

2.2.1.4. Судьба продуктов распада аминокислот
В отличие от углеводов и нейтральных жиров, при катаболизме которых образуются лишь СО2 и Н2О, при распаде аминокислот конечным продуктом, кроме вышеназванных веществ, является аммиак. Учитывая его вероятные токсические эффекты, следует остановиться на способах обезвреживания.
Пути обезвреживания NН3
Свободный аммиак токсичен. Во-первых, он обладает хорошей растворимостью в воде, что сопровождается следующей реакцией:

Ее продукт - сильная щелочь, что грозит развитием алкалоза. Во-вторых, нарушается ЦТК, так как его метаболит – α-кетоглутарат – используется клеткой для нейтрализации гидроксида аммония. Образующаяся и накапливающаяся при этом глутаминовая кислота повреждает баланс аминокислот, в ЦНС ее гиперпродукция провоцирует декарбоксилирование, т.е. генез ГАМК, отсюда изменяется равновесие среди тормозных и возбуждающих медиаторов.
Клинически накопление аммония (гипераммониемия) проявляется рвотой, судорогами, спутанностью и помутнением сознания, вплоть до комы. Поэтому в клетках существуют следующие механизмы, предотвращающие развитие этого явления:
1) Восстановительное аминирование:
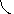
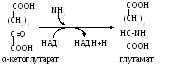




2) Во многих тканях (головном мозгу, печени, почках, мышцах) обезвреживание осуществляется путем синтеза глутамина (реже аспарагина) - нейтрального нетоксического вещества, способного диффундировать через мембраны (одна из форм транспорта NН3).
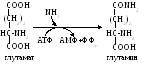
Амидная группа глутамина часто используется в реакциях аминирования (в получении глюкозамина, галактозамина, заменимых аминокислот и т.д.)
3) Работа внутри- и внеклеточных буферных систем; с помощью которых нейтрализуется NН4ОН до солей аммония.
Продукты обезвреживания аммиака (глн, асн, соли аммония) кровотоком доставляются в гепатоциты, где и происходит окончательная инактивация данного соединения:
4) Синтез мочевины.
Мочевина – полный амид угольной кислоты, значит субстратами в ее образовании служат различные формы углекислоты (НСО3– или карбоксибиотин) и аммиака (амидная группа глутамина или соль аммония): Первая реакция:
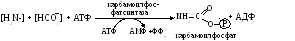
протекает в митохондриях, здесь карбамоилфосфат взаимодействует с орнитином, служащим своеобразным катализатором, вследствие чего данный процесс известен как орнитиновый цикл синтеза мочевины:
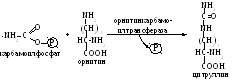
образовавшийся при этом цитруллин, выходя из митохондрий изомеризуется в енольную форму:
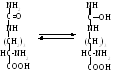
С
аргининосукцинат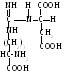
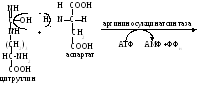 помощью специфической лигазы -
аргининосукцинатсинтазы взаимодействует
с аспарагиновой кислотой:
помощью специфической лигазы -
аргининосукцинатсинтазы взаимодействует
с аспарагиновой кислотой:


Аргининосукцинат из-за наличия сильно поляризованных связей довольно легко разрушается с помощью аргининосукцинатлиазы:
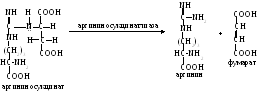
П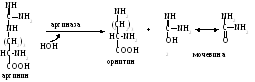 родукт
данной реакции фумарат обычно отправляется
в митохондрии, где включается в ЦТК, а
аргинин гидролизуется до мочевины и
орнитина.
родукт
данной реакции фумарат обычно отправляется
в митохондрии, где включается в ЦТК, а
аргинин гидролизуется до мочевины и
орнитина.

Последний вновь связывается с молекулой карбамоилфосфата, начиная новый цикл.
Суммарно:
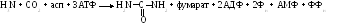
Подводя итог, следует заметить, что в целом мочевина собирает все атомы азота аминокислот: ее одна аминогруппа взята из аспартата, который может получиться путем переаминирования оксалоацетата с любой аминокислотой, другая же аминогруппа является производной аммиака, высвободившегося с помощью дезаминирования.
Двухколесный велосипед Кребса – так называют исследователи два цикла, описанные Г. Кребсом из–за их тесной связи между собой (Схема 2.2.1.4.1).
Цикл трикарбоновых кислот, тесно сопряженный с биологическим окислением и окислительным фосфорилированием, не только является поставщиком ОА для переаминирования в аспартат, но и дает СО2, НОН, АТФ для запуска и течения орнитинового цикла, одним из продуктов которого и является фумарат, служащий метаболитом ЦТК.
Нормальное течение синтеза мочевины обеспечивает:
1. окончательное обезвреживание аммиака;
2. избавление организма от NH3 и CO2;
3. баланс рН, поддерживая гомеостаз бикарбонатной буферной системы.
Для контроля нормального течения метаболизма азотсодержащих соединений, и в основном, процессов обезвреживания аммиака в клинике
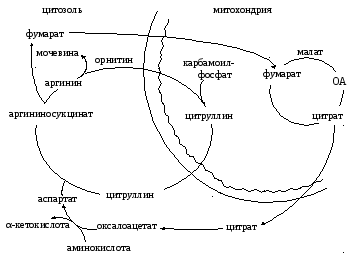
Схема 2.2.1.4.1. Взаимосвязь между ЦТК с орнитиновым циклом.
служит определение в крови следующего показателя - остаточного азота (Rest W) – т.е. оценивается суммарное содержание низкомолекулярных азотистых веществ, которые остаются фильтрате после осаждения белков. В их общем количестве около половины приходится на мочевину, 25% составляет доля аминокислот, величины креатина укладываются в 5%, ураты в 4% и т. д.
В норме значения остаточного азота колеблются в пределах 14–25 ммоль/л, у новорожденных они выше (42–71 ммоль/л). При различных патологических состояниях регистрируется гиперазотемия. В зависимости от механизмов возникновения выделяют продукционную и ретенционную гиперазотемии. Первый вариант может быть спровоцирован систематическим избыточным поступлением пищевых белков (у «мясоедов»), усиленный деструкцией тканей, сопровождающейся ускоренным распадом белков (при опухолях, лейкозах, ожоговой болезни и т.д.). Основной причиной ретенционной гиперазотемии служат обычно болезни почек при нарушении их выделительной способности, что обуславливает избыточное накопление в плазме крови мочевины. Кроме того, рост величин остаточного азота может наблюдаться при сердечно–сосудистой декомпенсации, при непроходимости кишечника (усилено всасывание продуктов гниения), сахарном диабете и т.д.
В последние годы стали проводить количественное определение отдельных компонентов остаточного азота – аминокислот (например, гипер-аминоацидемия при усиленном гидролизе тканевых белков распадающихся тканей) и мочевины (гиперкарбамидемия при болезнях почек). Гипокарбамидемия встречается довольно редко, обычно это следствие мутаций ферментов – участников орнитинового цикла (см. «Патохимия наследственных болезней» С. 57).