

Препарат 2. Поздняя гаструла амфибий (сагиттальный срез)
На стадии поздней гаструлы образуется вентральная губа бластопора, так как пигментированные клетки анимального полюса на этом этапе достигают вегетативного полюса и обрастают его, достигая латеральных губ бластопора. Через вентральную губу также происходит подворачивание клеточного материала, но незначительно. Бластопор становится широкоокруглым. Через него просвечивают макромеры, которые на этом этапе формируют желточную пробку. Несколько позднее за счет сокращений краев бластопора происходит втягивание желточной пробки внутрь. Гастроцель крупна, полностью вытесняет бластоцель и смещена к анимальному полюсу. Крыша гастроцели содержит в себе несколько зачатков и энтодерма на этом этапе называетсяпервичной кишкойилиархентероном.В ее состав входят: зачаток хордомезодермы, мезодерма сомитов и энтодерма. Энтодерма у амфибий представлена кишечной, идущей на выстилку средней кишки, и желточной, расходующейся на питание зародыша.
Препарат 3. Ранняя нейрула амфибий (поперечный срез, рис. 18, а)
Гаструла постепенно переходит в нейрулу, когда начинается образование нервной трубки и ганглиозной пластинки из нервных (медуллярных) валиков. Параллельно с этим процессом начинается дифференцировка мезодермы. Сегментированные участки ее, лежащие по бокам от хорды, на дорзальной стороне, образуют сомиты. Несегментированные участки, располагающиеся вентрально и обрастающие по бокам кишку, называются спланхнотомы. Нейруляция заканчивается формированием трехслойного зародыша.
Препарат 4. Поздняя нейрула амфибий (поперечный срез, рис. 18, в)
Поздняя нейрула или трехслойный зародыш амфибий характеризуется хорошо видимой закладкой комплекса осевых органов: нервной трубки, ганглиозной пластинки, хорды, кишечной энтодермы (позже – трубки) и дифференцировкой мезодермы на сомиты и спланхнотомы. Позднее в спланхнотомах образуется полость (целом).
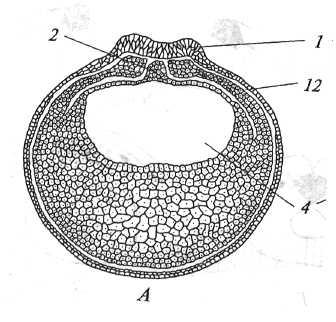
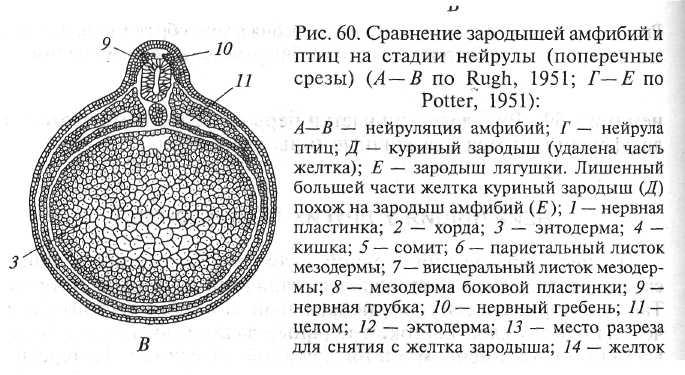
Рис. 18. Поперечные срезы нейрулы амфибий (Голиченков, 2004). А — ранняя; В — поздняя. 1 — нервная пластинка; 2 — хорда; 3 — энтодерма; 4 — полость кишки; 9 — нервная трубка; 10 — нервный гребень; 11 — целом; 12 — эктодерма.
Препарат 5. Гаструла морского ежа (поперечный срез)
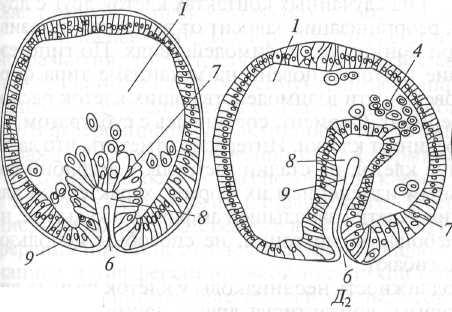
На рис. 19 видно, что гаструляция у морского ежа протекает по типу инвагинации с элементами униполярной иммиграции. Бластомеры вегетативного полюса целобластулы несколько крупнее, чем на анимальном полюсе. Часть клеток вегетативного полюса иммигрируют в бластоцель и располагаются там рыхло, формируя первичную мезенхиму. Оставшиеся клетки вегетативного полюса становятся высокими, цилиндрическими и клеточный пласт начинает инвагинировать в бластоцель.
Занятие 6 Развитие хрящевых ганоидов (осетровых рыб)
Хрящевым ганоидам свойственен тот же примитивный тип развития, какой характерен для амфибий, а также круглоротых, костных ганоидов, кистеперых и двоякодышащих рыб. У осетровых рыб умеренно телолецитальные яйца, полное неравномерное дробление, непрямое развитие с личиночной стадией и метаморфозом. Однако, яйца хрящевых ганоидов более дифференцированы в отношении разделения желточного запаса и образовательной цитоплазмы.
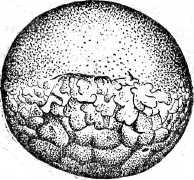
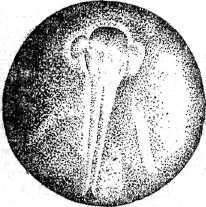
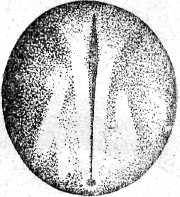
Неравномерность в дроблении проявляется в том, что некоторые борозды лишь врезаются в яйцо, не достигая вегетативного полюса, а другие делят яйцо полностью. В результате формируется амфибластула. Мелкие анимальные бластомеры занимают сначала 1/3 поверхности яйца. Гаструляция похожа на таковую амфибий и проявляется в инвагинации и эпиболии. При инвагинации мелкие клетки анимального полушария проникают внутрь через дорзальную и латеральные губы бластопора и формируют крышу архентерона. Здесь располагаются зачатки хорды и мезодермы. Клетки, проходящие внутрь через вентральную губу бластопора, образуют дно архентерона с энтодермальной подушкой. Эпиболия проявляется в наползании микромеров на вегетативные макромеры. Когда микромеры покрывают еще только половину поверхности яйца, инвагинация уже заканчивается, и эти микромеры, лежащие у дорзальной губы бластопора, являются материалом для образования нервной пластинки. Вслед за окончанием гаструляции этот материал образует два сближенных между собой медуллярных валика. К концу обрастания желточной пробки, передний конец медуллярных валиков раздвигается, образуя передний и задний мозговые пузыри. В то же время нервные валики в задней части удлиняются и задним краем нарастают на желточную пробку. К этому моменту начинается сегментация сомитов, в передних из них начинают формироваться пронефросы. После закрытия желточной пробки формирование туловищных сомитов завершается, а вольфовы протоки достигают полной длины. В головной части формируются мозговые пузыри, которые значительно выступают над поверхностью яйца, две пары жаберных щелей (их число затем возрастает до пяти пар), слуховые пузырьки. На заднем конце зародыша в виде бугорка появляется хвостовая почка. Все эти образования распластаны на сферической поверхности яйца.
Затем, по мере роста и дифференцировки сомитов, головной отдел зародыша и передние 20 сегментов туловища приподнимаются над поверхностью яйца (желтка). Хвостовая почка увеличивается в длину. На поверхности желтка под передним концом головы образуется сердце. По мере потребления желтка энтодермальной подушки его размеры уменьшаются только в дорзовентральном направлении. Снизу головного отдела и в области 40-го сомита, путем образования наружных выемок и полостей возникают ротовое и анальное отверстия. Происхождение последнего никак не связано с бластопором, который замкнулся на уровне 20-го сомита.
В образовании хвостовой почки принимают участие продолжения зачатков всех осевых органов, собирающихся в области бластопора, превратившегося при замыкании нервной трубки в нервно-кишечный канал. При продолжении роста хвостовой почки и достижения зародышем предельной длины, свойственной данному виду, рост зародыша в длину прекращается и нервно-кишечный канал зарастает.
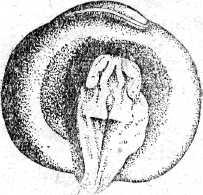
Из яйца осетровых рыб развивается личинка, которая имеет внутренние жабры, торчащие из-под жаберной крышки, короткую голову, обонятельные ямки и недоразвитые глаза. По краям рта возникают мелкие острые зубы, сходные по строению с зубами акул. Есть зачатки наружных жабр и брызгальца. Тело личинки сверху и снизу до конца желточного мешка окаймлено плавательной эктодермальной складкой. Личинка имеет зачатки грудных плавников в виде небольших полукруглых складок.
Метаморфоз протекает быстро. Образуются жаберные крышки, вырастает рострум. Размеры желточного мешка значительно сокращаются. Формируются спинные, грудные, брюшные и хвостовой плавники. Тело малька покрывается чешуёй (жучками) и появляются накладные кости черепа. Как и у амфибий, метаморфоз у хрящевых ганоидов связан с активностью щитовидной железы.

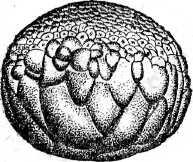

Д. Е.
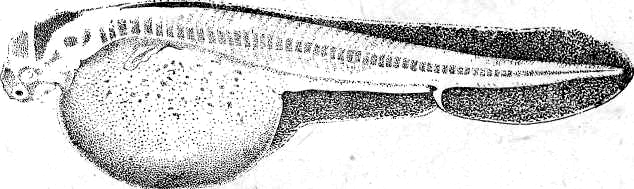
З.
Рис. 20. Развитие хрящевых ганоидов (Иванов, 1945). А начало дробления; Б бластула; В начало гаструляции; Г конец гаструляции; Д нейрула; Е обрастание желтка зародышем; Ж отделение зародыша от желтка; З личинка.