

ГОСУДАРСТВЕННОЕ БЮДЖЕТНОЕ ОБРАЗОВАТЕЛЬНОЕ УЧРЕЖДЕНИЕ
ВЫСШЕГО ПРОФЕССИОНАЛЬНОГО ОБРАЗОВАНИЯ
Новосибирский государственный медицинский университет
МИНИСТЕРСТВА ЗДРАВООХРАНЕНИЯ
РОССИЙСКОЙ ФЕДЕРАЦИИ
(ГБОУ ВПО НГМУ Минздрава России)
Фармацевтический факультет
Кафедра фармацевтической химии
Контрольная работа №1
По биохимии клетки
Вариант №36(2,4),39(2)
Выполнил:
студент 5 курса _ группы
фармацевтического факультета
заочного отделения
Коцаренко Илья Михайлович.
89139521604
Проверил:
Новосибирск-2016г.
Содержание
Регуляция метаболических процессов с помощью белок-белкового взаимодействия, путем фосфорилировани-дефосфорилирования и частичным (ограниченным) протеолизом.
Цикл лимонной кислоты. Ход процесса. Значение. Регуляция.
Ингибирование ферментативной активности. Обратимое ингибирование: конкурентное и неконкурентное. Лекарственные вещества – конкурентые ингибиторы ферментативной активности.
Список литературы
1.Регуляция метаболических процессов с помощью белок-белкового взаимодействия, путем фосфорилировани-дефосфорилирования и частичным (ограниченным) протеолизом.
Некоторые ферменты, функционирующие вне клеток (в ЖКТ или в плазме крови), синтезируются в виде неактивных предшественников и активируются только в результате гидролиза одной или нескольких определённых пептидных связей, что приводит к отщеплению части белковой молекулы предшественника. В результате в оставшейся части белковой молекулы происходит конформационная перестройка и формируется активный центр фермента.
Рассмотрим механизм частичного протеолиза на примере активации протеолитического фермента трипсина (рис. 1). Трипсиноген, синтезируемый в поджелудочной железе, при пищеварении по протокам поджелудочной железы поступает в двенадцатиперстную кишку, где и активируется путём частичного протеолиза под действием фермента кишечника энтеропептидазы. В результате отщепления гексапептида с N-конца формируется активный центр в оставшейся части молекулы. Следует напомнить,

Рис. 1. Регуляция активности ферментов осфорилированием/дефосфорилированием.
что трипсин относят к семейству "сериновых" протеаз - активный центр фермента содержит функционально важный остаток Сер.
Частичный протеолиз - пример регуляции, когда активность фермента изменяется необратимо. Такие ферменты функционируют, как правило, в течение короткого времени, определяемого временем жизни белковой молекулы. Частичный протеолиз лежит в основе активации протеолитических ферментов, белков свёртывающей системы крови и фибринолиза, белков системы комплемента, а также пептидных гормонов.[1]
2.Цикл лимонной кислоты. Ход процесса. Значение. Регуляция.
Трикарбоновых кислот цикл - цикл лимонной кислоты или цикл Кребса – широко представленный в организмах животных, растений и микробов путь окислительных превращений ди- и трикарбоновых кислот, образующихся в качестве промежуточных продуктов при распаде и синтезе белков, жиров и углеводов. Открыт Х.Кребсом и У.Джонсоном (1937). Этот цикл является основой метаболизма и выполняет две важных функции – снабжения организма энергией и интеграции всех главных метаболических потоков, как катаболических (биорасщепление), так и анаболических (биосинтез).
Цикл Кребса состоит из 8 стадий (в двух стадиях на схеме выделены промежуточные продукты), в ходе которых происходит:
1) полное окисление ацетильного остатка до двух молекул СО2,
2) образуются три молекулы восстановленного никотинамидадениндинуклеотида (НАДН) и одна восстановленного флавинадениндинуклеотида (ФАДН2), что является главным источником энергии, производимой в цикле и
3) образуется одна молекула гуанозинтрифосфата (ГТФ) в результате так называемого субстратного окисления.
В целом, путь энргетически выгоден (DG0' = –14,8 ккал.)
Цикл Кребса, локализованный в митохондриях, начинается с лимонной кислоты (цитрат) и заканчивается образованием щавелевоуксусной кислоты (оксалоацетата – ОА). К субстратам цикла относятся трикарбоновые кислоты – лимонная, цис-аконитовая, изолимонная, щавелевоянтарная (оксалосукцинат) и дикарбоновые кислоты – 2-кетоглутаровая (КГ), янтарная, фумаровая, яблочная (малат) и щавелевоуксусная. К субстратам цикла Кребса следует отнести и уксусную кислоту, которая в активной форме (т.е. в виде ацетилкофермента А, ацетил-SКоА) участвует в конденсации с щавелевоуксусной кислотой, приводящей к образованию лимонной кислоты. Окисляется именно ацетильный остаток, вошедший в структуру лимонной кислоты, подвергается окислению; атомы углерода окисляются до CO2, атомы водорода частично акцептируются коферментами дегидрогеназ, частично в протонированной форме переходят в раствор, то есть в окружающую среду.
Как исходное соединение для образования ацетил-КоА обычно указывается пировиноградная кислота (пируват), образующаяся при гликолизе и занимающая одно из центральных мест в перекрещивающихся путях обмена веществ. Под влиянием фермента сложной структуры – пируватдегидрогеназы (КФ1.2.4.1 – ПДГаза) пирувата окисляется с образованием CO2(первое декарбоксилирование), ацетил-КоА и восстановливается НАД (см. схему). Однако окисление пирувата – далеко не единственный путь образования ацетил-КоА, который также является характерным продуктом окисления жирных кислот (фермент тиолаза или синтетаза жирных кислот) и других реакций разложения углеводов и аминокислот. Все ферменты, участвующие в реакциях цикла Кребса, локализованы в митохондриях, причем большинство из них растворимы, а сукцинатдегидрогеназа (КФ1.3.99.1) прочно связана с мембранными структурами.
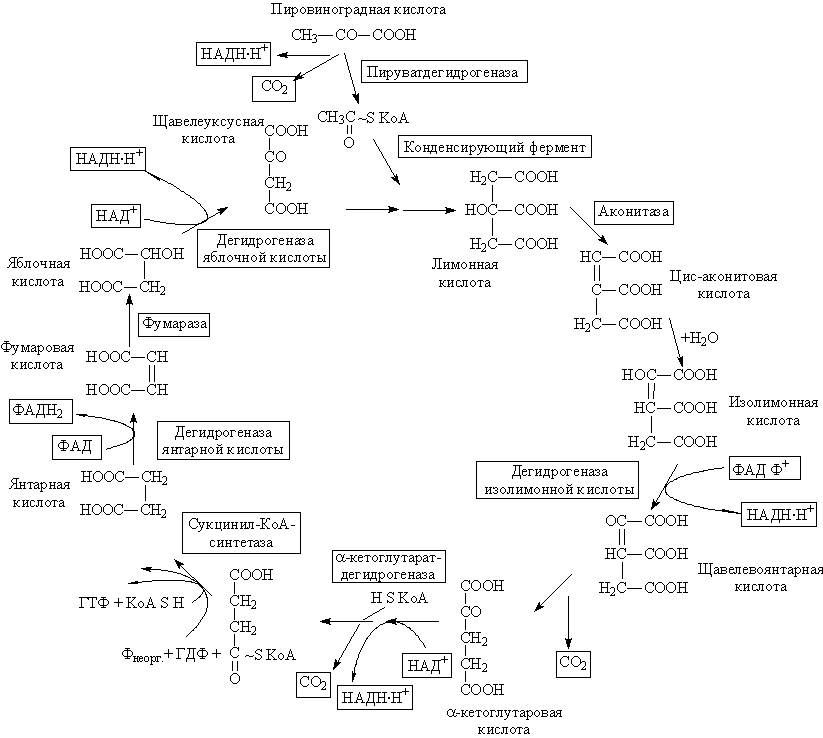
Образование лимонной кислоты, с синтеза которой и начинается собственно цикл, при помощи цитратсинтазы (КФ4.1.3.7 – конденсирующий фермент на схеме), является реакцией эндергонической (с поглощением энергии), и ее реализация возможна благодаря использованию богатой энергией связи ацетильного остатка с KoA [СН3СО~SKoA]. Это главная стадия регуляции всего цикла. Далее следует изомеризация лимонной кислоты в изолимонную через промежуточную стадию образования цис-аконитовой кислоты (фермент аконитаза КФ4.2.1.3, обладает абсолютной стереоспецифичностью – чувствительностью к местоположению водорода). Продуктом дальнейшего превращения изолимонной кислоты под влиянием соответствующей дегидрогеназы (изоцитратдегидрогеназа КФ1.1.1.41) является, по-видимому, щавелевоянтарная кислота, декарбоксилирование которой (вторая молекула CO2) приводит к КГ. Эта стадия также строго регулируется. По ряду характеристик (высокая молекулярная масса, сложная многокомпонентная структура, ступенчатые реакции, частично те же коферменты и т.д.) КГдегидрогеназа (КФ1.2.4.2) напоминает ПДГазу. Продуктами реакции являются CO2(третье декарбоксилирование), Н+и сукцинил-КоА. На этой стадии включается сукцинил-КоА-синтетаза, иначе называемая сукцинаттиокиназой (КФ6.2.1.4), катализирующая обратимую реакцию образования свободного сукцината: Сукцинил-КоА + Рнеорг+ ГДФ = Сукцинат + KoA + ГТФ. При этой реакции осуществляется так называемое субстратное фосфорилирование, т.е. образование богатого энергией гуанозинтрифосфата (ГТФ) за счет гуанозиндифосфата (ГДФ) и минерального фосфата (Рнеорг) с использованием энергии сукцинил-КоА. После образования сукцината вступает в действие сукцинатдегидрогеназа (КФ1.3.99.1) – флавопротеид, приводящий к фумаровой кислоте. ФАД соединен с белковой частью фермента и является метаболически активной формой рибофлавина (витамин В2). Этот фермент также характеризуется абсолютной стереоспецифичностью элиминирования водорода. Фумараза (КФ4.2.1.2) обеспечивает равновесие между фумаровой кислотой и яблочной (также стереоспецифична), а дегидрогеназа яблочной кислоты (малатдегидрогеназа КФ1.1.1.37, нуждающаяся в коферменте НАД+, также стереоспецифична ) приводит к завершению цикла Кребса, то есть к образованию щавелевоуксусной кислоты. После этого повторяется реакция конденсации щавелевоуксусной кислотой с ацетил-КоА, приводящая к образованию лимонной кислоты, и цикл возобновляется.
Сукцинатдегидрогеназа входит в состав более сложного сукцинатдегидрогеназного комплекса (комплекса II) дыхательной цепи, поставляя восстановительные эквиваленты, (НАД-Н2), образующиеся прив реакции, в дыхательную цепь.
На примере ПДГазы можно познакомиться с принципом каскадной регуляции активности метаболизма за счет фосфорилирования-дефосфорилирования соответствующего фермента специальными киназой и фосфатазой ПДГазы. Обе они присоединены к ПДГазе.
Предполагается, что катализ индивидуальных ферментативных реакций осуществляется в составе надмолекулярного «сверхкомплекса», так называемого «метаболона». Преимущества такой организации ферментов состоят в том, что нет диффузии кофакторов (коферментов и ионов металлов) и субстратов, а это способствует более эффективной работе цикла.
Энергетическая эффективность рассмотренных процессов невелика, однако образующиеся при окислении пирувата и последующих реакциях цикла Кребса 3 моля НАДН и 1 моль ФАДН2являются важными продуктами окислительных превращений. Дальнейшее их окисление осуществляется ферментами дыхательной цепи также в митохондриях и сопряжено с фосфорилированием, т.е. образованием АТФ за счет этерификации (образования фосфороорганических эфиров)минерального фосфата. Гликолиз, ферментное действие ПДГазы и цикл Кребса – всего в сумме 19 реакций – определяют полное окисление одной молекулы глюкозы до 6 молекул CO2с образованием 38 молекул АТФ – этой разменной «энергетической валюты» клетки. Процесс окисления НАДН и ФАДН2ферментами дыхательной цепи энергетически весьма эффективен, происходит с использованием кислорода воздуха, приводит к образованию воды и служит основным источником энергетических ресурсов клетки (более 90%). Однако в его непосредственной реализации ферменты цикла Кребса не участвуют. В каждой клетке человека есть от 100 до 1000 митохондрий, обеспечивающих жизнедеятельность энергией.
В основе интегрирующей функции цикла Кребса в метаболизме лежит то, что углеводы, жиры и аминокислоты из белков могут превращаться в конечном счете в интермедиаты (промежуточные соединения) этого цикла или синтезироваться из них. Выведение интермедиатов из цикла при анаболизме должно сочетаться с продолжением катаболической активности цикла для постоянного образования АТФ, необходимого для биосинтезов. Таким образом, цикл должен одновременно выполнять две функции. При этом концентрация интермедиатов (особенно ОА) может понижаться, что способно привести к опасному понижению производства энергии. Для предотвращения служат «предохранительные клапаны», называемые анаплеротическими реакциями (от греч. «наполнять»). Важнейшей является реакция синтеза ОА из пирувата, осуществляемая пируваткарбоксилазой, также локализованной в митохондриях. В результате накапливается большое количество ОА, что обеспечивает синтез цитрата и др. интермедиатов, что позволяет циклу Кребса нормально функционировать и, вместе с тем, обеспечивать выведение интермедиатов в цитоплазму для последующих биосинтезов. Таким образом, на уровне цикла Кребса происходит эффективно скоординированная интеграция процессов анаболизма и катаболизма под действием многочисленных и тонких регуляторных механизмов, в том числе гормональных.
В анаэробных условиях вместо цикла Кребса функционируют его окислительная ветвь до КГ (реакции 1, 2, 3) и восстановительная – от ОА до сукцината (реакции 8®7®6). При этом много энергии не запасается и цикл поставляет только интермедиаты для клеточных синтезов.
При переходе организма от покоя к активности возникает потребность в мобилизации энергии и обменных процессов. Это, в частности, достигается у животных шунтированием наиболее медленных реакций (1–3) и преимущественным окислением сукцината. При этом КГ – исходный субстрат укороченного цикла Кребса – образуется в реакции быстрого переаминирования (переноса аминной группы)
Глутамат + ОА = КГ + аспартат
Другая модификация цикла Кребса (так называемый 4-аминобутиратный шунт) – это превращение КГ в сукцинат через глутамат, 4-аминобутират и янтарный семиальдегид (3-формилпропионовую кислоту). Эта модификация важна в ткани мозга, где около 10% глюкозы расщепляется по этому пути.
Тесное сопряжение цикла Кребса с дыхательной цепью, особенно в митохондриях животных, а также ингибирование большинства ферментов цикла под действием АТФ, предопределяют снижение активности цикла при высоком фосфорильном потенциале клетки, т.е. при высоком соотношении концентраций АТФ/АДФ. У большинства растений, бактерий и многих грибов тесное сопряжение преодолевается развитием несопряженных альтернативных путей окисления, позволяющих поддерживать одновременно дыхательную активность и активность цикла на высоком уровне даже при высоком фосфорильном потенциале.[2,3,4,5]