

биофизика
.pdfЧаще всего Na+-К+-АТФаза работает в режиме, когда m = 3/2, m всегда больше 1. (Нет ионных насосов, перекачивающих С1-, поэтому в уравнении Томаса отсутствуют члены PCl[Cl-]).
Коэффициент m > 1 усиливает вклад градиента концентрации калия в создание мембранного потенциала, поэтому мембранный потенциал, рассчитанный по Томасу, больше по абсолютной величине, чем мембранный потенциал, рассчитанный по Гольдману, и дает совпадение с экспериментальными значениями для мелких клеток.
Нарушение биоэнергетических процессов в клетке и работы Na+-К+-АТФазы приводит к уменьшению Vm, в этом случае мембранный потенциал лучше описывается уравнением Гольдмана.
В создании потенциала покоя роль иона хлора по сравнению с ролью иона калия представляется второстепенной. Это связано с тем, что внутриклеточная концентрация хлора очень мала и при малом притоке или оттоке ионов подвергается большим относительным изменениям хлора.
Потенциал действия
При возбуждении нервных и мышечных клеток между внутриклеточной средой и межклеточной жидкостью возникает изменение мембранного потенциала, называемое потенциалом действия (см. рис.). Трансмембранный потенциал действия, показанный на рис. 3.7., типичен для потенциалов, наблюдаемых в эксперименте на нерве и мышце. Во всех случаях в покое мембрана имеет отрицательный потенциал в пределах 60–100 мВ. Процесс активации вызывает внезапное и быстрое увеличение этого потенциала с изменением полярности до пиковых значений, достигающих 40 мВ.
После активации следует фаза реполяризации, восстанавливающая состояние покоя. Потенциал, однако, может на некоторое время выйти на гиперполяризованный или деполяризованный по сравнению с потенциалом покоя. Эти следовые потенциалы, показанные на рисунке, могут присутствовать или отсутствовать, причем в первом случае обычно можно наблюдать только один из них.
51

50 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
50 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
0 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
0 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
0 |
5 |
10 |
|
0 |
5 |
10 |
|
|
|
|
|
0 |
|
400 |
600 |
||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||
-50 |
|
|
|
|
а |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
б |
-50 |
|
|
|
|
|
|
|
|
|
|
в |
|
|
|
|
|||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||||||||
|
|
|
|
|
|
|
|
|
Пик |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Овершут |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||||||
|
|
+40мВ |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
Фаза нарастания |
|
|
|
|
|
|
|
|
|
|
Фаза спада |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||||||
|
|
|
|
Подножие |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Деполяризационный след |
||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||
|
|
-60мВ |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Гиперполяризационный след |
||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||||||
Рис. 3.7. а, б, в. – виды потенциалов действия, г - терминология, применяемая для описания потенциала действия и сопровождающих его следовых потенциалов.
Потенциалом действия (ПД) называется электрический импульс, обусловленный изменением ионной проницаемости мембраны. С ним связано распространение по нервам и мышцам волны возбуждения.
Опыты по исследованию потенциала действия вначале были проведены на гигантских аксонах кальмара при помощи микроэлектродов с использованием высокоомных измерителей напряжения, а также методом меченых атомов. На рисунке показаны схема опытов и результаты исследований.
В опытах по исследованию потенциала действия использовали три микроэлектрода, введенных в аксон. На микроэлектрод Э1 подается импульс с амплитудой V от генератора Г прямоугольных импульсов, меняющий мембранный потенциал. Мембранный потенциал измеряется при помощи второго Э2 и третьего Э3 микроэлектродов высокоомным регистратором напряжения О.
Возбуждающий импульс вызывает лишь на короткое время смещение мембранного потенциала, который быстро пропадает и восстанавливается потенциал покоя. В том случае, когда возбуждающий импульс смещается еще дальше в отрицательную сторону, он сопровождается гиперполяризацией мембраны. Также не формируется потенциал действия, когда возбуждающий импульс положительный (деполяризующий), но его
52

амплитуда меньше порогового значения Vmпор. Если амплитуда положительного, |
||||||
деполяризующего импульса окажется больше значения Vmпор, Vm становится больше |
||||||
Vmпор и в мембране развивается |
процесс, |
в результате которого происходит резкое |
||||
повышение мембранного потенциала и мембранный потенциал Vm даже меняет свой знак |
||||||
- становится положительным (рис. 3.8., б). |
|
|
|
|||
Э1 |
|
Э2 |
Э3 |
|
|
|
Г |
Н |
х |
|
У О |
|
|
|
|
|
|
|
|
|
Э0 |
|
|
|
|
|
|
|
|
а |
|
|
|
|
V10 |
|
|
|
|
|
|
V2 |
|
|
t |
|
|
|
|
|
|
|
|
|
|
0 |
|
|
t |
|
|
|
V3 |
|
|
|
|
|
|
|
|
|
|
|
|
|
0 |
|
|
t |
|
|
|
|
|
t |
|
|
|
|
|
|
|
|
|
|
|
Vm |
Vmрев |
|
|
|
|
|
50 |
|
|
|
|
||
|
|
|
|
|
|
|
0 |
|
|
|
t |
|
|
Vmпор |
Vmп |
Vmд |
|
|
|
|
-60 |
|
|
|
|
|
|
|
Vпор |
|
|
t |
|
|
|
|
|
|
|
|
|
|
|
б |
|
|
|
|
Рис. 3.8. Исследование потенциала действия: а - схема опыта; б - потенциал |
||||||
действия. (Vmп- потенциал покоя, Vmрев- |
потенциал реверсии, Vmd |
- |
амплитуда |
|||
потенциала действия, Vmпор- пороговый потенциал) |
|
|
||||
53
Достигнув некоторого положительного значения Vmрев- потенциала реверсии,
мембранный потенциал возвращается к значению потенциала покоя Vmп, совершив нечто вроде затухающего колебания. В нервных волокнах и скелетных мышцах длительность потенциала действия около 1 мс (а в сердечной мышце около 300 мс). После снятия возбуждения еще в течение 1 – 3 мс в мембране наблюдаются некоторые остаточные явления, во время которых мембрана рефрактерна (невозбудима).
Новый деполяризующий потенциал Vm > Vmпор может вызвать образование нового потенциала действия только после возвращения мембраны в состояние покоя,
причем амплитуда потенциала действия: Vmd = |Vmп| + Vmрев не зависит от амплитуды
деполяризующего потенциала (если только Vm >Vmпор).
Если в покое мембрана поляризована (потенциал цитоплазмы отрицателен по отношению к внеклеточной среде), то при возбуждении происходит деполяризация мембраны (потенциал внутри клетки положителен), и после снятия возбуждения происходит реполяризация мембраны.
Как было указано выше, ответ в возбудимых мембранах не возникает, пока стимул не достигнет некоторого уровня, называемого пороговым потенциалом. Для всех надпороговых стимулов, приложенных к клетке, потенциалы действия одинаковы.
Чтобы стимулирующий ток возбудил мембрану, он должен иметь достаточно большую интенсивность и нужную полярность. Данный стимулирующий импульс должен также иметь достаточную длительность.
Ответы трансмебранного потенциала на импульсы стимулирующего тока для аксона краба показаны на следующем рисунке 3.9.
54

Vm
+0,4 С
С
b
а |
|
0,5 |
1,0 |
t,мс |
|
-0,4
Рис. 3.9. Подпороговые ответы, записанные внеклеточно от аксона краба в окрестности стимулирующих электродов. Аксон был помещен в парафиновое масло, и, следовательно, измеренный внеклеточный потенциал прямо связан с трансмембранным потенциалом. Черный прямоугольник указывает период действия стимула, длительность которого составляла около 50 мкс. По оси ординат отложена шкала напряжений; максимальная амплитуда потенциала действия принята за единицу.
Здесь за нулевой, или опорный, потенциал принят потенциал покоя. Отмеченная на рисунке длительность стимулирующего импульса поддерживалась неизменной, в то время как его амплитуда и знак изменялись.
О стимуле, который заставляет трансмембранный (внутриклеточный минус) потенциал становиться более отрицательным, чем он был в покое, говорят, что он гиперполяризует мембрану. При гиперполяризации возбужение не возникает, сколь бы велик ни был стимул, хотя, как видно на рис. 3.9, пассивный ответ увеличивается с увеличением силы стимула. С другой стороны, для деполяризующих стимулов с увеличивающейся амплитудой достигается уровень, при котором возникает ответ С, показанный на рисунке. На кривой ответа видна нижняя часть вызванного потенциала действия, что свидетельствует о достижении порогового условия.
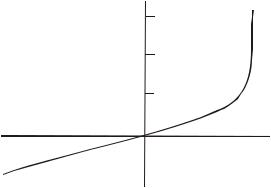
Рассмотрение ответов на подпороговые импульсы показывает, что они в принципе аналогичны тем, которые можно ожидать от пассивной электрической RС-цепочки. Симметричны ли ответы на стимулы противоположной полярности на рисунке, например, ответы а и b.
55
0,3
0,2
0,1
- |
|
|
|
|
|
|
|
|
1,0 |
0,5 |
1,0 |
||||||
-0,5 |
||||||||
-0,1
Рис. 3.10. Связь между стимулом и ответом для аксона краба
В случае деполяризации в ответ на потенциалы, составляющие от 80 до 100% порога, ответ не является зеркальным отражением ответа на гиперполяризующий стимул той же амплитуды и длительности, но противоположной полярности.
Таким образом, ответ не строго пассивен. Отсутствие симметрии свидетельствует о том, что в ответе должна присутствовать нелинейная активная компонента.
Приведенные выше выводы нашли отражение также на данном рисунке, который базируется на предыдущем, и представляет собой график напряжения, измеренного спустя 0,29 мкс после стимула. Напряжение выражено в долях пиковой амплитуды потенциала действия и представлено в виде функции от амплитуды стимула.
Отметим, что зависимость линейна для всех гиперполяризующих стимулов, как можно было бы ожидать при линейной пассивной системе. Линейность наблюдается также для малых деполяризующих сигналов, следовательно, в этой области система тоже может описываться пассивной цепью. Для стимулов с большей амплитудой наблюдаемое поведение становится нелинейным, поэтому она должно рассматриваться как активная.
Уравнение Ходжкина-Хаксли
Структура модели связывает ионные токи с соответствующими движущими силами. Последние описываются как разности между трансмембранным потенциалом и потенциалами Нернста для ионов соответствующих типов.
Главным достижением Ходжкина и Хаксли была их схема для оценки удельных проводимостей gК(t) и gNa(t).
56
gK ( t ) |
|
I K ( t ) |
|
|
g Na ( t ) |
|
I Na ( t ) |
|||
(V |
m |
V K ) |
; |
(V |
m |
V Na ) |
||||
|
|
|||||||||
|
|
m |
|
|
m |
|||||
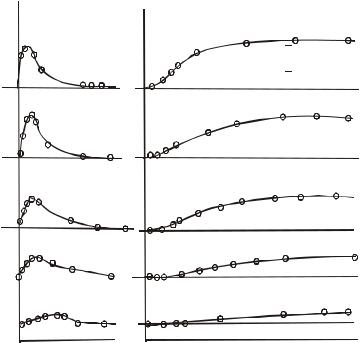
Поскольку при фиксации потенциала знаменатели в выражениях постоянны, gK (t)~ Iк(t) и gNa(t) ~ INa(t). Результаты серии экспериментов с фиксацией потенциала, позволяющие определить IK(t) и INa(t) представлены на следующем рисунке в виде семейства кривых для калиевых и натриевых проводимостей.
Натриевая проводимость |
Калиевая проводимость |
|||||||||||||||
|
|
|
мВ |
|
|
|
|
мВ |
|
|
|
|
|
|
||
|
|
|
109 |
|
|
|
|
109 |
|
|
|
|
10ммО/см2 |
|||
|
|
|
88 |
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
88 |
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
63 |
|
|
|
|
|
63 |
|
|
|
|
|
|
|
|
|
|
38 |
|
|
|
|
|
38 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
26 |
|
|
|
|
|
26 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
0 |
2 |
4 |
|
0 |
2 |
4 |
6 |
8 |
||||||||
Рис. 3.11. Изменения ионных проводимостей, вызываемые деполяризацией с фиксацией напряжения. Кружки представляют значения, полученные на основе экспериментальных измерении ионного тока, а кривые проведены в соответствии с уравнениями. Указанные значения трансмембранного потенциала в милливольтах при фиксации даны относительно уровня покоя, т.е представляют собой vm
Из представленных рисунков следует, что, во-первых: чем ближе смещается фиксированный Vm к значению равновесного потенциала, определяемого по уравнению Нернста для ионов Na+ и К+, тем меньше значение соответствующей проводимости и вовторых, меняется временной ход gк и gNa при изменениях vm.
57
Таким образом, семейство кривых для проводимостей gк и gNa при различных значениях фиксированных vm экспериментально показывает зависимости проводимостей (и, соответственно, токов) от vm и времени.
Впоследствии по семейству полученных кривых были построены зависимости изменения параметров натриевых и калиевых токов в процессе генерации потенциала действия.
Ходжкин и Хаксли построили математическую модель, соответствующую данным приведенного выше рисунка.
Таким образом, Ходжкин и Хаксли обосновали ионную теорию возбудимых мембран и смогли удовлетворительно описать в рамках этой теории изменение ионной проводимости и процесс генерации потенциала действия нервной клетки.
Энергия раздражения и возбуждения, порог раздражения
Способность возбудимой ткани изменять свои свойства или состояние под действием раздражителей называют возбудимостью (ε). Её количественной мерой служит интенсивность порогового раздражителя (Jп), то есть самого слабого стимула, в ответ на который возникает ПД, а вслед за ним и специфическая реакция. Чем ниже Jп, тем выше возбудимость. Следовательно, между Jп и ε имеется обратно пропорциональная зависимость: = 1/Jn.
Реакция возбудимой ткани на раздражитель называется возбуждением. У него много разнообразных проявлений (изменения в возбуждённой ткани обмена веществ, температуры, электрического импеданса, рассеяние света, флуоресценция, двулучепреломление, а также развитие специфических реакций – нервной импульсации и мышечного сокращения), но одним из важнейших является ПД, поскольку он служит непременным звеном развития любой из специфических реакций.
Под действием многих раздражителей изменяется уровень мембранного потенциала. Такая реакция невозбудимых мембран связана, как правило, с изменением их ионной проницаемости за счёт открывания или закрывания неспецифических потенциалнезависимых каналов. Между сдвигом мембранного потенциала невозбудимой мембраны и интенсивностью раздражителя, вызвавшего его, существует линейная зависимость. Это свойство получило название градуальности. Оно присуще невозбудимой мембране во всём диапазоне изменений разности потенциалов на ней.
58

Градуальность свойственна и возбудимой мембране, но только в том случае, если трансмембранная разность потенциалов, изменяясь под действием раздражителя, не достигает критического мембранного потенциала КМП. Такие раздражители, не доводящие деполяризацию до КМП, и, следовательно, не вызывающие потенциал действия ПД, называются подпороговьши для возбудимой ткани. Стимул, вызывающий сдвиг мембранного потенциала до КМП, считается пороговым, поскольку под действием его возникает ПД (возбуждение).
V,мВ |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||
60 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
40 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Vинв |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
20 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
0 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
ПД |
|
|
|
|
|
|
|
|
t,c |
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
-20 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||
-40 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
ПП |
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||
-60 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
КМП |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
-90 |
|
|
|
|
|
|
2 |
3 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||||
1 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||||||||||
Рис. 3.12 Генерация потенциала действия при сдвиге мембранного потенциала от ПП до КМП. По оси абсцисс – время (мс), по оси ординат – разность электрических потенциалов на возбудимой мембране (мВ).
Раздражитель может иметь разную природу (механическую, химическую, электрическую и т. д.), но пороговым он будет тогда, когда сдвинет уровень мембранного потенциала от ПП до КМП: Vп = │ПП│ – │КМП│. Надпороговые (более сильные) раздражители возбудимой мембраны тем более вызывают ПД.
Понятно, что градуальность характерна и для отклонений трансмембранной разности потенциалов (от уровня ПП) в сторону, противоположную деполяризации (в аксоне кальмара от -85 до -90 мВ и более). Такой сдвиг мембранного потенциала называют гиперполяризацией. Следовательно, градуальность присуща возбудимой мембране при любой её гиперполяризации и при подпороговой (до КМП) деполяризации.
59
Рефрактерные периоды, реобаза, хронаксия
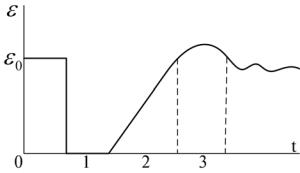
Рефрактерность – изменение возбудимости при возбуждении. Выделяют три основные стадии. Их принято называть фазами (см. рис.3.13).
Рис.3.13. Динамика возбудимости при возбуждении (рефрактерные фазы). Обозначения: 1 – абсолютно рефрактерная фаза; 2 – относительно рефрактерная фаза; 3 – фаза экзальтации. По оси абсцисс – время (t); по оси ординат – уровень возбудимости ; 0
–исходный уровень возбудимости.
Развитие возбуждения вначале сопровождается полной утратой возбудимости (ε = 0). Это состояние называют абсолютно рефрактерной фазой (АРФ). Она соответствует времени деполяризации возбудимой мембраны. В течение АРФ возбудимая мембрана не может генерировать новый ПД, даже если на неё подействовать сколь угодно сильным раздражителем (Jп ). Природа АРФ состоит в том, что во время деполяризации все потенциалзависимые ионные каналы находятся в открытом состоянии, и дополнительные стимулы не могут вызвать воротный процесс.
АРФ сменяется относительно рефрактерной фазой (ОРФ), в течение которой возбудимость от нуля возвращается к исходному уровню (ε0). ОРФ совпадает с реполяризацией возбудимой мембраны. С течением времени во всё большем числе потенциалзависимых ионных каналов завершаются воротные процессы, с которыми было связано предшествующее возбуждение, и каналы вновь обретают способность к следующему переходу из закрытого в открытое состояние под действием очередного стимула. Во время ОРФ пороги возбуждения постепенно снижаются J n0 J n и,
следовательно, возбудимость восстанавливается до исходного уровня (0 < ε < ε0).
За ОРФ следует фаза экзальтации (ФЭ), для которой характерна повышенная возбудимость (ε > ε0). Она, очевидно, связана с изменением свойств сенсора напряжения во время возбуждения. За счёт перестройки конформации белковых молекул изменяются
60