

МДФд_13
.pdfМДФд. Занятие № 13 |
- 11 - |
|
|
|
|
|
|
|
|

МДФд. Занятие № 13 |
- 12 - |
|
|
|
|
|
|
|
|

МДФд. Занятие № 13 |
- 13 - |
|
|
|
|
|
|
|
|

МДФд. Занятие № 13 |
- 14 - |
|
|
|
|
|
|
|
|
|
|
13В. Теоретический материал
38.Гуморальный иммунный ответ
38.1.Активация В-лимфоцитов
38.2.Отличия плазмоцита от В-лимфоцита
38.3.Особенности иммунного ответа на Т-независимые антигены
38.4.Общая схема гуморального иммунного ответа
38.5.Эффекторные функции антител
38.6.Антителозависимые механизмы защиты от патогена
39.Супрессия иммунного ответа. Динамика иммунного ответа. Первичный и вторичный иммунный ответ. Кооперативный механизм действия и регуляции иммунной системы
39.1.Супрессия иммунного ответа
39.2.Динамика иммунного ответа
39.3. Первичный и вторичный иммунный ответ
39.4.Кооперативный механизм действия и регуляции иммунной системы
40.Иммуноглобулины (антитела)
38. ГУМОРАЛЬНЫЙ ИММУННЫЙ ОТВЕТ
38.1. Активация В-лимфоцитов Активация В-лимфоцитов – ключевое событие (точнее – цепь событий) гуморального иммунного

МДФд. Занятие № 13 |
- 15 - |
ответа. Общая схема этого процесса приведена на рисунках 38.1-1 и 38.1-2.
Рис. 38.1-1. Принципиальная схема активации В-лимфоцитов
Рис. 38.1-2. Схема активации В-лимфоцитов (по Воробьеву, 2002)
А. В антигенпрзентирующей клетке (например, макрофаге) происходит процессинг антигена, который для сложных антигенов занимает 30-60 минут, а для менее сложных – 20-30 минут. В результате происходит ферментативная деградация антигена и его презентация на поверхности АПК в комплексе с антигенпредставляющей молекулой (как это описано выше в разделах 37.2 и
37.3).
Б. Вступивший в контакт с макрофагом нулевой (наивный) Т-хелпер активируется и додифференцируется в (в случае гуморального иммунного ответа)Т-хелпер второго типа(об активации Т- хелперов в ходе иммунного ответа был рассказано в разделе 37.4), который взаимодействует с соответствующим В-лимфоцитом.
В. В результате клон В-лимфоцита претерпевает ряд изменений, обусловленных главным образом информационными сигналами, предаваемыми ему Т-хелпером посредством цитокинов.
1.Сначала под воздействием интерлейкина-4 происходит активация В-лимфоцита.
2.Затем под воздействием интерлейкина-4 происходит пролиферация активированного В- лимфоцита (т.е. его размножение, так называемая экспансия клона).
3.И, наконец, на таких В-лимфоцитах появляется рецептор для взаимодействия с интерлейки- ном-6, который является сигналом дифференциации для этих клеток.
а. Под воздействием ИЛ-6 В-лимфоциты дифференцируются в плазматические клетки.
б. Однако, часть В-лимфоцитов проходит путь дифференциации не до конца. Такие не до конца дифференцированный В-лимфоциты, в отличие от плазмоцитов, являются долгоживущими клетками. При повторном контакте с причинным антигеном они заканчивают процесс дифференцировки в плазмоциты, обеспечивая более быстрый и более сильный иммунный ответ на повторный контакт с антигеном, т.е. – вторичный иммунный ответ. Благодаря этому им-

МДФд. Занятие № 13 |
- 16 - |
мунная система «запоминает» антиген, с которым ей уже приходилось «иметь дело». Такие не до конца дифференцированные В-лимфоциты называются клетками иммунологической памяти (сравните со схожими процессами при клеточном иммунном ответе, раздел 37.7.Б).
Г. Единственная задача плазматической клетки – синтез иммуноглобулинов, действие которых и составляет эффекторное звено гуморального иммунного ответа.
Д. Все виды клеток, участвующих в гуморальном иммунном ответе, находятся под контролем Т- супрессоров, способных в любой момент остановить начавшуюся иммунную реакцию (например, если она по каким-либо причинам «выйдет из-под контроля»).
38.2. Отличия плазмоцита от В-лимфоцита Несмотря на существующее мнение, что плазмоцит не является самостоятельным видом клетки, а
представляет собой терминально дифференцированный В-лимфоцит, большинство авторов считают его особой клеткой, происходящей от В-лимфоцита и имеющей, по сравнению с последней, ряд принципиальных отличий.
А. На поверхности плазматической клетки нет мембранных иммуноглобулинов (другими словами, на плазмоците не экспрессируетсяBCR).
Б. Не экспрессируются на плазматической клетке и МНС второго класса.
В. Продукцияплазмоцитомиммуноглобулиновне зависит ни от его контакта с антигеном ни от его контакта с Т-лимфоцитом. Другими словами, «запущенный» плазмоцит практически невозможно «остановить».
Г. Плазмоциты мигрируют из лимфатического узла в костный мозг или laminapropria эпителиальных тканей, где живут, продуцируя иммуноглобулины (составляющие более 20% всего белкового синтеза плазматической клетки) до 4 недель (что и соответствует продолжительности гуморального иммунного ответа).
38.3. Особенности иммунного ответа на Т-независимые антигены Развитие гуморального иммунного ответа возможно и без участия в нем Т-лимфоцитов. Антигены,
индуцирующие эту разновидность иммунного ответа, называются Т-независимыми, и такой иммунный ответ отличается рядом особенностей.
А. РаспознаваниеТ-независимогоантигена проводится самим В-лимфоцитом.
Б. При иммунном ответе на Т-независимые антигены продуцируются иммуноглобулины только класса М. Переключение на синтез IgG, как это бывает в плазматических клетках, образованных в ходе иммунного ответа на Т-зависимые антигены (эти плазмоциты тоже начинают антителопродукцию с IgM), не происходит.
В. В ходе иммунного ответа на Т-независимые антигены не образуются клетки иммунологической памяти. Это значит, что на такие антигены возможен только первичный иммунной ответ
(см. раздел 38.8).
Г. А вторичный иммунный ответ на Т-независимые антигены не возможен.
38.4. Общая схема гуморального иммунного ответа Таким образом, при гуморальном иммунном ответе события могут развиваться по двум сценари-
ям, в зависимости от того, на какой антиген – Т-зависимый или Т-независимый – формируется этот иммунный ответ (Рис. 38.4-1).
Рис. 38.4-1. Общая схема гуморального иммунного ответа
38.5. Эффекторные функции антител Основная функция антител в организме человека (но, строго говоря, не единственная, о чем уже
упоминалось, например, в разделе 31 и будет говориться в разделе 40) – осуществлять эффекторное звено гуморального иммунного ответа. Эффекторное действие антител осуществляется четырьмя основными механизмами: нейтрализацией антигена, опсонизацией, антителозависмойци-

МДФд. Занятие № 13 |
- 17 - |
тотоксичностью и активацией комплемента.
А. Если молекула иммуноглобулина реагирует с молекулой, представляющей причинный антиген, по физико-химическому или биохимическому механизму, речь идет о нейтрализации антителом соответствующего антигена.
1.При нейтрализации бактериальных токсинов, антитела, например, могут «нагружать» собой молекулу токсина и препятствовать тем самым его проникновению в клетку, не допуская, таким образом, токсин к контакту с тем внутриклеточным субстратом, действие на который и опосредует действие токсина на данный орган или ткань.
2.Действуя на ферменты вирулентности микроорганизмов, антитела, например, могут взаимодействовать на ту химическую группу в составе молекулы фермента, которая обуславливает взаимодействие этого фермента с его субстратом. В результате такое взаимодействие становится биохимически невозможным.
3.При нейтрализации вирусов антитела могут, например, «закрыть» собой молекулу на поверхности вирусной частицы, которая служит для последней молекулой адгезии. В результате вирус не может проникнуть в клетку, а, следовательно (так как он является абсолютным внутриклеточным паразитом), ни реплицироваться в ней, ни внедриться в ее геном.
Б. О явлении опсонизации, в осуществлении которого участвуют, в том числе, и антитела, рассказывалось в разделе 27.2.Б.
В. О явлении антителозависимойантитоксичности (АЗЦТ) или, что точнее, антителозависимой клеточной антитоксичности (АЗКЦТ) будет сказано ниже (раздел 38.6).
Г. О роли антител в активации комплемента на поверхности клеток, несущих чужеродный антиген, в том числе на поверхности микроорганизмов, рассказывалось в разделе 26.2.А.
38.6. Антителозависимые механизмы защиты от патогена В целом, все процессы, направленные на деструкцию и элиминацию антигена, успешное протека-
ние которых зависит от эффективного участия в них антител, называются антителозависимыми механизмами защиты от патогена. К ним относятся нейтрализация антигена связыванием его в иммунный комплекс, деструкция антигена, элиминация иммунных комплексов в совокупности с активными фракциями системы комплемента, сосудистые и гладкомышечные контрактильные реакции, абзимные свойства антител.
А. О механизме нейтрализующегодействия антител, было сказано в предыдущем (38.5.А) разделе.
Б. Деструкция иммунных комплексов, в которых антиген связывается антителами, происходит вследствие активации комплемента или усиленного поглощения их фагоцитами.
1.Активация комплемента иммунным комплексом приводит или к осмотическому лизису клетки, несущей чужеродный антиген (комплиментзависимый лизис) или к другим реакциям, которые опосредуются активными фракциями системы комплемента, присоединяющимися к иммунному комплексу (в том числе описанным ниже).
2.Иммунные комплексы а, следовательно, и заключенные в них антигены фагоцитируются с последующим уничтожением внутри фагоцитирующих клеток под влиянием факторов внутриклеточнойцитотоксичности, описанных в разделе 27.3.
В. Элиминация иммунных комплексов вследствие ассоциирования с ними активных фракций системы комплемента, осуществляется следующим образом. Эритроциты, которые обладают рецепторами к активным фракциям системы комплемента, адсорбируют на своей поверхности такие иммунные комплексы и транспортируют их в селезенку и в печень, где осуществляется их деструкция макрофагами.
Г. Антителозависимаяклеточнаяцитотоксичность (АЗКЦТ) осуществляется NK-клетками и эо-
зинофилами. Ее объектом является клетка, несущая чужеродный антиген (Рис. 38.6-1).
Рис. 38.6-1. Общая схема АЗКЦТ
МДФд. Занятие № 13 |
- 18 - |
1.NK-клетки несут на своей внешней мембране рецептор для Fc-фрагментов IgG. Благодаря этому они распознают клетки, на поверхность которых «сели» иммуноглобулины класса G (чаще всего – это клетки, инфицированные вирусами). ВнеклеточнаяцитотоксичностьNKклеток осуществляется посредством действия перфоринов и запуска в клетке-мишени процесса апоптоза.
2.Если патоген слишком велик для фагоцитирования (например, гельминт), то АЗКЦТ осуществляются с помощью эозинофилов, которые имеют рецепторы для адсорбции IgA илиIgE, благодаря чему «нагружают» собой патоген и, синтезируя высокотоксичные протеины, запускают так называемый «эозинофильный воспалительный процесс».
Д. Иммунные комплексы, формирующиеся в ходе гуморального иммунного ответа на поверхности тучных клеток и базофилов, активируют синтез этими клетками биологически активных ве-
ществ, которые вызывают сосудистые и гладкомышечные контрактильные реакции (срав-
ните с содержанием раздела 41.5).
Е. Вносят свой вклад в защиту от патогенна и абзимные свойства антител (см. раздел 40), их протеазная и нуклеазная активность
39. СУПРЕССИЯ ИММУННОГО ОТВЕТА. ДИНАМИКА ИММУННОГО ОТВЕТА. ПЕРВИЧНЫЙ И ВТОРИЧНЫЙ ИММУННЫЙ ОТВЕТ. КООПЕРАТИВНЫЙ МЕХАНИЗМ ДЕЙСТВИЯ И РЕГУЛЯЦИИ ИММУННОЙ СИСТЕМЫ
39.1. Супрессия иммунного ответа При любом виде иммунного ответа, в случае достижения эффекта (деструкции и элиминации при-
чинного антигена) этот иммунный ответ завершается своей супрессией (т.е. подавлением), которая обуславливается целым рядом факторов (см. раздел 37.7.А).
А. Элиминация (уничтожение) причинного антигена означает устранение фактора активации лимфоцитов через их антигенраспознающие рецепторы (TCRи BCR).
Б. Продолжительность жизнитерминальнодифференцированных лимфоцитов ограничена, это короткоживущие клетки, они погибают по механизму апоптоза. В результате, без новых «порций» активированных клеток, общее количество активированных лимфоцитов данного клона неудержимо стремится к нулю.
В. Кроме этого, к супрессии иммунного ответа приводит и торможение активности лимфоцитов по принципу обратной связи (в результате взаимодействия цитокинов и IgG с ингибирующими рецепторами, которые экспрессируются на терминально дифференцированных лимфоцитах).
Г. На стадии супрессии иммунного ответа активируются особые Т-киллеры, которые запускают процесс апоптоза в активированных Т-лимфоцитах. Эти Т-киллеры носят название Т- супрессоров, их особенно много в печени.
Д. Активированный лейкоциты также погибают в результате апоптоза.
Е. Кроме этого, формируется супрессия активности «нанятых» для осуществления иммунного ответа лейкоцитов цитокинами, действующими на негативные корецепторы, которые сопутствуют на этих клетках рецепторам для факторов роста и для гормонов).
39.2. Динамика иммунного ответа Деление иммунного ответа на клеточный и гуморальный имеет, в основном, дидактическое значе-
ние и свидетельствует лишь о том, какие эффекторные механизмы – клеточные или гуморальные – преобладают в каждом конкретном случае. В целом же иммунный ответ проходит следующие основные этапы своего развития (завершаясь, как было сказано в предыдущем разделе, своейсупрессией, механизмы которой начинают стимулироваться одновременно с развитием иммунного ответа).

МДФд. Занятие № 13 |
- 19 - |
Рис. 39.2-1. Интегральная схема иммунного ответа и связь между неспецифической резистентностью и антигенспецифическим иммунным ответом
А. Процессинг и представление антигена.Процессинг (деградация) антигенапроисходит внутри клетки и приводят к фрагментации (протеолизу) белков, их связыванию с молекулами MHC и экспрессии в виде короткого пептида на мембране клетки в комплексе с молекулами МНС (представление). В таком виде антиген может распознаваться Т-клеточным рецептором Т- хелпера. Молекулы MHCI класса представляют продукт деградации эндогенных белков, которые находятся в цитозоле АПК, а MHCII класса представляют пептидные фрагменты – продукты образовавшиеся при разрушении экзогенных белков, локализованных во внутриклеточных компартментах. Все ядросодержащие клетки имеют на мембране МНСI, а комплекс МНСII обнаружен только на мембране антигенпредставляющих клеток (макрофагов, дендритных клеток и В- лимфоцитов).
Б. Рестрикция и индукция иммунного ответа. В зависимости от того, в составе какого комплекса представлен антиген, происходит стимуляция соответствующего клона Т-клеток: антиген в составе МНСI активирует цитотоксические лимфоциты. Т-хелперы распознают антиген в составе МНСII. Если антиген представляют В-лимфоциты происходит активация Т-хелперов 2 порядка.
В. Эффекторная стадия. На этой стадии происходит реализация цитотоксичности ЦТЛ и превращение В-лимфоцитов в плазматические клетки, а также синтез ими антител.
Г. Одновременно с развитием иммунного ответа начинают стимулироваться механизмы, тормозящие его, включая активацию клеток-супрессоров.
39.3. Первичный и вторичный иммунный ответ В зависимости от характера контакта с антигеном (а точнее – наличием в организме клеток имму-
нологической памяти, несущих рецептор к причинному антигену) различают первичный и вторичный иммунный ответ (Рис. 39.3-1).
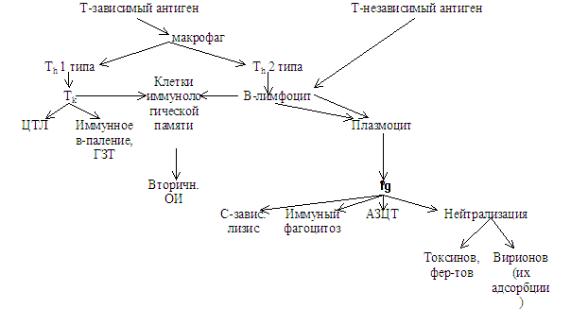
МДФд. Занятие № 13 |
- 20 - |
Рис. Рис. 39.3-1. Общая схема гуморального и клеточного иммунного ответа на Т-зависимые и Т- независимые антигены
А. Первичный иммунный ответ развивается после первого контекта с антигеном. Для него характерны следующие особенности.
–Наличие латентного периода (2-3 дня после первого контакта с антигеном). Это связано с отсутствием лимфоцитов памяти. Все клоны лимфоцитов находятся в фазе покоя G0. При поступлении в организм антигена вначале синтезируются IgM (антитела выявляются через 2-3 суток), а затем – IgG (пик приходится на 10-14 сутки, причем эти антитела могут сохранятся в низком титре в течение всей жизни). Отмечается также небольшое увеличение уровней IgA, IgE и IgD. Образуются комплексы антиген-антитело.
–Уже с третьих суток появляются иммунные Т-лимфоциты.
–Первичный иммунный ответ затихает через 2-3 недели после стимуляции антигеном.
–Появляются лимфоциты памяти и может долго поддерживаться следовой уровень IgG.
Б. Вторичный иммунный ответ развиваетсяпосле повторного контакта с тем же антигеном и имеет следующие особенности.
–В организме уже имеются долгоживущие клоны антигенспецифическихТ- и В-лимфоцитов памяти, ответственных за «память» об антигене и способных к рециркуляции, они находятся не в покое, а в фазе G1.
–Стимуляция синтеза антител и иммунных Т-лимфоцитов наступает через 1-3 дня.
–Т-клетки памяти быстро превращаются в эффекторные.
–Количество антител сразу резко увеличивается, причем синтезируются иммуноглобулины высокой специфичности – IgG.
–Чем больше контактов с антигенами имело место в данном организме, тем выше будет концентрация и специфичность (аффинность) антител.
39.4. Кооперативный механизм действия и регуляции иммунной системы Кооперативные механизм действия и регуляции иммунной системы осуществляется на двух уровнях.
А. Внутри иммунной системы кооперация и регуляция осуществляется посредством межклеточных взаимоотношений иммуноцитов и «нанятых» иммунной системой клеток, в том числе с помощью цитокинов.
Б. Межсистемная кооперация и регуляция иммунной системы осуществляется на уровне организма с вовлечением центральной нервной и эндокринной систем (Рис. 39.4-1), алгоритм кото-
рой впервые предложил П.Ф.Здродовский (гипоталамоадреналовая теория иммунитета Здро-
довского). О вовлечении системы иммунитета в месистемную кооперацию на организменном уровне свидетельствуют следующие факты.