

22. Возбудимость мембраны различных частей нейрона
В нейроне подавляющее большинство нейрональных синапсов находится на дендритах нейрона. Однако наиболее эффективно вызывают возбуждение нейрона синаптические контакты, расположенные на теле нейрона. Это связано с тем, что постсинаптические мембраны этих синапсов располагаются в непосредственной близости от места первичного возникновения ПД, располагающегося в аксонном холмике (место отхождения аксона от тела). Близость соматических синапсов к аксонному холмику обеспечивает участие их возбуждающего постсинаптического потенциала (ВПСП) в механизмах генерации ПД (некоторые авторы называют их генераторными синапсами). Также большой возбудимостью обладает мембрана в местах первого ветвления крупных дендритов. Чем ближе конкретный синапс к этим точкам, тем больше его вклад в управление генерацией ПД. Одного ТПСП, возникшего рядом с аксональным холмиком, может оказаться достаточно для прекращения проведения сигнала.
Генераторный пункт нейрона, т. е. место возникновения ПД - аксонный холмик (но ПД также может возникать в перехватах Ранвье, и на части мембраны дендритов). Синапсы на нем отсутствуют, отличительной особенностью мембраны аксонного холмика является ее высокая возбудимость, в 3 — 4 раза превосходящая возбудимость сома-дендритной мембраны нейрона, что объясняется более высокой концентрацией Na-каналов на аксонном холмике, отсутствием на нем миелиновой оболочки. ВПСП достигает аксонного холмика, обеспечивая здесь уменьшение мембранного потенциала до критического уровня. В этот момент в аксонном холмике возникает ПД.
Роль дендритов в возникновении возбуждения до сих пор дискутируется. Дендритные синапсы удалены на значительное расстояние от генераторного пункта нейрона, по этой причине их ВПСП не могут вызвать там должную деполяризацию и обеспечить генерацию ПД. Считают, что синаптический аппарат дендритов проявляет себя при одновременном поступлении возбуждения к значительному числу дендритных синапсов. При этом суммарный дендритный ВПСП, изменяя мембранный потенциал генераторного пункта на подпороговом уровне, лишь делает возбудимость большей или меньшей в зависимости от временных и амплитудных характеристик колебаний мембранного потенциала генераторного пункта относительно величины критического уровня деполяризации.
23. Возможные механизмы генерации потенциала действия нейрона в естественных условиях
Потенциал действия - это электрофизиологический процесс, выражающийся в быстром колебании мембранного потенциала покоя вследствие перемещения ионов в клетку и из клетки и способный распространяться без затухания. ПД обеспечивает передачу сигналов между нервными клетками, между нервными центрами и рабочими органами. ПД подчиняется закону «все или ничего», но не подчиняется закону силовых отношений, т. е. закону силы. При малом раздражении клетки ПД либо совсем не возникает, либо достигает максимальной величины, если раздражение является пороговым или сверхпороговым. Слабое (подпороговое) раздражение может вызвать локальный потенциал, который подчиняется закону силы: с увеличением силы стимула величина его также возрастает.
Запуск импульсной активности в нервной системе осуществляют два основных фактора. Первый из них — стимулы, действующие на чувствительные клетки сенсорных систем и изменяющие проницаемость их мембраны. Это приводит к развитию особых рецепторных потенциалов и в итоге — к генерации ПД.
Второй фактор — выделение медиатора из пресинаптического окончания. Попав в синаптическую щель, медиатор воздействует на постсинаптическую мембрану, возбуждая или тормозя следующий нейрон.
Механизм возникновения ПД. Если действие раздражителя на клеточную мембрану приводит к началу развития ПД, далее сам процесс развития ПД вызывает фазовые изменения проницаемости клеточной мембраны, что обеспечивает быстрое движение Na+ в клетку, а К+ — из клетки. Это наиболее часто встречаемый вариант возникновения ПД. Величина мембранного потенциала при этом сначала уменьшается до нуля, изменяет знак заряда, а затем снова восстанавливается до исходного уровня. Эти изменения мембранного потенциала предстают в виде пикового потенциала - ПД. Если заблокировать процесс выработки энергии, ПД некоторое время будет возникать. Но после исчезновения градиентов концентраций ионов (устранения потенциальной энергии) клетка генерировать ПД не будет. ПД проходит следующие фазы: 1). фаза деполяризации — процесс исчезновения заряда клетки до нуля; 2) фаза инверсии - изменение заряда клетки на противоположный, т.е. весь период ПД, когда внутри клетки заряд положительный, а снаружи отрицательный; 3) фаза реполяризации — восстановление заряда клетки до исходной величины (возврат к потенциалу покоя). Главную роль в возникновении ПД играет Na+, входящий в клетку при повышении проницаемости клеточной мембраны и обеспечивающий всю восходящую часть пика ПД. Однако проницаемость мембраны для К+ тоже играет важную роль. Если повышение проницаемости для К+ предотвратить, то мембрана после ее деполяризации реполяризуется гораздо медленнее, только за счет медленных неуправляемых каналов (каналов утечки ионов), через которые К+ будет выходить из клетки.
Для достижения порога запуска ПД необходима суммация (наложение) нескольких ВПСП. Выделяют два варианта суммации — временную и пространственную. Временная суммация — объединение эффектов стимулов, пришедших по одному «каналу» с большой частотой: если к еще не угасшему ВПСП присоединить второй, затем третий и т.д., — возникнет реальная возможность запустить ПД.
Пространственная суммация заключается в наложении друг на друга ВПСП соседних синапсов в некоторой близлежащей точке постсинаптической мембраны.
В ходе деятельности нейронов эффекты пространственной и временной суммации объединяются, и чем больше синапсов участвуют в этом процессе (срабатывают относительно одномоментно), тем больше вероятность достичь порога запуска ПД. При этом часть синапсов может обладать тормозными свойствами и вызывать ТПСП, вычитающиеся из суммы возбуждающих влияний. В результате условие запуска ПД в каждый момент времени можно определить следующим образом:
ПП + (сумма всех ВПСП) − (сумма всех ТПСП) > порог запуска ПД
И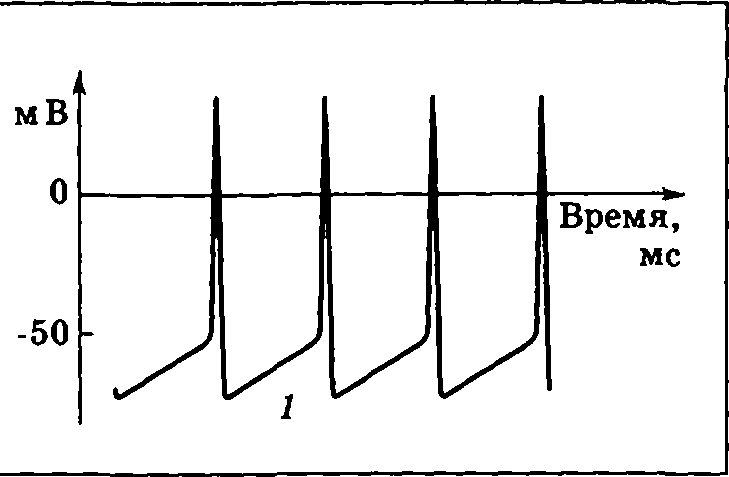 нтересным
вариантом генерации ПД являютсяпейсмекерные нейроны
(клетки-пейсмекеры).
Они обладают большой постоянной
проницаемостью мембраны для ионов Na+.
В результате у клеток-пейсмекеров не
существует стабильного ПП. Разность
потенциалов на их мембране постоянно
стремится вверх. Когда она достигает
порогового значения, происходит запуск
ПД. После ПД заряд внутри клетки
оказывается на довольно низком уровне,
ПП вновь растет и запускается следующий
ПД, в целом же наблюдается ритмический
рисунок разрядов (рис. 3.12). Рост заряда
1 внутри
клетки, связанный с большим током утечки
Na+,
приводит к периодической самопроизвольной
генерации потенциалов действия.
Нейроны-пейсмекеры находятся в дыхательном
центре продолговатого мозга, сходные
же свойства имеют клетки центра сердечного
автоматизма.
нтересным
вариантом генерации ПД являютсяпейсмекерные нейроны
(клетки-пейсмекеры).
Они обладают большой постоянной
проницаемостью мембраны для ионов Na+.
В результате у клеток-пейсмекеров не
существует стабильного ПП. Разность
потенциалов на их мембране постоянно
стремится вверх. Когда она достигает
порогового значения, происходит запуск
ПД. После ПД заряд внутри клетки
оказывается на довольно низком уровне,
ПП вновь растет и запускается следующий
ПД, в целом же наблюдается ритмический
рисунок разрядов (рис. 3.12). Рост заряда
1 внутри
клетки, связанный с большим током утечки
Na+,
приводит к периодической самопроизвольной
генерации потенциалов действия.
Нейроны-пейсмекеры находятся в дыхательном
центре продолговатого мозга, сходные
же свойства имеют клетки центра сердечного
автоматизма.
Рис. 3.12. Изменение разности потенциалов на мембране пейсмекерного нейрона
Принципиальна новое, что вносит пейсмекерный потенциал в функционирование нейрона, заключается в следующем: пейсмекерный потенциал превращает нейрон из сумматора синаптических потенциалов в генератор.
При изучении нейронов оказалось что даже если нейрон «молчит», на постсинаптической мембране все равно наблюдаются периодические изменения мембранного потенциала - редкие, очень низкоамплитудные, кратковременные случаи деполяризации. Эти потенциалы были названы миниатюрными потенциалами (МП).
МП возникают в ответ на случаи спонтанного выброса медиатора из пресинапса в синаптическую щель. Как правило, свое содержимое в этом случае выбрасывают буквально единичные везикулы, поэтому МП отражают действие на постсинаптическую мембрану одного кванта медиатора - минимально возможной порции медиатора, которая может быть выброшена в щель, то есть содержимого одной везикулы.