

1.Кле́тка — элементарная единица строения и жизнедеятельности всех организмов (кроме вирусов), обладающая собственным обменом веществ, способная к самостоятельному существованию, самовоспроизведению и развитию.
-
Клетки многоклеточного животного организма разнообразны. Значительно различаются по форме, размерам, по строению и функциям клетки крови, мышечные и нервные клетки, сперматозоиды и клетки желез.
Клетки растений также достаточно разнообразны: это клетки эпидермы и мезофила, ситовидные трубочки и волокна колленхимы.
Однако исходя из постулата Р. Вирхова о гомологичности клеток, можно говорить о едином плане строения всех клеток: они состоят из поверхностного аппарата, ядра и цитоплазмы.
-
Мембраны биологические
(лат. membrana оболочка, перепонка)
функционально активные поверхностные структуры толщиной в несколько молекулярных слоев, ограничивающие цитоплазму и большинство органелл клетки, а также образующие единую внутриклеточную систему канальцев, складок, замкнутых областей.
Биологические мембраны имеются во всех клетках. Различают митохондриальные, ядерные, лизосомные мембраны, мембраны пластинчатого комплекса аппарата Гольджи, эндоплазматического ретикулума, саркоплазматического ретикулума и др. Химический состав и строение биологических мембран. Состав М.б. зависит от их типа и функций, однако основными составляющими являются Липиды и Белки, а также Углеводы
Функции биологических мембран следующие:
1.Отграничивают содержимое клетки от внешней среды и содержимое органелл от цитоплазмы.
2.Обеспечивают транспорт веществ в клетку и из нее, из цитоплазмы в органеллы и наоборот.
3.Выполняют роль рецепторов (получение и преобразование сит-налов из окружающей среды, узнавание веществ клеток и т. д.).
4.Являются катализаторами (обеспечение примембранных химических процессов).
5.Участвуют в преобразовании энергии.
-
Включения цитоплазмы — это необязательные компоненты клетки, появляющиеся и исчезающие в зависимости от интенсивности и характера обмена веществ в клетке и от условий существования организма. Включения имеют вид зерен, глыбок, капель, вакуолей, гранул различной величины и формы. Их химическая природа очень разнообразна. В зависимости от функционального назначения включения объединяют в группы:
трофические;
секреты;
инкреты;
пигменты;
экскреты и др.
специальные включения (гемоглобин)
Среди трофических включений (запасных питательных веществ) важную роль играют жиры и углеводы. Белки как трофические включения используются лишь в редких случаях (в яйцеклетках в виде желточных зерен).
Пигментные включения придают клеткам и тканям определенную окраску.
Секреты и инкреты накапливаются в железистых клетках, так как являются специфическими продуктами их функциональной активности.
Экскреты - конечные продукты жизнедеятельности клетки, подлежащие удалению из нее.
2.Плазмолемма. ХИМИЧЕСКИЙ СОСТАВ ПЛАЗМОЛЕММЫ.
1) собственно мембрана;
2) надмембранный слой – гликокаликс;
3) субмембранный (подмембранный) слой.
Функции плазмолеммы. Эта мембрана выполняет ряд важнейших клеточных функций, ведущими из которых являются барьерная функция (разграничения цитоплазмы с внешней средой), функции рецепции и транспорта различных веществ как внутрь клетки, так и из нее.
Состав мембран
-
Липиды
-
Углеводы
-
Белки
Эндоцитоз — это транспорт макромолекул через плазмолемму. Выполняя транспортную функцию, плазмолемма обеспечивает диффузию (пассивный перенос) ряда веществ, например воды, ионов, некоторых низкомолекулярных соединений. Крупные молекулы биополимеров практически не проникают сквозь плазмолемму. В ряде случаев макромолекулы и даже их агрегаты, а часто и крупные частицы попадают внутрь клетки в результате процессов эндоцитоза (рис. 6, А, Б). Эндоцитоз формально разделяют нафагоцитоз (захват и поглощение клеткой крупных частиц, например бактерий или даже фрагментов других клеток), и пиноцитоз (захват макромолекулярных соединений).
Эндоцитоз начинается с сорбции на поверхности плазмолеммы поглощаемых веществ. Связывание их с плазмолеммой определяется наличием на ее поверхности рецепторных молекул. После сорбции веществ на поверхности плазмолемма начинает образовывать сначала небольшие впячивания внутрь клетки. Эти впячивания могут иметь вид еще незамкнутых округлых пузырьков или представлять собой глубокие инвагинации, впячивания внутрь клетки. Затем такие локальные впячивания отшнуровываются от плазмолеммы и в виде пузырьков свободно располагаются под ней.
В дальнейшем эндоцитозные пузырьки могут сливаться друг с другом, расти и в их внутренней полости, кроме поглощенных веществ, начинают обнаруживаться гидролитические ферменты (гидролазы), поступающие сюда из лизосом (см. дальше). Эти ферменты расщепляют биополимеры до мономеров, которые в результате активного транспорта через мембрану пузырька переходят в гиа-лоплазму. Таким образом, поглощенные молекулы внутри мембранных вакуолей, образовавшихся из элементов плазмолеммы, подвергаются внутриклеточному пищеварению. Плазмолемма принимает участие в выведении веществ из клетки (т.н. экзоцитоз). В этом случае внутриклеточные продукты (белки, мукополисахариды, жировые капли и др.), заключенные в вакуоли или пузырьки и отграниченные от гиалоплазмы мембраной, подходят к плазмолемме. В местах контактов плазмолемма и мембрана вакуоли сливаются и содержимое вакуоли поступает в окружающую среду.
Процесс эндоцитоза и экзоцитоза осуществляется при участии связанной с плазмолеммой системы фибриллярных компонентов цитоплазмы — таких, как микротрубочки и сократимые микрофиламенты. Последние, соединяясь с определенными участками плазмолеммы, могут, изменяя свою длину, втягивать мембрану внутрь клетки, что приводит к отделению от плазмолеммы эндоцитозных вакуолей. Часто, непосредственно примыкая к ней, микрофиламенты образуют сплошной, так называемый кортикальный слой.
Плазмолемма многих клеток животных может образовывать выросты различной структуры. У ряда клеток такие выросты включают в свой состав специальные компоненты цитоплазмы (микротрубочки, микрофибриллы), что приводит к развитию специальных структур — микроворсинок, ресничек, жгутиков.
Межклеточные соединения (контакты)
 Рис.
7. Межклеточные соединения (схема). 1 —
простое соединение; 2 — пальцевидное
соединение; 3 — десмосома; 4 — плотное
соединение; 5 — щелевидное соединение
(нексус).
Рис.
7. Межклеточные соединения (схема). 1 —
простое соединение; 2 — пальцевидное
соединение; 3 — десмосома; 4 — плотное
соединение; 5 — щелевидное соединение
(нексус).
Плазмолемма многоклеточных животных организмов принимает активное участие в образовании специальных структур — межклеточных соединений (junctiones intercellulares), обеспечивающих межклеточные взаимодействия. Различают несколько типов таких структур (рис. 7).
Простое межклеточное соединение, (junctio intercellularis simplex) — сближение плазмолемм соседних клеток на расстояние 15—20 нм. При этом происходит взаимодействие слоев гликокаликса соседних клеток. Разновидностью простого соединения является "пальцевидное", или соединение по типу замка.
Плотное соединение (запирающая зона) (zonula occludens) — зона, где слои двух плазмолемм максимально сближены, здесь происходит как бы слияние участков плазмолемм двух соседних клеток. Роль плотного замыкающего соединения заключается в механическом соединении клеток друг с другом. Эта область непроницаема для макромолекул и ионов и, следовательно, она запирает, отграничивает межклеточные щели (и вместе с ними собственно внутреннюю среду организма) от внешней среды.
Часто встречается, особенно в эпителии, особый тип соединения — пятно сцепления, или десмосома (desmosoma). Эта структура представляет собой небольшую площадку, иногда имеющую слоистый вид, диаметром до 0,5 мкм, где между мембранами располагается зона с высокой электронной плотностью. К плазмолемме в зоне десмосомы со стороны цитоплазмы прилегает участок электронноплотного вещества, так что внутренний слой мембраны кажется утолщенным. Под этим утолщением находится область тонких фибрилл, которые могут быть погружены в относительно плотный матрикс. Функциональная роль десмосом заключается главным образом в механической связи между клетками.
Щелевидное соединение, или нексус (nexus), представляет собой область протяженностью 0,5—3 мкм, где плазмолеммы разделены промежутком в 2—3 нм. Со стороны цитоплазмы никаких специальных примембранных структур в данной области не обнаруживается, но в структуре плазмолемм соседних клеток друг против друга располагаются специальные белковые комплексы (коннексоны), которые образуют как бы каналы из одной клетки в другую. Этот тип соединения встречается во всех группах тканей. Функциональная роль щелевидного соединения заключается в переносе ионов и мелких молекул от клетки к клетке. Так, в сердечной мышце возбуждение, в основе которого лежит процесс изменения ионной проницаемости, передается от клетки к клетке через нексус.
Синаптические соединения, или синапсы (synapsis). Этот тип соединений характерен для нервной ткани и встречается в специализированных участках контакта как между двумя нейронами, так и между нейроном и каким-либо иным элементом, входящим в состав рецептора или эффектора (например, нервно-мышечные, нервно-эпителиальные синапсы). Синапсы — участки контактов двух клеток, специализированных для односторонней передачи возбуждения или торможения от одного элемента к другому.
3. Органеллы — постоянно присутствующие и обязательные для всех клеток микроструктуры, выполняющие жизненно важные функции.
Классификация органелл. Различают мембранные и немембраные органеллы. К мембранным органеллам относятся митохондрии, эндоплазматическая сеть, аппарат Гольджи, лизосомы. Немембранные органеллы: свободные рибосомы и полисомы, микротрубочки, центриоли и филаменты (микрофиламенты, промежуточные филаменты). Во многих клетках органеллы могут принимать участие в образовании особых структур, характерных для специализированных клеток. Так, реснички и жгутики образуются за счет центриолей и плазматической мембраны, микроворсинки — это выросты плазматической мембраны с гиалоплазмой и микрофиламентами, акросома спермиев — это производное элементов аппарата Гольджи, «эллипсоид» зрительных клеток — скопления митохондрий и пр.
-
Митохондрии (mitochondriae) — органеллы синтеза АТФ. Их основная функция связана с окислением органических соединений и использованием освобождающейся при распаде этих соединений энергии для синтеза молекул АТФ. Исходя из этого, митохондрии часто называют энергетическими станциями клетки, или органеллами клеточного дыхания.
Термин «митохондрия» был введен Бенда в 1897 г. для обозначения зернистых и нитчатых структур в цитоплазме разных клеток. Митохондрии можно наблюдать в живых клетках, так как они обладают достаточно высокой плотностью. В живых клетках митохондрии могут перемещаться, сливаться друг с другом, делиться. Форма и размеры митохондрий животных клеток разнообразны, но в среднем толщина их около 0,5 мкм, а длина — от 1 до 10 мкм. Подсчеты показывают, что количество их в клетках сильно варьирует — от единичных элементов до сотен. Так, в клетке печени они составляют более 20% общего объема цитоплазмы и содержат около 30—35% общего количества белка в клетке. Площадь поверхности всех митохондрий печеночной клетки в 4—5 раз больше поверхности ее плазматической мембраны.
 Рис.
12. Ультрамикроскопическое строение
митохондрии. А — схема; Б — электронная
микрофотография среза митохондрии
печеночной клетки; 1 — наружная
митохондриальная мембрана; 2 — внутренняя
митохондриальная мембрана; 3 — кристы;
4 — митохондриальный матрикс.
Рис.
12. Ультрамикроскопическое строение
митохондрии. А — схема; Б — электронная
микрофотография среза митохондрии
печеночной клетки; 1 — наружная
митохондриальная мембрана; 2 — внутренняя
митохондриальная мембрана; 3 — кристы;
4 — митохондриальный матрикс.
Обычно митохондрии скапливаются вблизи тех участков цитоплазмы, где возникает потребность в АТФ. Так, в сердечной мышце митохондрии находятся вблизи миофибрилл. В сперматозоидах митохондрии образуют спиральный футляр вокруг оси жгутика и т. д. Увеличение числа митохондрий в клетках происходит путем деления, или почкования, исходных митохондрий.
Митохондрии ограничены двумя мембранами толщиной около 7 нм (рис. 12. А). Наружная митохондриальная мембрана (membrana mitochondrialis externa) отделяет их от гиалоплазмы. Обычно она имеет ровные контуры и замкнута, так что представляет собой мембранный мешок. Внешнюю мембрану от внутренней отделяет межмембранное пространство шириной около 10—20 нм. Внутренняя митохондриальная мембрана (membrana mitochondrialis interna) ограничивает собственно внутреннее содержимое митохондрии, ее матрикс (matrix mitochondrialis). Характерной чертой внутренних мембран митохондрий является их способность образовывать многочисленные впячивания внутрь митохондрий. Такие впячивания чаще всего имеют вид плоских гребней, или крист (crista).
Матрикс митохондрий имеет тонкозернистое строение (рис. 12, Б) в нем иногда выявляются тонкие нити (толщиной около 2—3 нм) и гранулы размером около 15—20 нм. Нити матрикса митохондрий представляют собой молекулы ДНК, а мелкие гранулы — митохондриальные рибосомы.
Основной функцией митохондрий является синтез аденозин-трифосфата (АТФ), происходящий в результате процессов окисления органических субстратов и фосфорилирования АДФ. Начальные этапы этих сложных процессов совершаются в гиалоплазме. Здесь происходит первичное окисление субстратов (например, сахаров) до пировиноградной кислоты (пирувата, ПВК) с одновременным синтезом небольшого количества АТФ. Эти процессы совершаются в отсутствие кислорода (анаэробное окисление, гликолиз). Все последующие этапы выработки энергии (дыхания) — аэробное окисление и синтез основной массы АТФ — осуществляются с потреблением кислорода и локализуются внутри митохондрий. При этом происходит дальнейшее окисление пирувата и других субстратов энергетического обмена с выделением CO2 и переносом протонов на их акцепторы. Эти реакции осуществляются с помощью ряда ферментов так называемого цикла трикарбоновых кислот, которые локализованы в матриксе митохондрии.
В мембранах крист митохондрии располагаются системы дальнейшего переноса электронов и сопряженного с ним фосфорилирования АДФ (окислительное фосфорилирование). При этом происходит перенос электронов от одного белка-акцептора электронов к другому и, наконец, связывание их с кислородом, вследствие чего образуется вода. Одновременно с этим часть энергии, выделяемой при таком окислении в цепи переноса электронов, запасается в виде макроэргической связи при фосфорилировании АДФ, что приводит к образованию большого числа молекул АТФ — основного внутриклеточного энергетического эквивалента. Именно на мембранах крист митохондрии происходит процесс окислительного фосфорилирования с помощью здесь расположенных белков цепи окисления и ферментов фосфорилирования АДФ, АТФ-синтетазы.
Выявлено, что в матриксе митохондрии локализуется автономная система митохондриального белкового синтеза. Она представлена молекулами собственной ДНК, свободными от гистонов, что сближает их с ДНК бактериальных клеток. На этих ДНК происходит синтез молекул РНК разных типов: информационных, трансферных (транспортных) и рибосомных. В матриксе митохондрий наблюдается образование рибосом, отличных от рибосом цитоплазмы. Эти рибосомы участвуют в синтезе ряда митохондриальных белков, не кодируемых ядром клетки.
Однако такая система белкового синтеза не обеспечивает всех функций митохондрии, поэтому автономию митохондрий можно считать ограниченной, относительной. Малые размеры молекул митохондриальных ДНК не могут определить синтез всех белков митохондрий. Показано, что большинство белков митохондрий находится под генетическим контролем со стороны клеточного ядра и синтезируется в цитоплазме. Наиболее вероятно, что митохондриальная ДНК кодирует лишь немногие митохондриальные белки, которые локализованы в мембранах и представляют собой структурные белки, ответственные за правильную интеграцию в митохондриальных мембранах отдельных функциональных белковых комплексов.
Митохондрии в клетках могут увеличиваться в размерах и числе. В последнем случае происходит деление перетяжкой или фрагментация исходных крупных митохондрий на более мелкие, которые в свою очередь могут расти и снова делиться.
4. Лизосомы
Лизосомы (lysosomae) — это разнообразный класс шаровидных структур размером 0,2—0,4 мкм, ограниченных одиночной мембраной. Характерным признаком лизосом является наличие в них гидролитических ферментов — гидролаз, расщепляющих различные биополимеры. Примеры лизосомных гидролаз: фосфатазы, протеиназы, липазы, etc. Лизосомы были открыты в 1949 г. де Дювом.
Среди лизосом можно выделить по крайней мере 3 типа: первичные лизосомы, вторичные лизосомы (фаголизосомы и аутофагосомы) и остаточные тельца (рис. 11). Разнообразие морфологии лизосом объясняется тем, что эти частицы участвуют в процессах внутриклеточного переваривания, образуя сложные пищеварительные вакуоли как экзогенного (внеклеточного), так и эндогенного (внутриклеточного) происхождения.
Первичные лизосомы представляют собой мелкие мембранные пузырьки размером около 0,2—0,5 мкм, заполненные бесструктурным веществом, содержащим гидролазы, в том числе активную кислую фосфатазу, которая является маркерным для лизосом ферментом. Эти мелкие пузырьки практически очень трудно отличить от мелких везикул на периферии зоны комплекса Гольджи, которые также содержат кислую фосфатазу. Местом ее синтеза является гранулярная эндоплазматическая сеть, затем этот фермент появляется в проксимальных участках диктиосом, а затем в мелких везикулах по периферии диктиосом и, наконец, в первичных лизосомах. Таким образом, весь путь образования первичных лизосом очень сходен с образованием секреторных (зимогенных) гранул в клетках поджелудочной железы, за исключением последнего этапа — выбрасывания из клетки.
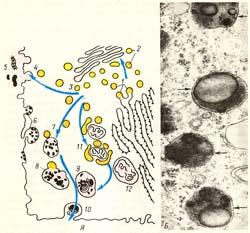 Рис.
11. Строение лизосом. А — схема
участия структур клетки в образовании
лизосом и во внутриклеточном пищеварении:
1 — образование из гранулярной
эндоплазматической сети мелких пузырьков,
содержащих гидролитические ферменты;
2— перенос ферментов в комплекс Гольджи;
3 — образование первичных лизосом; 4 —
выделение и использование (5) гидролаз
при внеклеточном расщеплении; 6 —
эндоцитозные пузырьки; 7 — слияние
первичных лизосом и эндоцитозных
пузырьков; 8 — образование вторичных
лизосом (фаголизосом); 9 — телолизосомы;
10 — экскреция остаточных телец; 11 —
слияние первичных лизосом с разрушающимися
структурами клетки; 12 — аутофагосома.
Б —
электронная микрофотография среза
вторичных лизосом (обозначены стрелками).
Рис.
11. Строение лизосом. А — схема
участия структур клетки в образовании
лизосом и во внутриклеточном пищеварении:
1 — образование из гранулярной
эндоплазматической сети мелких пузырьков,
содержащих гидролитические ферменты;
2— перенос ферментов в комплекс Гольджи;
3 — образование первичных лизосом; 4 —
выделение и использование (5) гидролаз
при внеклеточном расщеплении; 6 —
эндоцитозные пузырьки; 7 — слияние
первичных лизосом и эндоцитозных
пузырьков; 8 — образование вторичных
лизосом (фаголизосом); 9 — телолизосомы;
10 — экскреция остаточных телец; 11 —
слияние первичных лизосом с разрушающимися
структурами клетки; 12 — аутофагосома.
Б —
электронная микрофотография среза
вторичных лизосом (обозначены стрелками).
Вторичные лизосомы, или внутриклеточные пищеварительные вакуоли, формируются при слиянии первичных лизосом с фагоцитарными вакуолями (фагосомами) или пиноцитозными вакуолями, образуя фаголизосомы, или гетерофагосомы. Первичные лизосомы могут сливаться с измененными органеллами самой клетки, подвергающимися перевариванию (т.н. аутофагосомы). При этом ферменты первичной лизосомы получают доступ к субстратам, которые они и начинают расщеплять. Вещества, попавшие в состав вторичной лизосомы, расщепляются гидролазами до мономеров, которые транспортируются через мембрану лизосомы в гиалоплазму, где они реутилизируются, т. е. включаются в различные обменные процессы.
Однако расщепление, переваривание биогенных макромолекул внутри лизосом может идти в ряде клеток не до конца. В этом случае в полостях лизосом накапливаются непереваренные продукты. Такая лизосома носит название «телолизосома», или остаточное тельце (corpusculum residuale). Остаточные тельца содержат меньше гидролитических ферментов, в них происходит уплотнение содержимого, его перестройка. Часто в остаточных тельцах наблюдается вторичная структуризация неперевариваемых липидов, которые образуют слоистые структуры. Там же происходит отложение пигментных веществ. Так, у человека при старении организма в клетках мозга, печени и в мышечных волокнах в телолизосомах происходит отложение т.н. «пигмента старения» — липофусцина.
При участии лизосом в переваривании внутриклеточных элементов (аутолизосомы) они могут обеспечивать модификацию продуктов, приготавливаемых самой клеткой, например, с помощью гидролаз лизосом. В клетках щитовидной железы гидролизуется тироглобулин, что приводит к образованию гормона тироксина, который затем выводится в кровеносное русло.
В аутофагосомах обнаруживаются фрагменты или даже целые цитоплазматические структуры, например митохондрии, элементы цитоплазматической сети, рибосомы, гранулы гликогена и др., что является доказательством их определяющей роли в процессах деградации.
Функциональное значение аутофагоцитоза
еще не ясно. Есть предположение, что
этот процесс связан с отбором и
уничтожением измененных, поврежденных
клеточных компонентов. В этом случае
лизосомы выполняют роль внутриклеточных
«чистильщиков», убирающих дефектные
структуры. Интересно, что в нормальных
условиях число аутофагосом увеличивается
при метаболических стрессах, например
при гормональной индукции активности
клеток печени. Значительно возрастает
число аутофагосом при различных
повреждениях клеток; в этом случае
аутофагоцитозу могут подвергаться
целые зоны внутри клеток. Увеличение
числа аутолизосом в клетках при
патологических процессах — обычное
явление.
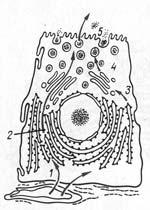 Рис.
10. Участие клеточных структур в белковой
секреции (схема). 1 — поступление
аминокислот из гемокапилляра к рибосомам
гранулярной эндоплазматической сети;
2 — синтез и сегрегация белков; 3 —
переход белков в вакуоли комплекса
Гольджи; 4 — отщепление от комплекса
Гольджи пузырьков с секреторными
продуктами; 5 — экструзия, выход секрета
из клетки.
Рис.
10. Участие клеточных структур в белковой
секреции (схема). 1 — поступление
аминокислот из гемокапилляра к рибосомам
гранулярной эндоплазматической сети;
2 — синтез и сегрегация белков; 3 —
переход белков в вакуоли комплекса
Гольджи; 4 — отщепление от комплекса
Гольджи пузырьков с секреторными
продуктами; 5 — экструзия, выход секрета
из клетки.
5. Комплекс Гольджи (внутренний сетчатый аппарат)
В 1898 г. К. Гольджи, используя свойства связывания тяжелых металлов (осмия или серебра) с клеточными структурами, выявил в нервных клетках сетчатые образования, которые он назвал внутренним сетчатым аппаратом (apparatus reticularis internus). Этот сетчатый аппарат позднее стали называть комплексом Гольджи (complexus Golgiensis). Подобные структуры описаны во всех клетках эукариот.
 Рис.
9. Комплекс Гольджи. А — нервная клетка
спинного мозга, импрегнация серебром
по методу Гольджи: 1 — ядро; 2 — ядрышко;
3 — комплекс Гольджи. Б — схема
ультрамикроскопического строения
(трехмерная реконструкция). В — комплекс
Гольджи на ультратонком срезе (печеночная
клетка). 1 — пузырьки; 2 — трубочки; 3 —
уплощенные мешочки (цистерны); 4 —
пластинки гранулярной эндоплазматической
сети.
Рис.
9. Комплекс Гольджи. А — нервная клетка
спинного мозга, импрегнация серебром
по методу Гольджи: 1 — ядро; 2 — ядрышко;
3 — комплекс Гольджи. Б — схема
ультрамикроскопического строения
(трехмерная реконструкция). В — комплекс
Гольджи на ультратонком срезе (печеночная
клетка). 1 — пузырьки; 2 — трубочки; 3 —
уплощенные мешочки (цистерны); 4 —
пластинки гранулярной эндоплазматической
сети.
При рассмотрении в электронном микроскопе комплекс Гольджи представлен мембранными структурами, собранными вместе в небольшой зоне (рис. 9). Отдельная зона скопления этих мембран называется диктиосомой. Таких зон в клетке может быть несколько. В диктиосоме комплекса Гольджи плотно друг к другу (на расстоянии 20—25 нм) расположены 5—10 плоских цистерн, между которыми располагаются тонкие прослойки гиалоплазмы. Каждая цистерна имеет переменную толщину: в центре ее мембраны могут быть сближены (до 25 нм), а на периферии иметь расширения, ампулы, ширина которых непостоянна. Кроме плотно расположенных плоских цистерн, в зоне комплекса Гольджи наблюдается множество мелких пузырьков (везикул), которые встречаются главным образом в его периферических участках. Иногда видно, как они отшнуровываются от ампулярных расширений на краях плоских цистерн. Принято различать в зоне диктиосомы проксимальный (cis-полюс) и дистальный (trans-полюс) участки. В секретирующих клетках обычно комплекс Гольджи поляризован: его проксимальная часть обращена к ядру, в то время как дистальная —к поверхности клетки.
В клетках отдельные диктиосомы могут быть связаны друг с другом системой везикул и цистерн, примыкающих к проксимальному концу скопления плоских мешков так, что образуется рыхлая трехмерная сеть, выявляемая в световом микроскопе.
Комплекс Гольджи участвует в сегрегации и накоплении продуктов, синтезированных в цитоплазматической сети, в их химических перестройках, созревании; в цистернах комплекса Гольджи происходит синтез полисахаридов, их связывание с белками, что приводит к образованию мукопротеидов, и, главное, с помощью элементов аппарата Гольджи происходит процесс выведения готовых секретов за пределы клетки. Кроме того, комплекс Гольджи обеспечивает формирование клеточных лизосом.
Секреторная функция комплекса Гольджи заключается в том, что синтезированный на рибосомах экспортируемый белок отделяется и накапливается внутри цистерн эндоплазматической сети, по которым он транспортируется к зоне мембран пластинчатого комплекса (рис. 10). Затем накопленный белок может конденсироваться, образуя секреторные белковые гранулы (как это наблюдается в поджелудочной, молочной и других железах), или же оставаться в растворенном виде (как иммуноглобулины в плазматических клетках).
В дальнейшем от ампулярных расширений цистерн комплекса Гольджи отщепляются пузырьки, содержащие эти белки. Такие везикулы также могут сливаться друг с другом и увеличиваться в размерах, образуя секреторные гранулы. После этого секреторные гранулы начинают двигаться к поверхности клетки, соприкасаются с плазмолеммой, с которой сливаются их собственные мембраны, и таким образом содержимое гранул оказывается за пределами клетки. Морфологически этот процесс называется экструзией (выбрасывание, экзоцитоз), напоминает пиноцитоз только с обратной последовательностью стадий.
Н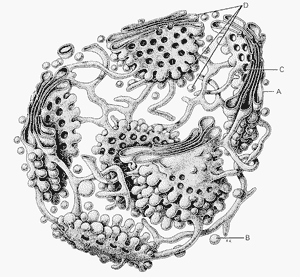 ужно
отметить, что с самого момента образования
до выведения из клеток секретируемые
продукты отделены мембраной от
гиалоплазмы. Следовательно, мембраны
комплекса Гольджи выполняют сегрегирующую
роль при образовании клеточных секретов.
В зоне комплекса Гольджи могут происходить
многие метаболические процессы. Здесь
большинство белков подвергается
модификации, некоторые их аминокислоты
фосфорилируются, ацетилируются или
глюкозилируются. Во многие секреторные
продукты входят сложные белки —
гликопротеиды и мукопротеиды (муцины)
— белки, связанные в единую цепь с
сахарами и полисахаридами разной
природы. Синтез этих полисахаридов идет
в комплексе Гольджи.
ужно
отметить, что с самого момента образования
до выведения из клеток секретируемые
продукты отделены мембраной от
гиалоплазмы. Следовательно, мембраны
комплекса Гольджи выполняют сегрегирующую
роль при образовании клеточных секретов.
В зоне комплекса Гольджи могут происходить
многие метаболические процессы. Здесь
большинство белков подвергается
модификации, некоторые их аминокислоты
фосфорилируются, ацетилируются или
глюкозилируются. Во многие секреторные
продукты входят сложные белки —
гликопротеиды и мукопротеиды (муцины)
— белки, связанные в единую цепь с
сахарами и полисахаридами разной
природы. Синтез этих полисахаридов идет
в комплексе Гольджи.
В пузырьках комплекса Гольджи иногда происходит накопление ресинтезированных молекул липидов и образование сложных белков липопротеидов, которые могут транспортироваться пузырьками за пределы клетки.
Мембраны комплекса Гольджи образуются при участии гранулярной эндоплазматической сети.
6. Агранулярная (гладкая) эндоплазматическая сеть(reticulum endoplasmaticum nongranulosum) также представлена мембранами, образующими мелкие вакуоли и трубки, канальцы, которые могут ветвиться, сливаться друг с другом. В отличие от гранулярной эндоплазматической сети на мембранах гладкой эндоплазматической сети нет рибосом. Диаметр вакуолей и канальцев гладкой эндоплазматической сети обычно около 50—100 нм.
Гладкая эндоплазматическая сеть возникает и развивается за счет гранулярной эндоплазматической сети (при освобождении ее от рибосом).
Деятельность гладкой эндоплазматической сети связана с метаболизмом липидов и некоторых внутриклеточных полисахаридов. Гладкая эндоплазматическая сеть участвует в заключительных этапах синтеза липидов. Она сильно развита в клетках, секретирующих такие категории липидов, как стероиды, например, в клетках коркового вещества надпочечников, в сустентоцитах семенников.
Тесная топографическая связь гладкой эндоплазматической сети с отложениями гликогена (запасной внутриклеточный полисахарид животных) в гиалоплазме различных клеток (клетки печени, мышечные волокна) указывает на ее возможное участие в метаболизме углеводов.
В поперечнополосатых мышечных волокнах гладкая эндоплазматическая сеть способна депонировать ионы кальция, необходимые для функции мышечной ткани.
Очень важна роль гладкой эндоплазматической сети в дезактивации различных вредных для организма веществ за счет их окисления с помощью ряда специальных ферментов. Особенно четко она проявляется в клетках печени. Так, при ряде отравлений в клетках печени появляются ацидофильные зоны (не содержащие РНК), сплошь занятые гладким эндоплазматическим ретикулумом.
7. К немембранным органеллам клетки относятся центриоли, микротрубочки, филаменты, рибосомы и полисомы.
Центриоли , обычно их две (диплосома), представляют собой мелкие тельца, окруженные плотным участком цитоплазмы. От каждой центриоли лучеобразно отходят микротрубочки, получившие название центросферы. Диплосома (две центриоли) и центросфера образуют клеточный центр, который располагается или возле ядра клетки, или возле поверхности комплекса Гольджи. Центриоли в диплосоме расположены под углом друг к другу. Каждая центриоль представляет собой цилиндр, стенка которого состоит из микротрубочек длиной около 0,5 мкм и диаметром около 0,25 мкм. Центриоли являются полуавтономными самообновляющимися структурами, которые удваиваются при делении клетки. Вначале центриоли расходятся в стороны, и возле каждой из них образуется дочерняя центриоль. Таким образом, перед делением в клетке имеются две попарно соединенные центриоли две диплосомы.
Микротрубочки представляют собой различной длины полые цилиндры диаметром 20-30 нм. Многие микротрубочки входят в состав центросферы, где они имеют радиальное направление. Другие микротрубочки расположены под цитолеммой, в апикальной части клетки. Здесь они вместе с пучками микрофиламентов образуют внутриклеточную трехмерную сеть. Стенки микротрубочек имеют толщину 6-8 нм. Микротрубочки образуют цитоскелет клетки и участвуют в транспорте веществ внутри нее.
Цитоскелет клетки представляет собой трехмерную сеть, в которой различные белковые нити связаны между собой поперечными мостиками. В образовании цитоскелета, помимо микротрубочек, участвуют также актиновые, миозиновые и промежуточные филаменты, которые выполняют не только опорную, но и двигательную функцию клетки.
Рибосомы имеются во всех клетках, они участвуют в образовании белковых молекул - в синтезе белка. Размер рибосомы 20х30 нм. Это сложные рибонуклеопротеиды, состоящие из белков и молекул РНК в соотношении 1: 1. Различают рибосомы одиночные - монорибосомы и собранные в группы - полирибосомы, или полисомы. Рибосомы располагаются свободно на поверхности мембран, в результате чего образуется зернистая (гранулярная) эндоплазматическая сеть.
Включения (клеточные гранулы) образуются в результате жизнедеятельности клеток. Их появление зависит от характера обменных процессов в клетке. Различают трофические включения: жировые, белковые, которые мотуг накапливаться в гиалоплазме в качестве резервных материалов, необходимых для жизнедеятельности клетки. К этим же включениям относятся полисахариды, находящиеся в клетках в виде гликогена. Секреторные включения, содержащие биологически активные вещества, накапливаются в железистых клетках. Включения мотуг быть пигментными, попавшими в организм (в клетки) извне (красители, пьтевые частицы) или образовавшимися в самом организме в результате его жизнедеятельности (гемоглобин, меланин, липофусцин и др.).
Микроворсинки могут рассматриваться двояко: - с одной стороны - это органеллы специального назначения; - с другой стороны - это одна из форм специализации цитолеммы на апикальном полюсе клетки. Расположение - наиболее типичны для эпителиальных тканей (редко - в клетках соединительной ткани) и расположены на апикальном полюсе эпителиоцита. Количество у одной клетки - варьируют в зависимости от функциональной активности клетки, примерно 2000 одного эпителиоцита.
Разновидности микроворсинок: 1. микроворсинки щеточной каемки (более крупные, разной длины); 2. микроворсинки исчерченной каемки (более мелкие, одинаковой длины); 3. особая разновидность - стереоцилии (очень крупные, разветвленные, неподвижные микроворсинки, выполняющие рецепторную функцию, встречаются - (1) в сенсорном эпителии внутреннего уха, (2) в канале придатка мужской половой системы). Функции микроворсинок: 1. увеличивают апикальную поверхность клеток (например - для оптимального всасывания в кишечнике, почечных канальцах и др.; увеличения "рецепторного поля"). 2. создают оптимальные условия для протекания примембранных процессов (например - мембранного пищеварения, рецепции (например, рецепторные клетки вкусовых почек)).
Реснички могут рассматриваться двояко: 1. с одной стороны - это органеллы специального назначения эпителиоцитов (реже -клеток нервной ткани); 2. с другой стороны - это одна из форм специализации цитолеммы на апикальном полюсе клетки.
Расположение - расположены на апикальном полюсе клетки и типичны для эпителиальных тканей. Эпителий с ресничками называется мерцательным (примеры - респираторный эпителий; эпителий яйцевода).
Также могут встречаться в нервной ткани: (1) эпендимоциты, (2) чувствительные нейроны органа зрения и обоняния. Количество - примерно 250 у одной клетки.
Разновидности ресничек: 1. жгутик - большая одиночная ресничка (" в 10-20 раз длиннее реснички) - у сперматозоидов; 2. реснички, участвующие в рецепции: - палочки и колбочки в чувствительных нейронах сетчатки, - обонятельные реснички на обонятельных булавах в чувствительных нейронах обонятельного эпителия, - киноцилия - в рецепторных клетках вестибулярного аппарата.
Функции ресничек: 1. движение - если клетка встроена в эпителиальный пласт, продвигают секрет, частицы по по-верхности эпителия; если клетка свободная (простейшие, сперматозоид) перемещают клетку. 2. участие в рецепции.
Отличия от микроворсинок:
У ресничек: 1. всегда большие размеры (примерно в 5-7 раз длиннее); 2. "скелет" составляют не микрофиламенты, а микротрубочки (более толстые, из несократимого белка тубулина); 3. есть базальное тельце в основании; 4. никогда не ветвятся; 5. выполняют совершенно другие функции.
Жгутики представляет собой тонкий вырост на поверхности клетки, одетый трёхслойной клеточной мембраной и содержащий аксонему — совокупность микротрубочек. Жгутик осуществляет движение по типу ундуляции, совершая 10—40 об/сек.
8.Цитоскелет состоит: (филаментов), называемых цитоскелетом. В зависимости от диаметра филаменты разделяются на три группы: микрофиламенты (6-8 нм), промежуточные волокна(около 10 нм) и микротрубочки (около 25 нм). Все эти волокна представляют собой полимеры, состоящие из субъединиц особых глобулярных белков. Микрофиламенты (актиновые нити) состоят из актина — белка. Белки промежуточных волокон.Структурными элементами промежуточных волокон являются белки, принадлежащие к пяти родственным семействам и проявляющие высокую степень клеточной специфичности. Типичными представителями этих белков являются цитокератины, десмин, виментин, кислый фибриллярный глиапротеин [КФГП (GFAP)] инейрофиламент. Микротрубочки — это структуры, в которых 13 протофиламентов, состоящих из гетеродимеров α- и β-тубулина, уложены по окружности полого цилиндра. Внешний диаметр цилиндра около 25 нм, внутренний — около 15.
Один из концов микротрубочки, называемый плюс-концом, постоянно присоединяет к себе свободный тубулин. От противоположного конца — минус-конца — тубулиновые единицы отщепляются.
В образовании микротрубочки выделяют три фазы:
замедленная фаза, или нуклеация. Это этап зарождения микротрубочки, когда молекулы тубулина начинают соединяться в более крупные образования. Такое соединение происходит медленнее, чем присоединение тубулина к уже собранной микротрубочке, поэтому фаза и называется замедленной;
фаза полимеризации, или элонгация. Если концентрация свободного тубулина высока, его полимеризация происходит быстрее, чем деполимеризация на минус-конце, за счет чего микротрубочка удлиняется. По мере её роста концентрация тубулина падает до критической и скорость роста замедляется вплоть до вступления в следующую фазу;
фаза стабильного состояния. Деполимеризация уравновешивает полимеризацию, и рост микротрубочки останавливается.
Гиалоплазма (от греч. hyalinos — прозрачный) — основная плазма, или матрикс цитоплазмы, представляет собой очень важную часть клетки, ее истинную внутреннюю среду.
Гиалоплазма является сложной коллоидной системой, включающей в себя различные биополимеры, такие как белки, нуклеиновые кислоты, полисахариды. Эта система способна переходить из золеобразного (жидкого) состояния в гелеобразное и обратно. В упорядоченной многокомпонентной системе гиалоплазмы отдельные зоны могут менять свое агрегатное состояние в зависимости от условий или от функциональной задачи; в гиалоплазме могут возникать и распадаться различные комплексы белковых молекул. В состав гиалоплазмы входят главным образом различные глобулярные белки. Они составляют 20—25% общего содержания белков в эукариотической клетке. К важнейшим ферментам гиалоплазмы относятся ферменты метаболизма сахаров, азотистых оснований, аминокислот, липидов и других важных соединений. В гиалоплазме располагаются ферменты активации аминокислот при синтезе белков, транспортные (трансферные) РНК (тРНК). В гиалоплазме при участии рибосом и полирибосом (полисом) происходит синтез белков, необходимых для собственно клеточных нужд, для поддержания и обеспечения жизни данной клетки. Включения
Включения цитоплазмы (inclusiones cytoplasmicae) — необязательные компоненты клетки, возникающие и исчезающие в зависимости от метаболического состояния клеток.
Различают включения трофические, секреторные, экскреторные и пигментные. К трофическим включениям относятся капельки нейтральных жиров, которые могут накапливаться в гиалоплазме. В случае недостатка субстратов для жизнедеятельности клетки эти капельки могут резорбироваться. Другим видом включений резервного характера является гликоген — полисахарид, откладывающийся также в гиалоплазме (рис. 15). Отложение запасных белковых гранул обычно происходит в связи с активностью эндоплазматической сети. Так, запасы белка вителлина в яйцеклетках амфибии накапливаются в вакуолях эндоплазматической сети.
Секреторные включения — обычно округлые образования различных размеров, содержащие биологически активные вещества, образующиеся в клетках в процессе жизнедеятельности.
Экскреторные включения не содержат каких-либо ферментов или других активных веществ. Обычно это продукты метаболизма, подлежащие удалению из клетки.
Пигментные включения могут быть экзогенные (каротин, пылевые частицы, красители и др.) и эндогенные (гемоглобин, гемосидерин, билирубин, меланин, липофусцин). Наличие их в цитоплазме может изменять цвет ткани органа временно или постоянно. Нередко пигментация (или депигментация) ткани служит диагностическим признаком.
9. Ядро
Ядро (nucleus) клетки — система генетической детерминации и регуляции белкового синтеза.
Ядро обеспечивает две группы общих функций: одну, связанную собственно с хранением и передачей генетической информации, другую — с ее реализацией, с обеспечением синтеза белка.
Хранение и поддержание наследственной информации в виде неизменной структуры ДНК связаны с наличием так называемых репарационных ферментов, ликвидирующих спонтанные повреждения молекул ДНК. В ядре происходит воспроизведение или редупликация молекул ДНК, что дает возможность при митозе двум дочерним клеткам получить совершенно одинаковые в качественном и количественном отношении объемы генетической информации.
Другой группой клеточных процессов, обеспечиваемых активностью ядра, является создание собственно аппарата белкового синтеза (рис. 16). Это не только синтез, транскрипция на молекулах ДНК разных информационных РНК, но и транскрипция всех видов транспортных и рибосомных РНК. В ядре происходит также образование субъединиц рибосом путем комплексирования синтезированных в ядрышке рибосомных РНК с рибосомными белками, которые синтезируются в цитоплазме и переносятся в ядро.
Таким образом, ядро является не только вместилищем генетического материала, но и местом, где этот материал функционирует и воспроизводится. Вот почему выпадание или нарушение любой из перечисленных выше функций гибельно для клетки в целом. Все это указывает на ведущее значение ядерных структур в процессах синтеза нуклеиновых кислот и белков.
Структура и химический состав клеточного ядра
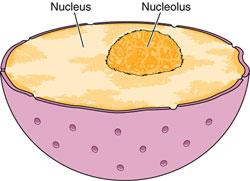
Ядро неделящейся, интерфазной клетки обычно одно на клетку (хотя встречаются и многоядерные клетки). Ядро состоит из хроматина, ядрышка, кариоплазмы (нуклеоплазмы) и ядерной оболочки, отделяющей его от цитоплазмы (рис. 17).
Хроматин
При наблюдении живых или фиксированных клеток внутри ядра выявляются зоны плотного вещества, которые хорошо воспринимают разные красители, особенно основные. Благодаря такой способности хорошо окрашиваться этот компонент ядра и получил название «хроматин» (от греч. chroma — цвет, краска). В состав хроматина входит ДНК в комплексе с белком. Такими же свойствами обладают и хромосомы, которые отчетливо видны во время митотического деления клеток. В неделящихся (интерфазных) клетках хроматин, выявляемый в световом микроскопе, может более или менее равномерно заполнять объем ядра или же располагаться отдельными глыбками.
Хроматин интерфазных ядер представляет собой хромосомы, которые, однако, теряют в это время свою компактную форму, разрыхляются, деконденсируются. Степень такой деконденсации хромосом может быть различной. Зоны полной деконденсации их участков морфологи называют эухроматином (euchromatinum). При неполном разрыхлении хромосом в интерфазном ядре видны участки конденсированного хроматина, иногда называемого гетерохроматином (heterochromatinum). Степень деконденсации хромосомного материала — хроматина в интерфазе может отражать функциональную нагрузку этой структуры. Чем «диффузнее» распределен хроматин в интерфазном ядре (т.е. чем больше эухроматина), тем интенсивнее в нем синтетические процессы. В ядрах, кроме хроматиновых участков и матрикса, встречаются перихроматиновые фибриллы, перихроматиновые и интерхроматиновые гранулы. Они содержат РНК и встречаются практически во всех активных ядрах, представляют собой информационные РНК, связанные с белками, — рибонуклеопротеиды (информосомы). Матрицами для синтеза этих РНК являются разные гены, разбросанные по деконденсированным участкам хромосомных (точнее, хроматиновых) фибрилл.
Ядрышко
Практически во всех живых клетках эукариотических организмов в ядре видно одно или несколько обычно округлой формы телец величиной 1—5 мкм, сильно преломляющих свет — это ядрышко, или нуклеола (nucleolus). К общим свойствам ядрышка относится способность хорошо окрашиваться различными красителями, особенно основными. Такая базофилия определяется тем, что ядрышки богаты РНК. Ядрышко — самая плотная структура ядра — является производным хромосомы, одним из ее локусов с наиболее высокой концентрацией и активностью синтеза РНК в интерфазе. Оно не является самостоятельной структурой или органеллой.
В настоящее время известно, что ядрышко — это место образования рибосомных РНК (рРНК) и рибосом, на которых происходит синтез полипептидных цепей уже в цитоплазме.
Образование ядрышек и их число связаны с активностью и числом определенных участков хромосом — ядрышковых организаторов, которые расположены большей частью в зонах вторичных перетяжек; количество ядрышек в клетках данного типа может изменяться за счет слияния ядрышек или за счет изменения числа хромосом с ядрышковыми организаторами. При исследовании фиксированных клеток вокруг ядрышка всегда выявляется зона конденсированного хроматина, часто отождествляемая с хроматином ядрышкового организатора. Этот околоядрышковый хроматин, по данным электронной микроскопии, представляет собой интегральную часть сложной структуры ядрышка. ДНК ядрышкового организатора представлена множественными (несколько сотен) копиями генов рРНК: на каждом из этих генов синтезируется высокомолекулярный предшественник РНК, который превращается в более короткие молекулы РНК, входящие в состав субъединиц рибосомы.
Схему участия ядрышек в синтезе цитоплазматических белков можно представить следующим образом: на ДНК ядрышкового организатора образуется предшественник рРНК, который в зоне ядрышка одевается белком, здесь происходит сборка рибонуклеопротеидных частиц — субъединиц рибосом; субъединицы, выходя из ядрышка в цитоплазму, участвуют в процессе синтеза белка.
Ядрышко неоднородно по своему строению: в световом микроскопе можно видеть его тонковолокнистую организацию. В электронном микроскопе выявляются два основных компонента: гранулярный и фибриллярный. Диаметр гранул около 15—20 нм, толщина фибрилл — 6—8 нм.
Фибриллярный компонент может быть сосредоточен в виде центральной части ядрышка, а гранулярный — по периферии. Часто гранулярный компонент образует нитчатые структуры — нуклеолонемы толщиной около 0,2 мкм. Фибриллярный компонент ядрышек представляет собой рибонуклеопротеидные тяжи предшественников рибосом, а гранулы — созревающие субъединицы рибосом. В зоне фибрилл можно выявить участки ДНК ядрышковых организаторов.
11. Митоз. Митоз-основной способ деления соматических (неполовых) клеток, при котором образуются 2 новые клетки с таким же набором хромосом, как и у материнской.Митоз представляет собой непрерывный процесс, который условно делят на 4 фазы:профазу,метафазу,анафазу,телофазу. Профаза. В начале профазы удвоенные тонкие нити хромосом лежат свободно внутри ядра. Затем начинается и постепенно нарастает их спирализация, в результате чего хромосомы принимают типичную двухроматидную структуру ядрышко постепенно исчезает, ядерная оболочка распадается. Начинается сборка веретена деления, хромосомы направляются к центру клетки. Метафаза. Хромосомы распологаются так, что их центомеры лежат в плоскости экватора клетки. Заканчивается формирование веретена деления клетки, нити которого направлены от одного полюса к другомы. Одни нити прикрепляются к центромерам хромосом другие проходят от полюса к полюсу, не прикрепляясь к хромосомам. Когда все хромосомы расположаться в эквоториальной плоскости, метафаза завершается. Анафаза. В этой фазе происходит разделение каждой хромосомы на хроматиды, которые становятся дочерними хромосомами. Нити веретена укорачиваются и тянут за собой хроматиды так, что они двигаются центромерами вперед, а их концы разъединяются последними. Телофаза. Обособление двух образовавшихся групп хромосом завершается, при этом вокруг каждой из них формируется ядерная оболочка. Хромосомы деспирализируются, удлиняются, становятся невидимыми. Нити веретена разрушаются, формируется ядрышко. Происходит образование клеточной стенки, которая делит цитоплазму пополам. После завершения деления молодые клетки, похожие на материнскую, но имеющие общие размеры, вступают в интерфазу.(интерфаза- часть клеточного цикла между двумя последовательными митозами,продолжительность интерфазы около 90% времени всего клеточного цикла.) В результате митоза образуются две клетки, ядра которых содержат столько же хромосом, сколько их было в родительском ядре. В результате мейоза образуются половые клетки,несущие одинарный набор хромосом. Клеточная теория — одно из общепризнанных биологических обобщений, утверждающих единство принципа строения и развития мира растений и мира животных, в котором клетка рассматривается в качестве общего структурного элемента растительных и животных организмов. Основные положения современной клеточной теории
Клетка — единица строения, жизнедеятельности, роста и развития живых организмов, вне клетки жизни нет.
Клетка — единая система, состоящая из множества закономерно связанных друг с другом элементов, представляющих собой определённое целостное образование.
Ядро − главная составная часть клетки (эукариот).
Новые клетки образуются только в результате деления исходных клеток.
Клетки многоклеточных организмов образуют ткани, ткани образуют органы. Жизнь организма в целом обусловлена взаимодействием составляющих его клеток. Значение клеточной теории.
Cтало ясно, что клетка — важнейшая составляющая часть живых организмов, их главный морфофизиологический компонент. Клетка — это основа многоклеточного организма, место протекания биохимических и физиологических процессов в организме. На клеточном уровне в конечном итоге происходят все биологические процессы. Клеточная теория позволила сделать вывод о сходстве химического состава всех клеток, общем плане их строения, что подтверждает филогенетическое единство всего живого мира.
12.Половые клетки
(синоним гаметы)
специализированные клетки, обладающие гаплоидным набором хромосом и обеспечивающие при половом размножении передачу наследственной информации от родителей к потомству. П. к. происходят от диплоидных клеток в результате редукции числа хромосом при эволюции полового процесса. У большинства многоклеточных организмов первичные П. к. (гоноциты) обособляются вследствие первых делений в начале эмбриогенеза из экто- или энтодермы, а затем различными путями перемещаются в формирующиеся половые железы. У особей мужского пола в половых железах образуются мужские П. к., или сперматозоиды, а у женского —яйцеклетки. Организм с истинным Гермафродитизмом продуцирует оба типа П. к. Процесс развития П. к. рассматривается как начальный этап Онтогенеза и называется гаметогенезом, образование сперматозоидов — сперматогенезом, а яйцеклеток — оогенезом. Зрелые половые клетки возникают в процессе гаметогенеза, который условно подразделяют на 4 периода — размножение, рост, созревание и формирование (рис. 1). В периоде размножения диплоидные клетки сперматогонии и оогонии (предшественники гамет) несколько раз делятся митотически, обеспечивая нарастание числа будущих П. к. Интенсивное размножение особенно характерно для сперматогонии. В периоде роста происходит увеличение размеров клеток, сопровождающееся накоплением в них питательных веществ, РНК, ряда структурных белков. Значительный рост клеток ярко выражен в оогенезе. Основным содержанием периода созревания является мейоз, в результате которого из каждой диплоидной клетки-предшественницы образуется 4 клетки с гаплоидным набором хромосом. При сперматогенезе эти клетки одинаковы по размеру и позже становятся сперматозоидами, а при оогенезе мейоз обеспечивает неравномерное деление цитоплазмы. В результате лишь одна гаплоидная клетка из четырех становится яйцеклеткой, способной к оплодотворению, а три другие представляют собой редукционные тельца, содержащие избыток хроматина и в конечном счете погибающие. Кроме обеспечения гаплоидности мейоз приводит также к возникновению качественного многообразия половых клеток. В профазе первого мейотического деления гомологичные хромосомы отцовского и материнского происхождения, спирализуясь, сближаются попарно соответствующими друг другу участками (так называемая конъюгация), образуя биваленты. При этом отдельные хроматиды переплетаются между собой и могут разрываться в аналогичных участках. В процессе восстановления целости хроматид гомологичные хромосомы способны обмениваться соответствующими участками. Этот процесс называется кроссинговером. В анафазе первого мейотического деления происходит независимое расхождение материнских и отцовских хромосом к полюсам клетки (Клетка), вследствие чего в гаплоидном наборе будущих гамет возникают разные сочетания материнских и отцовских хромосом. Последний период гаметогенеза (период формирования) наблюдается только при сперматогенезе, во время которого гаплоидные клетки — сперматиды — приобретают особенности строения, характерные для зрелых сперматозоидов. Сперматогенез у самцов млекопитающих протекает постоянно на протяжении всего периода половой зрелости и мало зависит от действия гормональных факторов. В отличие от этого этапы оогенеза не только растянуты во времени, но и гормонально зависимы. Запас ооцитов, содержащихся при рождении в яичниках самок, в дальнейшем не пополняется. Данная закономерность присуща и гаметогенезу человека. Так, в яичнике новорожденной девочки содержится около 2 млн. ооцитов, из них достигают зрелости и претерпевают овуляцию только приблизительно 400. Первое мейотическое деление в ооците происходит в момент овуляции, второе — уже после проникновения в него сперматозоида.
Зрелая яйцеклетка имеет неоднородную биохимическую организацию. На анимальном ее полюсе увеличена концентрация гликогена и РНК, по экватору располагается аскорбиновая кислота, желток собирается в основном на вегетативном полюсе. Такая неоднородность цитоплазмы называется ооплазматической сегрегацией. Она имеет большое значение для детерминации и последующей дифференцировки Зародыша на стадиях дробления и гаструляции. На характер дробления влияют также количество и распределение желтка в яйцеклетке. Снаружи яйцеклетка покрыта кроме цитоплазматической мембраны также и дополнительными оболочками. Так, яйцеклетка млекопитающих и человека имеет блестящую оболочку — zona pellucida, окруженную фолликулярными клетками, участвующими в ее метаболизме (рис. 2). Для оплодотворения необходимо сближение мужских и женских П. к., которое достигается благодаря осеменению. У большинства водных животных осеменение наружное, у наземных животных — внутреннее, при котором сперматозоиды попадают непосредственно в половые пути самки. Встрече мужских и женских гамет (рис. 3) и предохранению яйцеклетки от внедрения в нее избыточного количества сперматозоидов способствуют выделяющиеся П. к. специфические вещества — гино- и андрогамоны. В момент контакта половых клеток происходит выделение ферментов акросомы сперматозоида и растворение участка яйцевой оболочки. При этом объединяется содержимое обеих гамет и образуется единая клетка — зигота. У некоторых организмов ядра сперматозоида и яйцеклетки (пронуклеусы) сливаются вскоре после оплодотворения. У млекопитающих и человека их объединение происходит в метафазе первого деления — дробления.
Биологическое значение оплодотворения заключается в восстановлении в зиготе диплоидности хромосомного набора, характерного для данного вида организмов, и в возникновении ее генетической уникальности в результате объединения наследственного материала хромосом родительских особей, претерпевшего предварительную рекомбинацию в процессе мейоза. Кроме того, проникновение сперматозоида в яйцеклетку обеспечивает активацию в ней различных синтетических процессов и начало дробления.
13. Яйцеклетки образуются в результате овогенеза. После оплодотворения из оплодотворенной яйцеклетки (зиготы) развивается эмбрион. При партеногенезе эмбрион, а затем новый организм развивается из неоплодотворенной яйцеклетки. По количеству желтка
Полилецитальные — содержат большое количество желтка (членистоногие, рептилии, птицы, рыбы, кроме осетровых).
Мезолецитальные — содержат среднее количество желтка (осетровые рыбы, амфибии).
Олиголецитальные — содержат мало желтка (моллюски, иглокожие).
Алецитальные — не содержат желтка (млекопитающие, некоторые паразитические перепончатокрылые). Телолецитальные — желток смещён к вегетативному полюсу яйцеклетки. Противоположный полюс называется анимальным. Сюда относятся некоторые полилецитальные (рыбы, кроме осетровых, рептилии, птицы) и все мезолецитальные яйца (осетровые рыбы, амфибии).
Гомо (изо)- лецитальные — желток распределён равномерно. Сюда относятся олиголецитальные ядра (моллюски, иглокожие).
Центролецитальные — желток расположен в центре яйцеклетки. Сюда относятся некоторые полилецитальные яйца (членистоногие). Это совершенно особый тип яиц. Анимально-вегетативная полярность этих яиц не выражена, так как место выделения редукционных телец может быть различным. Вместо анимального и вегетативного полюсов у этих яиц говорят о переднем и заднем полюсах. В центре яйца расположено ядро, а по периферии — ободок свободной от желтка цитоплазмы. Оба этих района — центр и периферия яйца — связаны тонкими цитоплазматическими мостиками, а всё промежуточное пространство заполнено желтком. 1. Полное дробление. К полному, тотальному дроблению относятся те случаи, когда в процессе дробящего деления делится вся оплодотворенная яйцевая клетка и борозды дробления распространяются по всей ее поверхности. По этому типу дробятся яйцевые клетки голобластического вида. В зависимости от содержания в ооплазме большего или меньшего количества желтка, а также в зависимости от его распределения в ооплазме, при дроблении возникают бластомеры либо сравнительно одинаковой величины (полное равномерное, эквальное, или адэквальное дробление), либо бластомеры различной величины, а именно более крупные в области с большим содержанием желтка и менее крупные в том месте, где желтка меньше (полное неравномерное, инэквальное дробление). Более крупные бластомеры называются макромерами, менее крупные — микромерами.
Полное эквальное, или адэквальное, дробление свойственно олиголецитальным, изолецитальным яйцеклеткам (ланцетник, высшие млекопитающие и человек); по полному инэквальному типу дробятся мезолецитальные яйцевые клетки анизолецитального и умеренно телолецитального вида (некоторые низшие рыбы и земноводные).
2. Частичное, парциальное, дробление. По частичному типу дробятся яйцевые клетки, содержащие значительное количество желтка (полилецитальные яйцеклетки), у которых из-за их больших размеров борозды дробления при клеточном делении проникают только в область анимального полюса, где находится клеточное ядро и где слой ооплазмы содержит меньше желточных гранул (высшие рыбы, пресмыкающиеся, птицы и некоторые низшие млекопитающие, яйцеродные).
При таком дроблении на анимальном полюсе сравнительно крупного яйца дробится только круглое поле (диск), в то время как остаток яйцевой клетки (желточный шар) остается не раздробленным (парциальное дисковидное дробление). У насекомых их полилецитальные центролецитальные яйцеклетки хотя и дробятся по всей поверхности, но центр клетки, содержащий большое количество желтка, остается не раздробленным (парциальное поверхностное дробление).
В приведенном рисунке показаны отдельные виды яйцевых клеток в зависимости от содержания и распределения желтка в ооплазме, а также в зависимости от соответствующего типа дробления.
14. Понятие и этапы эмбриогенеза.
Эмбриогенез (греч. embryon - зародыш, genesis - развитие) - ранний период индивидуального развития организма от момента оплодотворения (зачатия) до рождения, является начальным этапом онтогенеза (греч. ontos - существо, genesis - развитие), процесса индивидуального развития организма от зачатия до смерти. Развитие любого организма начинается в результате слияния двух половых клеток (гамет), мужской и женской. Все клетки тела, несмотря на различия в строении и выполняемых функциях, объединяет одно - единая генетическая информация, хранящаяся в ядре каждой клетки, единый двойной набор хромосом (кроме узкоспециализированных клеток крови - эритроцитов, которые не имеют ядра). То есть, все соматические (сома - тело) клетки диплоидны и содержат двойной набор хромосом - 2 n, и лишь половые клетки (гаметы), формирующиеся в специализированных половых железах (семенниках и яичниках), содержат одинарный набор хромосом - 1 n. При слиянии половых клеток образуется клетка - зигота, в которой восстанавливается двойной набор хромосом. Напомним, что в ядре клетки человека содержится 46 хромосом, соответственно половые клетки имеют 23 хромосомы Образовавшаяся зигота начинает делиться. I этап деления зиготы называется дроблением, в результате которого образуется многоклеточная структура морула (тутовая ягода). Цитоплазма распределяется между клетками неравномерно, клетки нижней половины морулы крупнее, чем верхней. По объему морула сравнима с объемом зиготы. На II этапе деления, в результате перераспределения клеток, образуется однослойный зародыш - бластула, состоящий из одного слоя клеток и полости (бластоцель). Клетки бластулы различаются между собой по размерам. На III этапе, клетки нижнего полюса как бы впячиваются (инвагинируют) вовнутрь, и образуется двухслойный зародыш - гаструла, состоящий из наружного слоя клеток - эктодермы и внутреннего слоя клеток - энтодермы. Очень скоро, между I и II слоями клеток формируется, в результате деления клеток, еще один слой клеток, средний - мезодерма, и зародыш становится трехслойным. На этом завершается стадия гаструлы. Из этих трех слоев клеток (их называют зародышевыми слоями) формируются ткани и органы будущего организма. Из эктодермы развивается покровная и нервная ткань, из мезодермы - скелет, мышцы, кровеносная система, половые органы, органы выделения, из энтодермы - органы дыхания, питания, печень, поджелудочная железа. Многие органы формируются из нескольких зародышевых слоев.
Эмбриогенез или эмбриональное развитие – это развитие организма от оплодотворения яйцеклетки до рождения или вылупления. Рассмотрим стадии эмбрионального развития.
Дробление – это последовательных митотических делений зиготы и следующих поколений клеток, в результате которых образуются все более мелкие клетки, все вместе не превышающие размерами исходную зиготу. При дроблении появляющиеся поколения клеток не отличаются друг от друга ни строением, ни выполняемыми функциями. Про такие клетки говорят, что они не дифференцированы. В зависимости от особенностей строения зиготы дробление протекает по-разному. Можно выделить три типа дробления.
Неполное дробление наблюдается у животных, зиготы которых имеют огромный запас питательных веществ, например, у птиц и пресмыкающихся. Хорошо всем известный желток яйца курицы есть ничто иное, как зигота (или яйцеклетка). В такой зиготе ядро плавает на ее поверхности. Эта часть зиготы называется анимальным полюсом . На противоположном конце от ядра находится так называемый вегетативный полюс . Таким образом, можно сказать, что у при неполном дроблении деление протекает только на анимальном полюсе зиготы.
Полное неравномерное дробление наблюдается у животных, зиготы которых имеют много питательных веществ (но меньше, чем у птиц и пресмыкающихся) например у рыб и земноводных. Зигота этих животных (икринка) делится полностью, но уже после нескольких делений частота делений клеток анимального полюса становится заметно выше, чем у.
полюса.
Полное равномерное дробление встречается у животных, зиготы которых имеют относительно малое количество питательных веществ цитоплазмы. К ним относятся млекопитающие и головохордовые (ланцетник). В зиготе слабо выражены анимальный и вегетативный полюса, поэтому зигота делится полностью и дальнейшее деление клеток идет почти с одинаковой интенсивностью, как на анимальном, так и на вегетативном полюсе.
Рассмотрим более подробно дробление зиготы ланцетника. Первые два деления зиготы проходят меридионально, то есть в вертикальной плоскости. Следующее деление происходит в широтном направлении. Затем все последующие деления представляют строгое чередование делений клеток меридионально и широтно. Так постепенно количество клеток растет, их скопление имеет шаровидную форму и называется бластула . Бластула представляет собой полый шар, размеры которого не превосходят исходную зиготу. Полость внутри называется бластопор . Стенки бластулы образованы только одним слоем клеток. 5. Оплодотворение
Оплодотворение - процесс слияния мужской и женской гамет, приводящее к образованию зиготы. При оплодотворении взаимодействуют мужская и женская гаплоидные гаметы, при этом сливаются их ядра (пронуклеусы), объединяются хромосомы, и возникает первая диплоидная клетка нового организма - зигота. Начало оплодотворения - момент слияния мембран сперматозоида и яйцеклетки, окончание оплодотворения - момент объединения материала мужского и женского пронуклеусов.
Оплодотворение происходит в дистальном отделе маточной трубы и проходит 3 стадии.
I стадия - дистантное взаимодействие, включает в себя 3 механизма:
хемотаксис - направленное движение сперматозидов навстречу к яйцеклетке (гинигамоны 1,2);
реотаксис - движение сперматозоидов в половых путях против тока жидкости;
капацитация - усиление двигательной активности сперматозоидов, под воздействием факторов женского организма (рН, слизь и другие).
II стадия - контактное взаимодействие, за 1,5-2 ч сперматозоиды приближаются к яйцеклетке, окружают ее и приводят к вращательным движениям, со скоростью 4 оборота в минуту. Одновременно из акросомы сперматозоидов выделяются сперматозилины, которые разрыхляют оболочки яйцеклетки. В том месте, где оболочка яйцеклетки истончается максимально, происходит оплодотворение, оволемма выпячивается и головка сперматозоида проникает в цитоплазму яйцеклетки, занося с собой центриоли, но оставляя снаружи хвостик.
III стадия - проникновение, самый активный сперматозоид проникает головкой в яйцеклетку, сразу после этого в цитоплазме яйцеклетки образуется оболочка оплодотворения, которая препятствует полиспермии. Затем происходит слияние мужского и женского пронуклеусов, этот процесс носит название синкарион. Этот процесс (сингамия) и есть собственно оплодотворение, появляется диплоидная зигота (новый организм, пока одноклеточный). Условия, необходимые для оплодотворения:
концентрация сперматозоидов в эякуляте, не менее 60 млн в 1 мл;
проходимость женских половых путей;
нормальная температура тела женщины;
слабощелочная среда в женских половых путях.
Дробление - это последовательно протекающий митоз, без роста образовавшихся клеток до размеров исходной. При дроблении происходит относительно быстрое увеличение количества клеток (бластомеры). Дробление идет до тех пор, пока не восстановится соотношение объема ядра к объему цитоплазмы, характерное для данного вида. Количество бластомеров увеличивается от 2 до примерно 12-16 к третьим суткам после оплодотворения, когда концептус достигает стадии морулы и выходит в полость матки из маточных труб.
Различают дробление:
полное, неполное;
равномерное, неравномерное;
синхронное, асинхронное.
У человека дробление полное, асинхронное, неравномерное. В результате первого деления образуются 2 бластомера, темный и светлый, светлые делятся быстро и обволакивают зиготу снаружи - трофобласт, а темные находятся внутри и делятся медленно: эмбриобласт. Дробление зиготы у человека прекращается на стадии 107 бластомеров.
Имплантация состоит из 2-х этапов:
адгезия - прилипание;
инвазия - погружение.
15. Гаструляция - это период эбрионального развития, при котором происходит размножение, рост и перемещение отдельных клеток и обширных клеточных пластов. Главным отличием гаструляции от предшествующих периодов эмбриогенеза является приобретение клетками способности к направленным морфогенетическим перемещениям, которые приводят к глубокой перестройке зародыша. Если результатом дробления было формирование многоклеточности, то гаструляция приводит к образованию многослойного зародыша. Морфогенетические перемещения клеток сильно варьируют в разных классах животных.Рассмотрим основные типы гаструляции: 1) Инвагинация - это впячивание одной стенки бластулы внутрь бластоцеля.Такой способ гаструляции характерен для ланцентика. 2) Эпиболия - обрастание мелкими быстро делящимися клетками крупных, медленно делящихся клеток, перегруженных желтком и поэтому не обнаруживающих способности к перемещениям. Этот способ гаструляции наблюдается у амфибий. 3) Деламинация - расщепление бластодермы на два слоя. При этом способе гаструляции клеточные смещения почти отсутствуют. Элементы деламинации встречаются при гаструляции рыб, птиц, млекопитающих. 4) Иммиграция - активное выселение части клеток стенки бластулы внутрь бластоцеля. Когда клетки выселяются лишь с одного полюса бластулы, говорят об униполярной иммграции, с двух полюсов - биполярной, а при выселении клеток со всей поверхности зародыша - о мультиполярной. Это также довольно распространенный способ гаструляции и встречается у многих позвоночных (рыбы, птицы, млекопитающие). Встречаются смешанные типы гаструляции. Вообще, рассмотренные типы гаструляции в известной мере условны и в большинстве случаев правильнее говорить, что какой-либо из типов является доминирующим в характере морфогенетических движений, и в процессе гаструляции у многих животных могут иметь место одновремено несколько из этих типов. Механизмы гаструляции. Как ни разнообразны типы гаструляции, существуют общие изменения на клеточном уровне, которые приводят к морфогенетическим перемещениям. В основе большинства формообразовательных процессов лежат клеточные деления, возникновение механических напряжений в пласте, а затем чередование поляризации (растяжения) и сокращения клеток. Поляризация клетки - это перераспределение органелл с быстрой активацией синтеза актина, сборкой пучков микротрубочек и вытягиванием клетки в направлении предстоящего движения. При этом формируется так называемая колбовидная клетка. Характернымявляется то, что такая поляризация затрагивает не одну клетку, а целый клеточный пласт: т.е. поляризация одной клетки побуждает соседнюю к такому же превращению. Этот процесс осуществляется лишь при наличии клеточных контактов и поэтому носит название контактной клеточной поляризации. Клетки не могут оставаться в состоянии поляризации неограниченно долго: через определенное время происходит сокращение - такая деформация поляризованной клетки, которая уменьшает отношение ее поверхности к объему. Этот процесс осуществляется сократительным аппаратом клетки - микрофиламентами. В ходе последовательных сокращений пласт изгибается, происходит элементарное морфогенетическое перемещение. Таким образом,возникающие в определенном регионе в связи с особымит внешними условиями и воздействиями деления клеток создают механические напряжения в пласте. Эти напряжения ведут к возникновению поляризации клеток, которая несет "информацию" о направлении будущего перемещения, а ее реализация (т.е. собственно перемещение) происходит во время сокращения. Клеточные процессы, лежащие в основе формообразующих процессов раннего развития, их координация во времени и простарнстве, возможные иные причины удивительно сложных и упорядоченных механизмов гаструляции являются и на сегодняшний день по большей части неясными. Для удобства рассмотрения хода гаструляции и ее результатов у позвоночных, весь процесс условно разделен на два этапа: ранняя и поздняя гаструляция. В течение ранней гаструляции первоначально единственный слой клеток бластулы, реорганизуясь любым из приведенных выше способом, образует два слоя. Наружный слой клеток называют эктодермой, а внутренний - энтодермой. У низших позвоночных при этом формируется новая полость - гастроцель. Отверстие, ведущее наружу из гастроцеля, называют бластопором (первичным ртом), а его края - губами. Материал дорсальной губы бластопора у разных видов животных в дальнейшем преобразуется в хорду, а латеральных - в третий зародышевый листок - мезодерму. Поэтому для представления о морфогенетических перемещениях бластопор является достаточно важным ориентиром. Судьба бластопора у разных животных различна. У одних (первичноротые) бластопор, соответственно развиваясь и дифференцируясь, превращается в дефинитивный рот, у других (вторичноротые) - бластопор преобразуется в анальное отверстие. У высших позвоночных (птицы, млекопитающие) при иммиграции не формирует бластопор. Для ориентировки в будущих путях развития зародыша можно говорить лишь об аналоге бластопора. Таким образом, в результате ранней гаструляции формируется двухслойный зародыш и бластопор, а у млекопитающих кроме этого еще и некоторые внезародышевые органы. При поздней гаструляции образуется третий зародышевый листок - мезодерма, комплекс осквых органов и внезародышевые органы. Классическая эмбриология описывает два способа формирования мезодермы: энтероцельный и телобластический. При энтероцельном способе мезодерма формируется как совокупность клеток, отделившихся от первичной кишки, а при телобластическом - мезодерма образуется из клеток, располагавшихся на будущем заднем конце зародыша на границе экто- и энтодермы. Осевые органы - это хорда, нервная трубка и первичная кишка. Первой из материала дорсальной губы бластопора формируется хорда - плотный клеточный тяж, расположенный по средней линии зародышамежду экто- и энтодермой. Под ее влиянием в наружном зародышевом листке начинает формироваться нервная трубка. И в последнюю очередь энтодерма образует первичную кишку. Формирование нервной трубки непосредственно связано с нейруляцией - закладкой центральной нервной системы. Нейруляция - очень важный и интересный период в развитии зародыша не только потому, что происходит закладка сложнейшей системы, но и потому, что при формировании нервной трубки отмечается теснейшее взаимодействие между прилегающими структурами: эктодермой, хордой и мезодермой. Следует подчеркнуть, что одно из главных последствий морфогенетических перемещений состоит в том, что группы клеток, которые прежде могли быть значительно удалены друг от друга, настолько сближаются, что между ними становятся возможными взяимодействия,которые носят название индукционных. Нейруляция, в частности, формирование нервной трубки, является результатом таких индукционных взаимодействий.
16. Внезародышевые органы
Нервная пластинка, расположенная над хордой, превращается в нервную трубку. Мезодерма дифференцируется на сомиты и боковые пластинки. Полость, образующаяся между париетальным и висцеральным листками мезодермы, представляет зачаток целома. Из энтодермы формируется кишечная трубка, что, так же как у птиц, совершается одновременно с обособлением тела зародыша от желточного мешка.
Вместе с изменениями в зародышевом щитке, которые приводят к закладке органов, начинается образование внезародышевых органов: амниона, аллантоиса и серозной оболочки. На стадии закладки осевых органов весь зародыш покрыт трофобластом. Та часть трофобласта, которая находится над щитком, существует лишь короткое время, перед образованием зародышевых оболочек она рассасывается и исчезает. В результате этого края эктодермы щитка срастаются с остающейся частью трофобласта. Образуется сплошной эктодермальный слой, в котором зародышевая эктодерма переходит в эктодерму трофобласта. Одновременно с этим начинается разрастание энтодермы: она выходит из области зародышевого диска, обрастает с внутренней стороны трофобласт и образует энтодермальный мешок, наполненный жидкостью. Стенка бластодермического пузырька состоит теперь из трофобласта и внезародышевой энтодермы. Образовавшийся мешок называют желточным по аналогии с подобным органом у эмбрионов пресмыкающихся и птиц. Что касается млекопитающих, то у них желточный мешок желтка не содержит и, хотя образуется как наследие от развития яйцекладущих предков – пресмыкающихся, играет большую роль на ранних стадиях развития зародышей млекопитающих. Во внезародышевой мезодерме, врастающей между трофобластом и энтодермой желточного мешка, развиваются кровеносные сосуды. С помощью трофобласта сосуды желточного мешка связаны со слизистой оболочкой матки и через них осуществляется питание и дыхание зародыша. Кроме того, в стенке желточного мешка начинается кроветворение, которое позднее происходит в теле зародыша. Сейчас же после редукции участка трофобласта над щитком начинается обособление тела зародыша, который распластан на желточном мешке, как куриный зародыш на желтке. Появляются туловищная и амниотическая складки. По мере углубления туловищной складки тело зародыша все больше приподнимается над желточным мешком и отделяется от него. При этом энтодерма, расположенная, как у птиц, под зародышем, втягивается внутрь его и образует карманообразную складку. Последняя по мере отделения зародыша от мешка превращается в кишечную трубку, которая желточным протоком соединяется с желточным мешком. Развитие амниотической и туловищной складок у птиц сопровождается разрастанием внезародышевых экто- и мезодермы. У млекопитающих происходит иначе. К началу образования амниона внезародышевая эктодерма уже представлена трофобластом, поэтому развитие амниотической складки сопровождается разрастанием только внезародышевой мезодермы. При смыкании складок образуется амниотическая полость, в которой оказывается зародыш, и оболочка, соответствующая серозной у птиц. Эта оболочка окружает амнион вместе с зародышем и желточный мешок. С образованием амниотической полости тело зародыша связывается с внезародышевыми частями только при помощи пупочного канатика. В отличие от зародыша птиц у млекопитающих большое значение приобретает серозная оболочка. На начальных стадиях развития на гладкой поверхности трофобласта появляются выросты – первичные ворсинки, образованные только трофобластом. Затем в них врастает внезародышевая мезодерма серозной оболочки и сосуды аллантоиса; первичные ворсинки превращаются во вторичные. Образовавшаяся оболочка с ворсинками получила название хорион. Через нее происходит всасывание и проведение питательных веществ к зародышу. Развивающиеся ворсинки хориона увеличивают его всасывающую поверхность, что связано с ростом зародыша и его возрастающей потребностью в питательных веществах. Там, где хорион вступает в тесный контакт со слизистой оболочкой матки, образуется детское место, или плацента. Через этот важный орган осуществляется весь обмен веществ между зародышем и организмом матери. Одновременно с образованием амниотической полости развивается аллантоис. Он появляется в виде небольшого выроста на брюшной стенке заднего отдела кишки. Аллантоис у млекопитающих не разрастается так сильно, как у птиц. По выходе из кишки он располагается около желточного мешка и подрастает к хориону. В аллантоисе заключены пупочные сосуды, которые врастают в хорион и таким образом включаются в кровеносное русло материнского организма. С этого момента обмен веществ зародыша совершается по пупочным сосудам. В связи с тем, что питание зародыша происходит за счет материнского организма, значение некоторых внезародышевых органов у млекопитающих иное, чем у птиц и пресмыкающихся. Особенно большое значение приобретает хорион. Аллантоис служит только проводником пупочных сосудов, в то время как у птиц он является важным органом дыхания и выделения зародыша. Значение амниона не изменяется. Он значительно разрастается и настолько увеличивается, что в спинной части подходит к хориону, а в брюшной вплотную подрастает к пупочному канатику. Под давлением амниотической стенки аллантоис и желточный мешок сужаются. Последний, как уже отмечалось, играет роль в питании и дыхании зародыша только на ранних стадиях развития и поэтому довольно скоро редуцируется. Основную часть пупочного канатика составляют сосуды зародышевого кровообращения, для которых он служит проводником к материнскому организму. На более поздних стадиях развития амнион еще более увеличивается. Зародыш оказывается как бы подвешенным в нем на пупочном канатике. С внутренней стороны амниотическая полость выстлана эпителием, который покрывает пупочный канатик и переходит в кожный эпителий зародыша. Весь зародышевый пузырь, т.е. зародыш и его внезародышевые органы, покрыт хорионом.
17. Гаструляция у человека совершается в 2 этапа:
|
|
1 ЭТАП |
2 ЭТАП |
|
способ гаструляции |
деляминация |
миграция |
|
ОБРАЗУЮТСЯ |
||
|
зародышевые листки |
эктодерма, энтодерма |
мезодерма, мезенхима |
|
осевые органы |
|
хорда, нервная трубка, мезодерма, кишечная трубка |
|
провизорные органы |
амнион, желточный мешок, хорион (вторичные ворсинки) |
аллантоис, идет формирование плаценты - третичные ворсинки хориона |
18. ГАСТРУЛЯЦИЯ - это процесс образования зародышевых листков. Гаструляция у человека происходит в два этапа. Первый этап идет путем деляминации (расщепления), а второй - путем миграции.
В процессе первого этапа образуются два зародышевых листка (экто- и энтодерма), два провизорных органа (амнион и желточный мешок). Кроме того, непосредственно перед началом первого этапа происходит образование такого провизорного органа, как хорион. Формирование хориона - это второй этап в образовании плаценты.
Во время второго этапа образуются еще два зародышевых листка (мезодерма и мезенхима), провизорный орган - аллантоис и идет дальнейшее образование еще одного провизорного органа - плаценты: формируются третичные ворсинки хориона, которые в последующем соединяются с decidua basalis и формируют плаценту. Образуются осевые органы - хорда, нервная трубка, кишечная трубка, мезодерма.
первый этап гаструляции
Клетки эмбриобласта подразделяются на две части. Одна часть представляет собой пласт клеток, расположенных в один ряд. Эта группа клеток получает название зародышевого диска (щитка). Все остальные клетки эмбриобласта превращаются во внезародышевую мезенхиму.
Образование хориона. Внезародышевые мезенхима подрастает к трофобласту и получаются вторичные ворсинки хориона. Хорион - это трофобласт + внезародышевая мезенхима. Вторичные ворсинки хориона состоят из трофобласта и внезародышевой мезенхимы.
Образование эктодермы и энтодермы. Клетки зародышевого диска делятся в тангенциальной плоскости, то есть происходит его расщепление (деляминация) на два слоя. В результате этого зародышевый диск уже состоит из двух слоев клеток. Верхний слой клеток - это эктодерма (эпибласт), нижний слой - энтодерма (гипобласт).
Образование амниона. Над эктодермой скапливается жидкость, она отодвигает клетки внезародышевой мезенхимы. В результате образуется пузырек, заполненный жидкостью - амниотический пузырек. Клетки эктодермы разрастаются за пределы зародышевого диска и обрастают поверхность амниотического пузырька. Получается амнион. Стенка амниона состоит из внезародышевой эктодермы и внезародышевой мезенхимы. Внезародышевая эктодерма называется внезародышевой потому, что она находится за пределами зародышевого диска.
Образование желточного мешка. Под энтодермой скапливается жидкость, которая отодвигает внезародышевую мезенхиму. В результате этого образуется пузырек, заполненный жидкостью, называемый желточным пузырьком. Энтодерма разрастается за пределы зародышевого диска, обрастает поверхность желточного пузырька и получается желточный мешок. Его стенка состоит из внезародышевой энтодермы и внезародышевой мезенхимы. Внезародышевая энтодерма называется внезародышевой, так как она вышла за пределы зародышевого диска.
второй этап гаструляции
В эктодерме клетки начинают двигаться (мигрировать) с двух сторон от головного к каудальному (хвостовому) концу зародыша. В области каудального конца клеточные потоки встречаются и начинают двигаться кпереди. При движении клеток эктодермы в срединной части образуется нагромождение клеток, которое получает название первичной полоски. В средней части эктодермы движение клеток останавливается, и поэтому в передней части первичной этой полоски имеется еще большее нагромождение клеток, которое получает название первичного узелка.
Образование мезодермы происходит из клеток первичной полоски. Клетки первичной полоски, образовавшейся в эктодерме, прорастают в пространство между экто- и энтодермой и там разрастаются, образуя мезодерму.
Образование хорды происходит из клеток первичного узелка. Клетки первичного узелка, образующегося в эктодерме, прорастают в пространство между экто- и энтодермой, там разрастаются вперед и назад и образуют хорду.
Образование нервной трубки. В срединной части эктодермы образуется углубление - нервная пластинка. Нервная пластинка погружается под эктодерму, а края эктодермы смыкаются и образуется нервная трубка.
Образование кишечной трубки происходит в процессе формирования туловищной складки. Туловищная складка формируется путем подгибания всех имеющихся зародышевых листков под тело зародыша. В результате этого кишечная (зародышевая) энтодерма отделяется от желточного мешка (желточная энтодерма). Кишечная трубка остается связанной с желточным мешком только в одном небольшом участке - желточном стебельке (протоке). Часть желточного протока может остаться после рождения в виде Меккелева дивертикула подвздошной кишки).
Дифференцировка мезодермы и образование 4 зародышевого листка - мезенхимы. Сразу после своего образования мезодерма подразделяется на два главных отдела: сомиты - спинной отдел и сплахнотом - брюшной отдел. Между сомитами и сплахнотомом имеется еще один отдел - сегментная ножка, с помощью которой они соединяются. Сомиты разделяются на три части: дерматом, склеротом, миотом. Дерматом дает начало мезенхиме дерматома, из нее образуется дерма - собственно кожа. Миотом является источником поперечнополосатых мышц. Из склеротома образуется мезенхима склеротома, которая дает начало костям и хрящам. Сплахнотом делится на висцеральный и париетальный листки, между которыми находится полость тела - целом. Висцеральный и париетальный листки дают начало висцеральным и париетальным серозным оболочкам. Кроме того, из висцерального листка спланхнотома выселяются клетки, которые дают начало мезенхиме спланхнотома, из которой развивается соединительная ткань. Таким образом, мезенхима образуется из трех источников - висцерального листка спланхнотома, дерматома, склеротома. Мезенхима является четвертым зародышевым листком. Из мезенхимы развивается вся соединительная ткань. Из сегментных ножек, находящихся в грудном отделе зародыша (первые 8-10 сегментов), закладывается пердпочка и мезонефральный (Вольфов) проток, из которого образуются каналец придатка семенника и семявыносящий проток. Из сегментных ножек, находящихся в туловищных отделах зародыша развивается первичная почка, которая сначала функционирует у зародыша, а потом из канальцев первичной почки образуются прямые канальцы, канальцы сети семенника, выносящие канальцы придатка семенника. Сегментные ножки каудального отдела зародыша формируют нефрогенный тяж, из которого развивается окончательная почка.
Образование аллантоиса. Аллантоис образуется как вентральный вырост передней части задней кишки. Он растет кпереди, дорастает до желточного стебелька и в этой области выходит из тела зародыша. Стенка аллантоиса состоит из энтодермы и внезародышевой мезенхимы. Урахус и верхушка мочевого пузыря - это остатки аллантоиса.
Образование плаценты (III этап). По аллантоису ко вторичным ворсинкам хориона подрастают кровеносные сосуды и такие ворсинки, состоящие уже из трофобласта, внезародышевой мезенхимы и кровеносных сосудов называются третичными ворсинками хориона. Хорион прикрепляется к подлежащему участку слизистой оболочки матки - decidua basalis и вместе с ней формирует плаценту. Таким образом, плацента в буквальном смысле представляет собой соединение третичных ворсинок хориона (плодная часть) и decidua basalis (материнская часть).
19. Мезенхима
(от Мезо... и греч. énchyma — налитое, наполняющее; здесь — ткань)
зародышевая соединительная ткань большинства многоклеточных животных и человека. На ранних стадиях зародышевого развития М. состоит из подвижных отростчатых клеток, бо́льшая часть которых затем теряет подвижность; при этом клетки соединяются отростками в сеть (синцитий) или образуют местные скопления. М. возникает за счёт клеток, выселяющихся из разных зародышевых листков (См. Зародышевые листки). Главный источник её, например у кишечно-полостных, червей и моллюсков, — Эктодерма, но часто и Энтодерма, у хордовых, иглокожих и других — Мезодерма. М., возникающая из энто- и мезодермы, называется энтомезенхимой, а из эктодермы (материала нервных валиков) — эктомезенхимой. У позвоночных животных и человека энтомезенхима даёт начало различным формам соединительной ткани взрослого организма, форменным элементам крови, кровеносным сосудам и гладким мышцам. Из эктомезенхимы образуется почти весь висцеральный скелет (слуховые косточки, жаберные дуги и т. д.), пигментные клетки и часть дермы.
Иногда М. называются также малодифференцированные соединительнотканные клетки (фибробласты, ретикулярные клетки), имеющиеся в организме животных и человека в послезародышевый период их жизни. Дифференцировка мезенхимных клеток подушек в валвулосептальные фибробласты коррелирует с экспрессией двух микрофибриллярных белков, фибриллина и фибулина. Возможная роль этих белков - обеспечить поддержку клеточной адгезии, когда клетки дифференцируются. Другими матриксными белками, участвующими в ремоделировании подушек в клапанно-перегородчатую ткань, являются сульфатированные протеогликаны. Они накапливаются на клеточной поверхности или в околоклеточном матриксе мигрирующих подушечных клеток, которые их секретируют, они, по-видимому, играют важную роль в финальном позиционировании и распределении клеток внутри дифференцирующейся ткани.
20. ДИФФЕРЕНЦИРОВКА ЗАРОДЫШЕВЫХ ЛИСТКОВ.
Дифференцировка - это изменения в структуре клеток, связанные со
специализацией их функций и обусловленные активностью определенных генов.
Различают 4 этапа дифференцировки:
1. Оотипическая дифференцировка на стадии зиготы представлена
предположительными, презумптивными зачатками - участками оплодотворенной
яйцеклетки.
2. Бластомерная дифференцировка на стадии бластулы заключается в появлении
неодинаковых бластомеров (например, бластомеры крыши, дна краевых зон у
некоторых животных).
3. Зачатковая дифференцировка на стадии ранней гаструлы Возникают
обособленные участки - зародышевые листки.
4. Гистогенетическая дифференцировка на стадии поздней гаструлы. В
пределах одного листка появляются зачатки различных тканей (например, в
сомитах мезодермы). Из тканей формируются зачатки органов и систем. В
процессе гаструляции, дифференцировки зародышевых листков появляются осевой
комплекс зачатков органов.
Зародышевые листки дифференцируются у большинства позвоночных одинаково,
При этом каждый листок дифференцируется в определенном направлении. Из
первичной эктодермы образуется нервная трубка, ганглиозные пластинки,
плакоды, кожная эктодерма, прехордальная пластинка и внезародышевая
эктодерма. Первичная энтодерма является источником зародышевой кишечной
энтодермы и внезародышевой (желточной). При дифференцировке мезодермы
возникают три части: в дорсальном отделе появляются (1) сомиты, за ним
следуют (2) сегментные ножки (нефротомы), из которых образуется эпителий
почек и гонад. Вентральная мезодерма не сегментируется и формирует (3)
спланхнотом расщепляющийся на два листка: париетальный, сопровождающий
эктодерму, и висцеральный, прилежащий к энтодерме. Между листками возникает
целомическая полость, из листков спланхнотома образуется эпителий серозных
оболочек - брюшины. плевры, перикарда. Далее в теле сомита дифференцируется
из наружной его части дерматом- (источник дермы кожи), из центральной -
миотом (зачаток скелетной мышечной ткани) и из внутренней склеротом
(зачаток скелетных соединительных тканей - костей и хрящей). В процессе
дифференцировки зародышевых листков мезодермы у зародыша появляется
мезенхима.
На 20-21 сутки у эмбриона человека образуются туловищные складки,
обособляющие тело зародыша человека от внезародышевых органов и
окончательно формируются осевые зачатки органов: хорда, из эктодермы -
нервная трубка, замыкающаяся к 25 суткам. Формируется кишечная трубка.
Мезодерма зародыша дифференцируется на сомиты (сомитный период), нефротом и
спланхнотом с париетальным и висцеральными листками. В теле сомита
различают: дерматом, миотом и склеротом. В период дифференцировки мезодермы
изо всех трех зародышевых листков, но преимущественно из мезодермы,
появляются мезенхима зародыша - отростчатые клетки, эмбриональный зачаток
многих тканей и органов всех видов соединительной ткани (отсюда ее часто
называют эмбриональной соединительной тканью), а также гладкомышечной
ткани, микрооглии сосудов, крови, лимфы, кроветворных органов. Ко второму
месяцу у эмбриона человека произошел начальный гисто- и органогенез и
имеются закладки почти всех органов, К концу 8-й недели эмбриогенеза
заканчивается зародышевый период развития и начинается плодный.
Ранние стадии развития человека имеют ряд особенностей: 1. Асинхронный тип
полного неравномерного дробления с образованием “темных" и “светлых”
бластомеров; 2. Интерстициальный тип имплантации. 3. Наличие двух фаз
гаструляции - деляминации и иммиграции, между которыми бурно развиваются
внезародышевые органы; 4. Раннее обособление и формирование внезародышевых
органов; 5. Раннее образование амниотического пузырька без амниотических
складок; 6. Сильное развитие амниона, хориона и слабое-желточного метка и
аллантоиса.
Внезародышевые органы (провизорные, временные или зародышевые оболочки),
обеспечивающие развитие зародыша. В эволюции появляются впервые у рыб
(желточный мешок). У птиц имеются следующие внезародышевые органы: амнион,
сероза, желточный мешок и аллантоис. Амнион - водная оболочка, серозная -
орган дыхания. Образуются эти две оболочки у птиц путем смыкания
амниотических складок. Желточный мешок выполняет у птиц трофическую и
кроветворную функции, а аллантоис -орган выделения и газообмена у птиц.
В эмбриогенезе человека образуется пять внезародышевых органов: амнион,
желточный мешок, хорион, формирующий плаценту и аллантоис. Амнион,
создающий водную среду у человека, образуется без амниотических складок.
Желточный мешок у человека практически утрачивает трофическую и выполняет в
основном кроветворную функцию и образования первичных половых клеток.
Аллантоис. редуцирующийся на втором месяце является проводником кровеносных
сосудов к хориону. Хорошо развитый хорион у человека формирует плаценту,
за счет которой устанавливается связь зародыша и матери.
Плацента, обеспечивающая связь зародыша с организмом матери, выполняет
многочисленные функции: трофическую, дыхательную, выделительную,
эндокринную, защитную, депонирующую. По морфологическим признакам различают
четыре типа плаценты: эпителиохориальные, десмохориальные,
эндотелиохориальные и гемохориальные. Эпителиохориальные диффузные плаценты
(у дельфинов, свиней, лошадей) характеризуются врастанием ворсинок хориона
в маточные железы. В десмохориальных множественных плацентах (у корой,
овец) ворсинки хориона, разрушая эпителии маточных желез, врастают в
подлежащую соединительную ткань эндометрия матки. Эндотелиохориальный
поясной тип плаценты характерен для хищников (кошки, волки, куницы, лисы),
Хориальные ворсинки у подобного типа плацент разрушают эпителий,
соединительную ткань и контактируют с эндотелием сосудов эндометрия матки.
Гемохориальный тип плаценты (например, у летучих мышей, приматов, человека)
характеризуется разрушением стенок сосудов эндометрия матки ворсинками
хориона и непосредственным контактом их с материнской кровью. С рождением,
новорожденные, имеющие плаценты первых двух типов, способны к
самостоятельному питанию и передвижению. тогда как новорожденные с двумя
последними типами плацент после рождения долгое время не способны
самостоятельно питаться.
Мезодерма[1] (от мезо и дерма), или мезобласт — средний зародышевый листок у многоклеточных животных (кроме губок и кишечнополостных). Располагается междуэктодермой и энтодермой. У разных групп животных образуется различными способами. У плоских червей и немертин полоски мезодермы дают соединительную ткань, заполняющую пространство между внутренними органами, у кольчатых червей и большинства других беспозвоночных полоски мезодермы расчленяются на парныесомиты с вторичной полостью — целомом. У позвоночных в период нейруляции с боков от зачатка хорды мезодерма расчленяется на спинные (первичные) сегменты — сомиты, нефротомы и несегментированную брюшную мезодерму — боковые пластинки. Между двумя листками каждой из них образуется целом.
Из мезодермы впоследствии формируются хорда, хрящевой и костный скелет, мышцы, почки, кровеносные сосуды.
Мезодерма и её производные оказывают индуцирующее влияние на развитие производных эктодермы и энтодермы и в свою очередь испытывают индуцирующее влияние с их стороны.
Мезодерма дифференцируется на 3 части: 1) дорзальная часть получает название сомит и сегментируется на 44 сегмента;
2) вентральная часть — спланхнотом расщепляется на 2 листка — париетальный прилежит к эктодерме и висцеральный — прилежит к энтодерме., они замыкаются и заключают вторичную полость тела — целом;
3) участок, соединяющий сомиты и спланхнотом, — сегментная ножка, или нефрогонадотом. Нефрогонадотом сегментируется вслед за сомитами, но не до конца, в каудальном отделе ножки не разделяются и формируют диффузную нефрогенную ткань.
Каждый сомит в дальнейшем подразделяется на 3 части: склеротом — костная и хрящевая ткань осевого скелета, миотом — поперечно-полосатая скелетная мышечная ткань, и дерматом — соединительнотканная основа кожи. Нефрогонадотом даст начало эпителию выделительной и половой систем. Париетальный и висцеральный листки спланхнотома преобразуются соответственно в париетальный и висцеральный листки серозных оболочек (брюшины, плевры, перикарда), а целом — в соответствующие серозные полости тела. Помимо этого, из спланхнотома выселится большая часть клеток мезенхимы, которая даст начала соединительной и гладкомышечной ткани большинства внутренних органов. Из висцерального листка спланхнотома разовьются также корковое вещество надпочечников, миокард и эпикард сердца.