

Biologia_V_N_Yarygin_Tom_1_2008
.pdfРис. 3.67. Кариотипы организмов различных видов: I — скерды, II — дрозофилы. III — человека
Кариотип — диплоидный набор хромосом, свойственный соматическим клеткам организмов данного вида, являющийся видоспецифическим признаком и характеризующийся определенным числом, строением и генетическим составом хромосом (рис. 3.67). Ниже приведены количества хромосом соматических клеток некоторых видов организмов.
Если число хромосом в гаплоидном наборе половых клеток обозначить п, то общая формула кариотипа будет выглядеть как 2п, где значение п различно у разных видов. Являясь видовой характеристикой организмов, кариотип может отличаться у отдельных особей некоторыми частными особенностями. Например, у представителей разного пола, имеются в основном одинаковые пары хромосом (аутосомы), но их кариотипы отличаются по одной паре хромосом (гетерохромосомы, или половые хромосомы). Иногда эти различия состоят в разном количестве гетерохромосом у самок и самцов (XX или ХО). Чаще различия
151
касаются строения половых хромосом, обозначаемых разными буквами —X и Y (XX или XY).
Каждый вид хромосом в кариотипе, содержащий определенный комплекс генов, представлен двумя гомологами, унаследованными от родителей с их половыми клетками. Двойной набор генов, заключенный в кариотипе,— генотип — это уникальное сочетание парных аллелей генома. В генотипе содержится программа развития конкретной особи.
Животные |
Число хромосом |
Малярийный плазмодий |
2 |
Гидра |
32 |
Таракан |
48 |
Комнатная муха |
12 |
Сазан |
104 |
Окунь |
28 |
Зеленая лягушка |
26 |
Голубь |
80 |
Кролик |
44 |
Шимпанзе |
48 |
Человек |
46 |
3.6.2. Проявление свойств наследственного материала на геномном уровне его организации
3.6.2.1. Самовоспроизведение и поддержание постоянства кариотипа в ряду поколений клеток
В основе самовоспроизведения кариотипа в ряду клеточных поколений лежат процессы самоудвоения отдельных хромосом, описанные в разд. 3.5.3.1. Жизнедеятельность клеток как структурно-функциональных единиц живого обеспечивается всей совокупностью получаемого ими генетического материала, поэтому особое значение приобретает поддержание постоянства кариотипа в ряду их поколений. На геномном уровне организации наследственного материала это обеспечивается всей совокупностью процессов, происходящих в митотическом цикле (см. гл. 2).
152
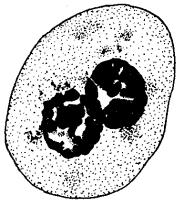
Рис. 3.68. Образование двуядерных клеток в результате торможения цитотомии при их делении
Если содержание ДНК в гаплоидном наборе хромосом, т.е. в геноме, является видоспецифическим и соответствует с, то в диплоидном кариотипе, равном 2n, оно составляет 2с. В ходе митотического цикла количество ДНК в клетке изменяется от 2с (сразу после ее образования, когда хромосомы состоят из одной нити) до 4с (после репликации ДНК в S-периоде интерфазы, когда хромосомы становятся двунитчатыми). Число хромосом при этом остается неизменным и равным 2n на протяжении всего митотического цикла. Благодаря описанным ранее процессам, происходящим в ходе интерфазы и последующего митоза (см. гл. 2), дочерние клетки получают от материнской полноценную наследственную программу — генотип, заключенный в кариотипе.
Однако не всегда клетки, удвоившие свой наследственный материал, вступают в митоз. Этим обусловлено существование в тканях некоторого количества клеток с формулой наследственного материала 2я4с, представляющих определенный резерв, готовый немедленно приступить к делению. Наконец, не всегда начавшееся деление заканчивается образованием двух дочерних клеток. Нарушение деления цитоплазмы при нормальном формировании дочерних ядер приводит к образованию двуядерных клеток (рис. 3.68). Разрушение ахроматанового веретена, делающее невозможным распределение дочерних хромосом между его полюсами, сопровождается появлением полиплоидных клеток (рис. 3.69), в которых 4п дочерних хромосом остаются в одном неразделившемся ядре.
Указанные отклонения от обычного течения митотического цикла являются причиной появления в тканях клеток, несущих больший по объему наследственный материал.
153
Рис. 3.69. Образование полиплоидньк клеток в результате разрушения веретена деления. А — различные наборы хромосом (12, 24, 48) в пыльцевых зернах одного из сортов гиацинта; Б — образование полиплоидных клеток
/—в норме, II—при разрушении веретена деления колхицином
3.6.2.2.Механизмы поддержания постоянства кариотипа
вряду поколений организмов
Уорганизмов, размножающихся бесполым путем, новое поколение появляется из неспециализированных в отношении генеративной функции клеток тела. В основе их самовоспроизведения лежит митоз, обеспечивающий таким образом сохранение постоянной структуры наследственного материала в ряду поколений не только клеток, но и организмов.
154

Рис. 3.70. Сравнение первого мейотического (редукционного) деления с митозом
Профаза — спирализация хромосом, начало формирования веретена деления; в мейозе, кроме того, происходит конъюгация гомологичных хромосом с образованием бивалентов;
метафаза — в митозе в экваториальной плоскости веретена деления располагаются отдельные хромосомы числом 2n, в мейоэе в плоскости экватора выстраивается п бивалеитов;
анафаза —в митозе в результате расщепления центромер дочерние хромосомы (бывшие сестринские хроматиды) расходятся к разным полюсам (по 2n к каждому полюсу), в мейозе разрушаются бивалеиты и гомологи расходятся к разным полюсам (по одному из каждой лары); формируется гаплоидный набор хромосом;
телофаза—в митозе формируются ядра дочерних клеток, в мейозе телофаза сокращена во времени, так как не происходит полной деспирализации хромосом и клетки сразу переходят ко второму делению.
Результаты митоза — сохранение в дочерних клетках диплоидного набора хромосом (2n2с); результаты первого мейотического деления—образование клеток с гаплоидным набором двунитчатых хромосом (п2с)
При половом размножении процесс воспроизведения организмов осуществляется с участием специализированных половых клеток — гамет, вступающих в оплодотворение. При оплодотворении наследственный материал двух родительских гамет сливается, образуя генотип организма нового поколения — зиготы. Чтобы потомки получили соответствующую программу для развития видовых и индивидуальных характеристик, они должны обладать кариотипом, которым располагало предыдущее поколение. В такой ситуации поддержание постоянства кариотипа в ряду поколений организмов достигается предварительным
155
уменьшением вдвое набора хромосом в гаметах, который восстанавливается до диплоидного при их оплодотворении: п + п = 2n.
Образование гаплоидных гамет осуществляется в ходе гаметогенеза путем особой формы клеточного деления — мейоза. При мейозе из клеток с диплоидным набором In образуются гаметы с гаплоидным набором хромосом п (см. гл. 5). Такой результат достигается благодаря тому, что после однократного удвоения ДНК клетка делится дважды. В отличие от митоза в первом мейотическом делении в результате конъюгации гомологичные хромосомы объединяются в пары — биваленты. Последующее расхождение гомологов к разным полюсам веретена деления приводит к образованию клеток с гаплоидным набором хромосом: 2n4с → п2с. На рис. 3.70 представлены особенности первого деления мейоза в сравнении с митозом. В ходе второго мейотического деления сестринские хроматиды каждой хромосомы, как и в митозе, распределяются между дочерними клетками с наследственным материалом пс (рис. 3.71).
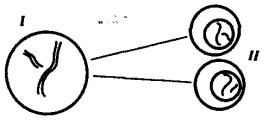
Рис. 3.71. Схема второго (эквационного) деления мейоза:
I — клетка, образованная в результате первого мейотического деления и несущая гаплоидный набор двунитчатых хромосом (n2c);
II — клетки, образующиеся после второго деления мейоза и несущие гаплоидный набор однонитчатых хромосом (nc)
Благодаря особенностям мейоза образуются клетки, несущие полноценный геном, в котором каждая группа сцепления представлена в единственном экземпляре (гаплоидный набор хромосом).
При самооплодотворении гаметы одного и того же родителя, а при перекрестном оплодотворении половые клетки разных организмов взаимодействуют друг с другом. Сперматозоиды, проникая в яйцеклетку, вводят в нее свой ядерный наследственный материал, заключенный в гаплоидном наборе хромосом. Ядра гамет сливаются и формируют диплоидное ядро зиготы, в котором каждая группа сцепления представлена в двойном экземпляре — отцовской и материнской хромосомами.
Таким образом, мейоз и последующее оплодотворение обеспечивают сохранение у нового поколения организмов диплоидного кариотипа, присущего всем особям данного вида.
156
3.6.2.3. Рекомбинация наследственного материала в генотипе. Комбинативная изменчивость
Мейоз и оплодотворение обеспечивают получение организмами нового поколения эволюционно сложившегося, сбалансированного по дозам генов наследственного материала, на основе которого осуществляется развитие организма и отдельных его клеток. Благодаря этим двум механизмам в ряду поколений особей данного вида формируются определенные видовые характеристики и вид как реальная единица живой природы существует продолжительное время. Однако у разных представителей вида в силу постоянно идущего мутационного процесса один и тот же набор генов генома представлен разными их аллелями. Так как при половом размножении у многих видов в воспроизведении потомства принимают участие две особи, то совершенно очевидно, что в результате оплодотворения разные зиготы получают неодинаковый набор аллелей в их генотипах. Увеличению генотипического разнообразия представителей вида способствуют также механизмы, приводящие к перекомбинации родительских аллелей особи в ее гаметах. Действительно, если бы гаметы, образуемые организмом, были одинаковы по набору аллелей в их геноме, то у потомков одной пары организмов при раздельнополости или одного гермафродитного организма не наблюдалось бы генотипического разнообразия. В каждом новом поколении вида генотипически различными были бы лишь дети разных родителей.
Реально в природе наблюдается разнообразие потомков одних и тех же родителей. Например, родные братья и сестры различаются не только по полу, но и по другим признакам. Такие различия потомков объясняются тем, что в каждом акте оплодотворения встречаются генетически различающиеся гаметы. Механизмом,обеспечивающим разнообразие гамет, образуемых одним и тем же организмом, является мейоз, в ходе которого происходит не только уменьшение вдвое наследственного материала, попадающего в гаметы, но и эффективное перераспределение родительских аллелей между гаметами. Процессами, приводящими к перекомбинации генов и целых хромосом в половых клетках, являются Кроссинговер и расхождение бивалентов в анафазе I мейоза (см. гл. 5).
Кроссинговер. Этот процесс происходит в профазе I мейоза в то время, когда гомологичные хромосомы тесно сближены в результате конъюгации и образуют биваленты. В ходе кроссинговера осуществляется обмен соответствующими участками между взаимно переплетающимися хроматидами гомологичных хромосом (рис. 3.72). Этот процесс обеспечивает перекомбинацию отцовских и материнских аллелей генов в каждой группе сцепления. В разных предшественниках гамет Кроссинговер происходит в различных участках хромосом, в результате чего образуется большое разнообразие сочетаний родительских аллелей в хромосомах.
157
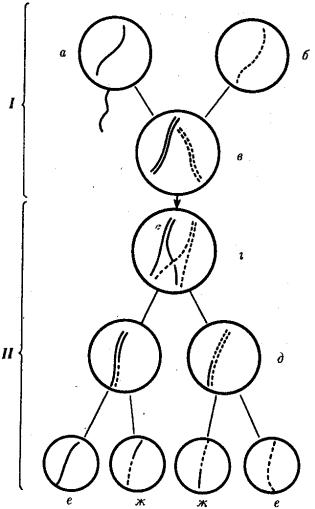
Рис. 3.72. Кроссинговер как источник генетического разнообразия гамет:
I — оплодотворение родительских гамет а и б с образованием зиготы в; II — гаметогенез в организме, развившемся из зиготы в; г — кроссинговер, происходящий между гомологами в профазе I; д — клетки, образовавшиеся после 1- го мейотического деления; е, ж — клетки, образовавшиеся после 2-го деления мейоза (е — некроссоверные гаметы с исходными родительскими хромосомами; ж
— кроссоверные гаметы с перекомбинацией наследственного материала в гомологичных хромосомах)
Понятно, что кроссинговер как механизм рекомбинации эффективен лишь в том случае, когда соответствующие гены отцовской и материнской хромосом представлены разными аллелями. Абсолютно идентичные группы сцепления при кроссинговере не дают новых сочетаний аллелей.
Кроссинговер происходит не только в предшественницах половых клеток при мейозе. Он наблюдается также в соматических клетках при митозе. Соматический кроссинговер описан у дрозофилы, у некоторых видов плесеней. Он осуществляется в ходе митоза между гомологичными хромосомами, однако его частота в 10 000 раз
158

меньше частоты мейотического кроссинговера, от механизма которого он ничем не отличается. В результате митотического кроссинговера появляются клоны соматических клеток, различающихся по содержанию в них аллелей отдельных генов. Если в генотипе зиготы данный ген представлен двумя разными аллелями, то в результате соматического кроссинговера могут появиться клетки с одинаковыми либо отцовскими, либо материнскими аллелями данного гена (рис. 3.73).
Рис. 3.73. Кроссинговер в соматических клетках:
1 — соматическая клетка, в гомологичных хромосомах которой ген А представлен двумя разными аллелями (А и а); 2 — кроссинговер; 3 — результат обмена соответствующими участками между гомологичяыми хромосомами; 4 — расположение гомологов в плоскости экватора веретена деления в метафазе митоза (два варианта); 5 — образование дочерних клеток; 6 — образование гетерозитотиых по гену А клеток, сходных с материнской клеткой по набору аллелей (Аа); 7 — образование гомозиготных по гену А клеток, отличающихся от материнской клетки по набору аллелей (АА или аа)
Расхождение бивалентов в анафазе I мейоза. В метафазе I мейоза в экваториальной плоскости ахромативнового веретена выстраиваются биваленты, состоящие из одной отцовской и одной материнской хромосомы. Расхождение гомологов, которые несут разный набор аллелей генов в анафазе I мейоза, приводит к образованию гамет, отличающихся по аллельному составу отдельных групп сцепления (рис. 3.74).
159

Рис. 3.74. Расхождение гомологичных хромосом в анафазе I мейоза как источник генетического разнообразия гамет:
1—метафаза I мейоза (расположение бивалента в плоскости экватора веретена деления); 2 — анафаза I мейоза (расхождение гомологов, несущих разные аллели гена А к разным полюсам); 3 — второе мейотическое деление (образование двух типов гамет, различающихся по аллелям гена А)
Рис. 3.75. Случайный характер расположения бивалентов в метафазе (1)
инезависимое расхождение их в анафазе (2) первого мейотического деления
В связи с тем что ориентация бивалентов по отношению к полюсам веретена в метафазе I оказывается случайной, в анафазе I мейоза в каждом отдельном случае к разным полюсам направляется гаплоидный набор хромосом, содержащий оригинальную комбинацию родительских групп сцепления (рис. 3.75). Разнообразие гамет, обусловленное независимым поведением бивалентов, тем больше, чем
больше групп сцепления в геноме данного вида. Оно может быть выражено формулой 2n, где п — число хромосом в гаплоидном наборе. Так, у дрозофилы п = 4
иколичество типов гамет, обеспечиваемое перекомбинацией родительских хромосом в них, равно 24 = 16. У человека п = 23, и разнообразие гамет, обусловленное этим механизмом, соответствует 223, или 8388608.
Кроссинговер и процесс расхождения бивалентов в анафазе I мейоза
160