения повышается лишь ненадолго (примерно на 1 мин.), ясно, что это не может непосредственно приводить к событиям, происходящим на более поздних стадиях активации яйцеклетки. На поздних стадиях
___________________________________________________________________________________
3-33
Глава 3. Раннее развитие млекопитающих
______________________________________________________________________________
активации |
важную |
ðîëü |
играет |
располагается в |
тангенциальном |
|||||
протеинкиназа С, которая активируется |
положении недалеко от краевой зоны |
|||||||||
диацилглицеролом. |
|
|
|
|
|
яйцеклетки. После активации, |
когда |
|||
Протеинкиназа С активирует |
мейоз возобновляется и |
метафаза |
||||||||
(преимущественно |
посредством |
переходит в анафазу (т.е. начинается |
||||||||
фосфорилирования) |
|
|
Nà+-Í+- |
расхождение хроматид и их движение |
||||||
ионообменник |
плазматической |
к полюсам) начинает формироваться |
||||||||
мембраны |
яйцеклетки (мембранный |
контрактильное |
кольцо. |
Åãî |
||||||
транспортный белок, откачивающий |
формирование происходит в том месте, |
|||||||||
èîíû Í+ из клетки). Отток протонов |
где находится мейотическая пластинка. |
|||||||||
приводит к тому, |
что величина рН |
Для образования борозды деления |
||||||||
внутри клетки возрастает |
è ïðè |
оказывается |
доступной |
ëèøü |
||||||
дальнейшем развитии |
|
зиготы |
небольшая область плазматической |
|||||||
поддерживается на этом уровне. |
|
мембраны. В результате происходит |
||||||||
Повышение |
уровня |
ðÍ |
â |
асимметричная цитотомия, то есть |
||||||
яйцеклетке приводит |
ê |
усилению |
отделяется второе полярное тельце, в |
|||||||
белкового синтеза и |
синтеза ДНК. |
состав которого попадает около 1% |
||||||||
Синтез |
белков |
усиливается |
â |
цитоплазмы яйца (рис.3.1). Аналогич- |
||||||
результате, |
по меньшей мере, двух |
ным образом происходит и выделение |
||||||||
независимых изменений: |
1) ранее |
первого полярного тельца. |
|
|
||||||
запасенные в яйце молекулы мРНК |
Выделение второго |
полярного |
||||||||
становятся доступными для синтеза |
тельца завершает второе мейотическое |
|||||||||
белков; 2) происходит активация |
деление. |
|
|
|
||||||
рибосом, что позволяет им быстрее |
|
|
|
|
||||||
транслировать мРНК. |
|
|
|
|
|
|
|
|
||
3.1.12.1. Завершение мейоза.
Расхождение хромосом из метафазы II является автоматическим процессом и осуществляется вне зависимости от присутствия генетически инертного хроматина спермия (рис.3.1). Нормальное расхождение хромосом ооцита происходит и при искусственной активации яйцеклетки к партеногенетическому развитию, Мейотическое веретено метафазы II имеет такое же строение, как и мейотическое веретено метафазы I. Это веретено лишено центриоль, обладает бочкообразной формой и
___________________________________________________________________________________
3-34