

Взаимосвязь обменов ак, жиров и углеводов
Метаболизм - закономерный порядок превращения веществ и энергии в живых организмах, направленных на обеспечение процесса жизнедеятельности и самовоспроизведения организма. Функции: 1)поддержание гомеостаза 2)выполнение жизненных функций Контроль, согласование и координация работы отдельных органов и ткани- три интегральных системы: -нервная система(все сигналы о изменении в окружающей среде, внутренней среде; обработка и принятие решений для адаптации организмов). -эндокринная система (открытая система- регуляция гормонами живого организма ,обеспечение гомеостаза) -сердечно-сосудистая и лимфатическая система(транспорт питательных веществ к тканях, конечных продуктов обмена к выделительным органам, транспорт всех хим. в-в, обмен между тканями)
Проявление взаимосвязи обменов: 1)одновременное поступление соединений всех классов в пищ. тракт; одновременное переваривание; одновременное всасывание; дальнейший транспорт образовавшихся мономеров в ткани 2)превращение жиров, углеводов, белков с образованием одинаковых промежуточных продуктов (ЩУК, ПВК, L-кетоглутарат, ацетил-КоА) 3)превращение по общим путям катаболизма (ПВК- окислительное декарбоксилирование, ацетил-КоА в ЦТК) 4)образование единых конечных продуктов (СО2, NН3, Н2О) 5)образование единого аккумулятора энергии-АТФ 6)все полимеры построены из небольшого числа мономеров: глюкоза, клетчатка.. 7) использование промежуточных продуктов для распада и синтеза соединений др. классов (СО2-образование ЩУК из пируватов ЦТК; глюконеогенез ацетилКоА - малонил КоА для синтеза ВЖК; синтез карбомоилфосфата-синтез пиримидиновых нуклеотидов, обезвреживание аммиака) 8)наличие соединений,являющихся пунктами переключения с одного обмена на другой 9)взаимосвязь органов и тканей печень: депонирует и поставляет глюкозу, синтез кетоновых тел, синтез холестерина, образует ЛПОНП, ЛПВП, общее обезвреживание NН3, глюкозоаланиновый и глюкозолактатный цикл почки:общее обезвреживание аммиака, кроветворение-синтез эритропоэтина, синтез ренина- АД, синтез активной формы D3-кальцитриол 10) единые механизмы регуляции на молекулярном, клеточном и организменном уровне(через изменение количества и активности ферментов)
Интеграция различных путей метаболизма углеводов четко просматриваетея на уровне метаболизма глюкозо-6-фосфата, который находится на пересечении нескольких метаболических путей. За глюкозо-6-фосфат могут конкурировать ферменты глюкозофосфапшзомераза, глюкозо - 6-фосфат -дегидрогеназа, фосфоглюкомутаза и глюкозо-6-фосфатаза. Глюкозофосфат- изомераза направляет метаболизм по гликолитическому пути. Пируват, в зависимости от обеспеченности клеток кислородом, включается в анаэробные или аэробныепроцессы. Фруктозо-6-фосфат и глицеральдегид-3-фосфат являются общими метаболитами гликолитического и пентозофосфатного пути. Они вступают в транскетолазную реакцию и дают начало не- окислительному пентозофосфатному пути. Глюкозо-6-фосфатдегидрогеназа ориентирует метаболизм глюкозо-6-фосфата через окислительный пентозофосфатный путь. Взаимосвязь между окислительно-декарбоксилируюшсй системой и системой трансферазных реакций пентозофосфатного пути (ПФП) метаболизма углеводов осуществляется ферментами рибозофосфатизомеразой и рибулозофосфат-3-эпимеразой. Взаимопревращение глюкозо-6-фосфата в глюкозо-1 -фосфат обеспечивает, с одной стороны, биосинтез гликогена, а с другой стороны, после фосфориолиза гликогена включение глюкозо-1- фосфата в гликолитический и пентозофосфатный пути метаболизма.Лактат через пируват может превращаться в оксалоацетат, который в свою очередь включается в глюкогеогенез с образованием глюкозо-6-фосфата. В печени, почках и тонком отделе кишечника происходит гидролиз глюкозо-6-фосфата с образованием глюкозы.
Взаимосвязь обмена углеводов и липидов
Биосинтез липидов из углеводов. L-Глицерофосфат, необходимый для биосинтеза ТГ (нейтральных жиров) и ФЛ (фосфатидов) образуется при восстановлении диоксиацетонфосфата (ДГАФ), который является продуктом расщепления фруктозо-1,6-бисфосфата в процессе фруктозобисфосфат-альдолазной реакции. Биосинтез высокомолекулярных карбоновых (жирных) кислот происходит из ацетил-СоА, который может образовываться в результате окислительного декарбоксилирования пирувата под действием пируватдегидрогеназного комплекса. Пируват является центральным промежуточным продуктом обмена углеводов.
Транспорт ацетил-СоА из митохондрий в цитоапазму тесно связан с обменом углеводов. Перенос ацетил-СоА из митохондрий в цитоплазму происходит в основном в виде цитрата. Цитрат, как известно, образуется в реакции конденсации ацетил-СоА с оксалоацетатом (ОАА), катализируемой цитратсинтазой. Оксалоацетат представляет собой продукт карбоксилирования пирувата.
Кроме того, окислительный пентозофосфатный путь метаболизма углеводов является одним из источников восстановительных эквивалентов при биосинтезе кислот. Так, например, для синтеза пальмитата требуется 14 молекул NAДФH, шесть из которых поставляет окислительный пентозофосфатный путь метаболизма углеводов. а восемь молекул NADPH образуется в процессе переноса 8 молекул ацетил-СоА в цитоплазму с участием пирувата и мататлегидрогеназной системы, перенос атомов водорода из разных субстратов сопровождается образованием кетокистот (предшественников глюконеогенеза) без участия атмосферного кислорода. кетокистоты в процессе карбоксилирования выделяют С02. Таким образом, образование и выделение С02 при биосинтезе жиров из углеводов происходит при меньших количествах 02,, потому что водород восстановленных коферментов идет не в дыхательную цепь, а на восстановительные биосинтетические процессы.
Биосинтез углеводов из жиров в организме животных ограничен, т. к. у них нет ферментов, способных превращать жирные кислоты в метаболиты глюконеогенеза, и только глицерол, как компонент жиров, может использоваться для биосинтеза углеводов. Растения и некоторые бактерии, благодаря наличию глиоксилатного цикла, способны использовать ацетил-СоА для биосинтеза углеводов. В результате одного оборота глиоксилатного цикла образуется две молекулы оксалоацетата, одна из которых поддерживает цикличность процесса, а вторая может быть субстратом глюконеогенеза.
Взаимосвязь обмена углеводов, нуклеиновых кислот и белков
Пентозофосфатный путь метаболизма углеводов поставляет рибозо-5-фосфат для биосинтеза 5-фосфорибозил-1-пирофосфата, который является донором рибозофосфатного остатка в биосинтезе нуклеотидов и нуклеиновых кислот.
Деградация нуклеиновых кислот и нуклеотидов сопровождается образованием рибозо-1-фосфата. Рибозо-1-фосфат изомеризуется в рибозо-5-фосфат, который, в свою очередь, может превращаться в гексозы.
Взаимосвязь между обменом нуклеиновых кислот и белков I состоит в том, что аминокислоты служат субстратами в биосинтезе нуклеиновых оснований. Так, пиримидиновый цикл синтезируется из аспартата и карбамоилфосфата, а пуриновый — из аспартата, глутамина и глицина. Кроме того, нуклеиновые кис лоты — непосредственные участники биосинтеза белков.
Биосинтез углеводов из белков. Определенная часть аминокислот у млекопитающих, после дезаминирования, в зависимости от состояния организма, может включаться в глюконеогенез с образованием глюкозы. В зависимости от способности к глюконеогенезу аминокислоты подразделяют: на глюкогенные, кетогенные, глюкогенные и кетогенные.
Глюкогенными являются четырнадцать аминокислот, из которых в процессе метаболизма образуются пируват, 2-оксоглутарат, сукцинил-КоА фумарат и оксалоацетат. Пируват и указанные метаболиты цикла трикарбоновых кислот через оксалоацетат могут превращаться в фосфоснолпируват, а затем — в глюкозу. К глюкогенным аминокислотам относятся аланин, аргинин, асппртат, аспарагин, цистсин. серин, глутамат, глутамин, глицин, гистидин, метионин, пролин. треонин, папин.
Глюкогенными и кетогенными являются пять аминокислот: изолейцин, тирозин, лизин, триптофан и фенилаланин. И3 этих аминокислот могут образовываться как предшественники глюкозы, так и аистил-КоА и 3-оксобугнрнл-КоА — предшественники кетоновых тел — 3-оксобуутирата,D-3-гидроксибутирата.У растений и многих микроорганизмов все аминокислоты могут принимать участие в синтезе глюкозы посредством реакций цикла трикарбоновых кислот и глиоксилатного цикла.
Биосинтез белков (аминокислот) из углеводов. Углеводы являются основным источником образования в организме оксокислот: пирувата, оксалоацетата и 2-оксоглутарата.В процессе восстановительного аминирования или транс- аминироваиня из пирувата будет образовывался аминокислота аланин; пируват является предшественником валина и лейцина. Оксалоацетат — предшественник аспартата. а аспартат — аспарагина, метионина’, треонина* и лизина*. Треонин в свою очередь является предшественником изолейцина*. 2-Оксоглутарат в процессе восстановительного аминирования или трансаминирования превращается в глутамат, а глутамат — в глутамин, пролин и аргинин*.
Из фосфоенолпирувата и эритрозо-4-фосфата синтезируются аминокислоты фенилаланин*, тирозин’ и триптофан*. Рибозо- 5-фосфат выполняет роль предшественника при биосинтезе гистидина’.
Аминокислота серии синтезируется из промежуточного метаболита гликолиза — 3-фосфоглицсрата. Углеводы являются не только источником оксокислот, но и атомов водорода, в виде восстановленных коферментов, в процессах биосинтеза аминокислот.
Взаимосвязь обмена белков и липидов Биосинтез жиров из белковДля биосинтеза жиров (триацилглииеронов) необходимы два компонента: жирные кислоты и глицеролфосфат Исходным метаболитом в биосинтезе жирных кислот является ацетил-СоА. Аминокислоты аланин, нистсин, глицин, серин и треонин после дезаминирования превращаются в ацетил-СоА через пируват. Безазотистые остатки лейцина, изолейцина, лизина, фенилаланина, тирозина и триптофана другими путями превращаются ацетил-СоА. Таким образом, больше половины аминокислот (И из 20) могут быть источником ацетил-СоА лля биосинтеза жирных кислот. Кроме того, существуют и другие пути образования ацетил-СоА из аминокислот.Прямых путей биосинтеза глицеролфосфата из аминокислот, по всей вероятности, не существует. Но большинство аминокислот, после дезаминирования, вовлекаются в глюконеогенез, в процессе которого легко образуются глицеральлсгид-3-фосфат и дигидроксиацетонфосфат. Последний восстанавливается до глицсролфосфата. С другой стороны, глюкоза или гликоген, синтезированные в процессе глюконеогенеза и гликогенеза, метаболизируясь гликолитическим путем, поставляют необходимые триозофосфаты для биосинтеза липидов.
Глюкозаминоглюканы
ТГ,ФЛ(соединит.ткань,
хрящи, кожа, роговица глаза)
глицерин




3-фосфоглицерин
Глюкуроновая
к-та глюкозамин
диоксиацетонфосфат ВЖК




Малонил-КоА
Глюкоза
АТФ глюкоза-6 фосфатфруктозо-1,6
бифосфат 3-ФГАПВК





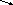


ПФП
Ацетил-КоАв-гидрокси-в-метил
НАДФН+Н
АК
глутамилКоА(ГМГ-КоА)

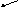


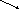
Рибозо-5-фосфат
АУК
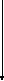
+ ЩУКФенилаланин,тирозин Холестерин желчные к-ты АСП стероидные гормоны -СО2 витаминD3

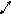
Фосфоенолпируват ЦТК
СО2, Н2О, АТФ

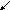

Синтез
нуклеотидов, нуклеиновых кислот
ГЕМ сукцинил КоАL-КГ
У
ГЛУ 

Б Л
(за счет глицерина 

