

Лекция 1. ДНК. ИСТОРИЯ ОТКРЫТИЯ. СОВРЕМЕННЫЕ ПРЕДСТАВЛЕНИЯ.
1869 г. - Фридерих Мишер впервые, изучая лейкоциты гноя человека, выделяет из ядер лейкоцитов вещество, которое резко отличалось от известных тогда белков, углеводов, жиров. Поскольку ядро клетки в 1831 г. англ. ученым Робертом Броуном было названо нуклеус, то выделенное вещество Мишер назвал нуклеином.
Братья Оскар и Рихард Гертвиги впервые предположили, что нуклеид - вещество наследственности.
В1891 г. профессор берлинского университета Альбрехт Коссель впервые при изучении нуклеина обнаружил в его составе пуриновые азотистые основания (аденин и гуанин). Он же в 1901 г. обнаружил в нуклеине пиримидиновые азотистые основания (тимин
ицитозин).
В1909 г. французский ученый Питер Левин установил, что в нуклеине есть остаток фосфорной кислоты и сахар рибоза. В 1930 г. он же находит дезоксирибозу.
В20-х гг. прошлого века ученые поняли, что нуклеид (позднее - нуклеиновые кислоты) разделяются на 2 вещества: перв. назвали тимонуклеиновой кислотой, т. к. ее обнаружили в тимусе теленка и стали считать, что эта кислота характерна для животных. Вторая - дрожжеая кислота (найдена в клетках дрожжей), и стали считать, что она характерна для растений.
В30-е гг. были найдены красители, которые в клетке по-разному окрашивали эти 2 кислоты. И вот, почти одновременно, сразу несколько ученых (французский ученый Касперсон, отечественный ученый Кедровский и американцы Мирский, Шпигельман, Камен) пришли к выводу, что первая тимонуклеиновая кислота есть и у растений, и у животных только в ядре клетки, и ее стали называть ДНК, а вторая кислота (дрожжевая) - у всех, но она содержится, в основном, в цитоплазме и ядрышке ядра клетки.
1950 г. - американский ученый Эривин Чаргафф впервые при изучении ДНК приходит к следующему заключению:
1) количество пуриновых основания = количеству пиримидиновых азотистых оснований; 2) содержание в клетке А = Т 3) содержание в клетке Ц = Г
Морису Уилкинсу и Розалинде Франклин впервые удалось сделать поперечный срез ДНК и сделать фото с помощью рентгеновского микроскопа. При этом они получили интересную картину.
Анализируя это изображение, они приходят к следующим соображениям:
1)ДНК состоит из 2 цепей
2)цепь спирально закручена, ее диаметр = 2 нм
3)цепь состоит из повторяющихся элементов, каждый из которых занимает 0,34 нм
4)на виток спирали ДНК приходится около 10 повторов, и сам виток равен 3,4 нм
В1953 г двое молодых ученых - американцы Джеймс Уотсон (биохимик) и Френсис Крик (биофизик), воспользовавшись данными, которые получили Уилкинс и Франклин, попытались создать представление о том, как устроена ДНК.
Они пришли к выводу, что нити ДНК антипараллельны друг другу
Предположим, что репликация:  Нить растет, на ее основе синтезируются новые цепи.
Нить растет, на ее основе синтезируются новые цепи.
Артур Корнберг открыл фермент ДНК-полимеразу-I, который способен синтезировать ДНК, но ему нужны одноцепочечные нити ДНК, которые идут в направлении 3'-5'.
1
Выходило, что реплицировать фермент может только на 3'-5', но не на др. (5'-3') Т. е.
1)реплицируется только 1 нить (опровергнуто)
2)существует другой фермент, который может реплицировать ДНК на 5'-3' Было открыто множество ферментов.
Филипп Хеневалт изучал процесс репликации у кишечной палочки. Установил, что
полная репликация ДНК происходит за 15 минут. Зная количество нуклеотидов, вычислив, сколько витков спирали ДНК имеет кишечная палочка, пришел к выводу: чтобы реплицироваться за счет раскручивания ДНК за 15 минут, надо крутиться со скоростью 60 тыс. оборотов/мин (что невозможно).
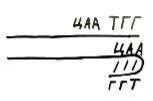
Впервые ученый Оказаки при изучении репликации обнаружил, что та идет фрагментами, примерно 1100 нуклеотидов. Их стали называть фрагментами Оказаки.
Ричардсон и Вейс обнаружили фермент, с помощью которого эти фрагменты могут сшиваться вместе - лигаза.
Еще в 1954 году американский ученый Дельбрюк и Гюнтер Стент после открытия Уотсона и Крика предположили, что репликация может идти:
1)консервативным способом, согласно которому двухцепочечная нить ДНК производит новую дочернюю нить ДНК, которая не содержит никаких старых элементов ДНК
2)полуконсервативным способом: старая нить дает 2 гибридных, в которых одна нить старая, а вторая новая
3)дисперсным способом: старая нить разрезается на кусочки, каждый из которых реплицируется отдельно, а затем эти куски сшиваются
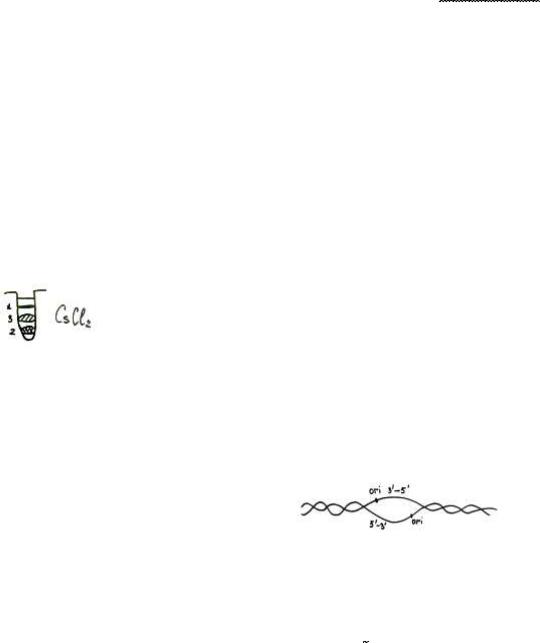
Мезельсон и Сталь решили проверить эти методы. Они работали на кишечной палочке, выращивая ее в питательной среде, содержащей тяжелый N15, и через одну генерацию они поместили выделенную ДНК в специальный раствор, содержащий CsCl2 . Этот раствор обладает градиентом плотности (чем ниже к донцу, тем плотнее).
Варианты:
1)обе нити имеют N14 и N14
2)-||- N15 и N15
3)-||- N14 и N15
Дисперсность не рассматривали. Все сосредоточилось в середине (3). Вывод: репликация идет полуконсервативным методом.
СОВРЕМЕННЫЕ ВОЗЗРЕНИЯ НА РЕПЛИКАЦИЮ ДНК.
Идея о раскручивании ДНК по все длине равномерно была отвергнута. Репликация идет участками (примерно 110 нуклеотидов). Эти участки называются репликонами. Одновременно репликация ДНК может идти во многих точках ДНК.
Точка репликации называется точкой ori.
Сперва нужно реплицировать маленький фрагмент, а для этого нужно раскрутить участок ДНК. Этим процессом занимается фермент ДНК-гираза (топоизомераза-II). Она начинает раскручивание в точке ori, при этом спиральность сдвигается на концы репликона, и здесь наблюдается суперскручивание ДНК и переплетение ДНК.
ДНК-хеликаза способна разрезать спутанные части ДНК. ДНК-свебилаза в области фосфатных мостиков делает надрез (шарнир Кэрнса), через него суперспираль сбрасывается. Чтобы нити вновь не склеились, появляются SSB-белки на поверхности шарнира Кэрнса.
Праймосома (мобильный промотор репликации) состоит из 3 белковых ферментов:
1)праймаза
2)ДНК-зависимая рибонуклеозид-3-фосфотаза (ДНАБ)
3)ДНК-зависимая атефаза (N'-белок)
2

Праймосома присоединяется к точке ori. Начинается синтез праймера (затравки), примерно 6 нуклеотидов РНК. К этой затравке присоединяется ДНК-полимераза, которая движется, синтезируя по 5'-3' и только на цепи 3'-5'. Эта нить является лидируещей, она непрерывна. Противоположная нить синтезируется участками [праймер -> затравка -> ДНК- полимераза-3 -> репликация до точки ori], эта нить называется запаздывающей.
Далее синтезируется участок соединяющийся с помощью ДНК-лигазы. В дальнейшем, когда репликация подойдет к концу, вновь начнет действовать топоизомераза- III, происходит восстановление спиральности.
Таким образом:
1)в начале репликации ДНК-гираза (топоизомераза-II) раскручивает участок ДНК - репликон в точке ori
2)на границах репликона возникает суперспираль
3)ДНК-свебилаза надрезает фосфатные мостики, образуя шарнир Кэрнса, и происходит сбрасывание излишек спирали
4)хеликазы надрезают нити ДНК в местах скручивания
5)SSB-белки стабилизируют одноцепочечную ДНК
6)праймосома за счет фермента праймазы, белков ДНАБ и N' образует РНК-затравку в точке ori
7)ДНК-полимераза-III присоединяется к затравке и стимулирует рост молекулы ДНК - элонгация
8)РНК-затравка удаляется с помощью РНКазы, и образовавшаяся брешь застраивается ДНК- полимеразой-I. Противоположная цепь ДНК синтезируется участками Оказаки. Все образовавшиеся ферменты сшиваются вместе ДНК-лигазой. Топоизомераза-III в дальнейшем восстанавливает спиральность ДНК.
ДОКАЗАТЕЛЬСТВА ГЕНЕТИЧЕСКОЙ РОЛИ ДНК. (ДНК - носитель генетической информации)
1) Впервые в 1928 г. немецкий ученый Гриффитс экспериментировал на бактерии пневмококк, обладающей полисахаридной капсулой. Убивает нагреванием опасный пневмококк. Убитых бактерий мешает с авирулентными. Этой смесью заражает мышей. Мышь гибнет. Из ее легких выделяет живые капсульные формы. Вывод: живые авирулентные микробы способны захватить генетическую информацию у убитой формы и превратиться в вирулентный штамм. Явление было названо трансформацией (способность к захвату генетической информации у другого организма с изменением генетических свойств).
1944 г. - американцы Эвери, МакКлеод, МакКарти решили повторить вышеупомянутый опыт с выделением ДНК. Они выделили ДНК у вирулентной особи и стали культивировать авирулентов с этой ДНК. В результате, обнаружили, что они превращаются в капсульные формы.
2) Американские ученые Хериш и Чейз провели эксперимент с бактериофагом. Они культивировали бактериофаг и вводили S35 и P32. Заразили бактерий бактериофагами. Оказалось, что внутрь проник только Р32, при этом размножение бактериофагов с биосинтезом капсида.
3
Лекция 2. ТРАНСКРИПЦИЯ Почему РНК стала посредником между ДНК и белком? По-видимому, в первичной древней
клетке различий между отдельными видами нуклеиновых кислот не существовало. Все типы РНК (иРНК (мРНК), рРНК, тРНК) синтезировались в ядре клетки на молекуле ДНК.
Согласно Фр. Крику центральная догма молекулярной биологии гласит: перенос генетической информации происходит в 3 этапа:
1)с ДНК на ДНК (репликация)
2)с ДНК на РНК (транскрипция)
3)с РНК на белок (трансляция)
Позднее было открыто явление обратной транскрипции, когда под влиянием обратных транскриптаз/ревертаз на основе молекулы РНК синтезируется ДНК.
У эукариотических организмов, имеющих настоящее ядро, транскрипция и трансляция разделены во времени и пространстве. Транскрипция - в ядре, трансляция - в цитоплазме. У прокариот эти два процесса происходят одновременно. Началась транскрипция, образуется иРНК, к ней тут же присоединяется рибосома, и тут же синтезируется белок. У прокариот иРНК с гена синтезируется за 2,5 мин, а уже через 30 с появляется белок. Но у прокариот иРНК существует только на 1 синтез белка, и тут же распадается. У эукариот иРНК может существовать очень долго.
Из 2 нитей ДНК только 1 имеет смысл, с нее синтезируется иРНК, эту нить называют антикодирующей, а вторая нить (кодирующая) по составу - копия иРНК. Синтез, как и у ДНК, идет в направлении 5'-3' и начинается с 3'-конца ДНК. При этом у двух рядом расположенных генов смысл антикодирующей цепи может быть разным, т.е. с первого гена иРНК образуется с одной цепи ДНК, а рядом со второго - с другой.
Существование иРНК теоретически было предсказано Уотсоном и Криком. РНК - линейные полинуклеотиды похожие на ДНК:
1)из 4 видов нуклеотидов, которые включают азотисты основания, фосфатный остаток и пентозный сахар
2)нуклеотиды связаны в цепь с помощью 5'-3'-фосфодиэфирными связями.
Отличия РНК от ДНК:
1)пентоза ДНК - дезоксирибоза, а в РНК - рибоза, которая имеет дополнительную гидроксид группу, и это является препятствием для двуцепочечности РНК
2)среди 4 нуклеотидов вместо Т стоит У, отличающийся тем, что у него отсутствует метильная группа в 5 положении.
Биосинтез РНК происходит под влиянием фермента РНК-полимеразы. РНК-полимераза E. coli имеет 5 типов белков, из которых 2 белка – альфа (α), 1 белок – бэта (β), еще один бэта' (β’), необычный белок сигма (σ).
αи β везде одинаковы, различаются по σ-фактору. Именно σ-фактор находит свой ген, с которого РНК-полимераза начнет считывать информацию, т. е. сколько генов, столько σ-факторов.
У эукариот известно 3 типа РНК-полимераз, наиболее активна РНК-полимераза-I, она находится в ядрышках ядра клетки, с ее помощью образуется рРНК и рибосомы. Ее количество 5070%.
РНК-полимераза-II (в ядре) - 20-40%, с ее помощью образуются предшественники иРНК. Их стали называть гетерогенные ядерные РНК. В дальнейшем она модифицируется и превращается в иРНК.
РНК-полимераза-III (в нуклеоплазме) - синтез тРНК. Этапы транскрипции:
1)инициация
2)элонгация
3)терминация
1) Инициация начинается с промотора (начало гена), который располагается на 5'-конце и заканчивается на терминаторе - 3'-конец. Транскриптон - современное понятие гена (5'-3' промежуток). В начале РНК-полимераза распознает промотор, состоящий из 41-44 нуклеотидов. Транскрипция происходит в направлении 5'-3' и все нуклеотиды, лежащие справа от стартовой точки обозначаются +1, +2, ...,+n влево -1,-2,...-n. РНК-полимераза садится на область от -20 до +20, минимальный участок связывания состоит из 12 нуклеотидов, во всех промоторах есть характерные
1
последовательности, т. е. они консервативны. Эти последовательности - сигналы для РНКполимеразы. Стартовая точка всегда представлена пурином (в основном, А), сразу влево от нее есть 6-9 нуклеотидов (ТАТААТ) - ящик Прибнова. Такого рода участки характерны тем, что между Т и А две связи, а между Г и Ц три связи, поэтому скопление Т и А в одном месте позволяет его легко разорвать, сделать этот участок одноцепочечным. Как правило, эти места являются началом синтеза. Считается, что ящик Прибнова нужен для ориентирования РНК-полимеразы, чтобы синтез шел в направлении 5'-3'.
Близкие по составу последовательности располагаются в области, которая называется "участок -35". Здесь 9 нуклеотидов такого же состава, это район распознавания, сюда присоединяются σ-факторы, именно в этом месте он распознает свой ген. Но есть промоторы, которые не могут прикрепить РНК-полимеразу без особых белков (САР и СRР). σ-фактор распознает участок от -10 до -35.
У эукариот подробно изучены промоторы, взаимодействующие с рНК-полимеразой-II. Здесь 3 похожих друг на друга участка. Они располагаются 1) в стартовой точке, 2) -25, 3) -75. Стартовый нуклеотид А. С боков он флангирован пиримидиновыми основаниями (Т и Ц). На расстоянии 19-27 нуклеотидов влево от этого участка располагаются 7 нуклеотидов (ТАТАА). Это место - ТАТААящик (ящик Хогнеса (Гольдберга-Хогнеса)). Еще левее оттуда в -70 - -80 находится последовательность ЦААТ - ЦААТ-ящик. Предполагается, что ТАТАА-ящик контролирует выбор стартового кодона, а ЦААТ-ящик - первичную связь ДНК и РНК-полимеразы.
2)В качестве предшественников роста цепи РНК необходимы рибонуклеозид-3-фосфаты. Рост начинается на 3'-конце, присоединяется рибонуклеозид-монофосфат и высвобождается дифосфат, при этом выделяется энергия для синтеза. У эукариот синтез происходит в пределах 1 гена, а у прокариот может охватывать несколько генов. Образуется длинная РНК, полицистронная РНК.
3)Она заканчивается на специфических последовательностях, кроме того, существует рофактор, который прикрепляется к 5'-концу растущей цепи РНК, как бы догоняя РНК-полимеразу. Когда РНК-полимераза застревает на терминирующих кодонах, ρ-фактор сбрасывает иРНК, при этом на конце иРНК имеются палиндромы, которые способны замыкаться в виде шпильки сами на себя.
2
Лекция 3. ГЕНЕТИЧЕСКИЙ КОД. БИОСИНТЕЗ БЕЛКА. ТРАНСЛЯЦИЯ.
Впервые 2 английских ученых Бидл и Татум доказали, что на основе молекулы ДНК происходит синтез полипептидной белковой молекулы и вывели формулу 1 ген = 1 полипептидная молекула. Позднее это оказалось не совсем так, потому что много генов определяют не белок, а, к примеру, тРНК.
Закономерно встал вопрос: как на основе ДНК информация идет на белковые молекулы? Впервые американец русского происхождения Гамов вывел закономерность: 4 нуклеотида определяют 20 аминокислот. Если 1 нуклеотид будет кодировать 1 аминокислоту (код моноплетен), то можно зашифровать только 4 аминокислоты (4^1=4). Если 2 нуклеотида определяют 1 аминокислоту (код диплетен), то можно зашифровать 16 аминокислот (4^2=16). Если код триплетен, то можно зашифровать 64 аминокислоты (4^3=64).
Впервые американский ученый Нирренберг синтезировал искусственную нуклеиновую кислоту. Она состояла только из У, т. н. полиуридиновая кислота. И в искусственных условиях ему удалось на ее основе получить полипептидную молекулу, которая состояла только из фенилаланина (1 аминокислота). Было принято решение, что УУУ определяет в белке аминокислоту фенилаланин. На ней же удалось получить все 64 варианта кодонов и определить, какую аминокислоту эти кодоны определяют. Были ведены общие свойства генетического кода:
1)генетический код универсален, практически одинаков для всех живых организмов;
2)код триплетен (1 аминокислота = 3 нуклеотида);
3)код не перекрывается (коды идут линейно друг за другом, и нуклеотид из одного кода используется другим);
4)нет разделительных знаков между триплетами;
5)код отличается коллинеарностью (совпадением кодонов иРНК и аминокислот в белке);
6)код является вырожденным, т. е. избыточным, поскольку 64 кодона определяют 20 аминокислот, то некоторые аминокислоты (большинство) имеют 2-3 (иногда даже 4) кодона;
7)из 3 нуклеотидов в кодоне главными являются первые 2, третий может варьировать;
8)в среднем, каждая аминокислота кодируется 3 триплетами;
9)частота использования различных кодонов может быть видоспецифичной, т. е. по частоте кодонов виды могут различаться;
10)инициирование кодонами, т. е. кодонами в начале молекулы ДНК являются УАГ и ГУГ. ОНи указывают начало синтеза полипептидной цепи, а перед ними еще стоит регуляторная часть, с которой полипептидная цепь не синтезируется.
Первая аминокислота - метиамин или формилметиамин. В конце - терминирующие нонсенскодоны, они не определяют никакую аминокислоту, а определяют точку конца информации (УАА, УГА, УАГ). За последним стоп-кодоном на 3'-конце иРНК располагаются некодирующие белок последовательности - трейлеры. В состав начального лидерного участка входит последовательность АГГ АГГ, она служит сигналом начала трансляции с иРНК. Ее стали называть последовательностью Шайн-Далгарно. Эта последовательность располагается на 5-7 нуклеотидов левее стартового кодона, она обеспечивает присоединение иРНК к рибосоме.
Имеются существенные отличия РНК прокариот и эукариот. У эукариот РНК более стабильна, это связано с процессами модификации незрелой иРНК. Сразу же после синтеза иРНК на 5'-головном конце образуется т. н. колпачок - кэп. Кэп представляет из себя гуанозин, у которого в 7 положении имеется метильная группа - 7-метилгуанозин. Кроме того, метилируется рибоза. Это формирует 2-й или даже 3-й кэп. Кроме того, к 3'-концу хвостовой части ферментами присоединяется до 200 адениловых остатков. Этот адениловый хвост не кодирует белок. После этого РНК покидает ядро, при этом число адениловых остатков может сокращаться. Установлено, что такие модификации (кэп и хвост) существенно стабилизируют молекулу, и уже клеточные нуклеазы не разрушают эту РНК. У прокариот это отсутствует и после 1 цикла синтеза белка РНК разрушается нуклеазами.
Когда ученые впервые определили, что количество нуклеотидов в гене и количество нуклеотидов в РНК не совпадают, было определено, что иРНК во много раз короче, чем ген. Было установлено, что в гене имеются незначимые участки, их назвали "интроны"; и значимые участки, которые несу информацию о белке, их назвали "экзоны". Кроме этого, в гене есть спейсеры – регуляторные участки. Оказалось, что из незрелой РНК происходит вырезание интронов и
1
спейсеров, это явление было названо сплайсингом, а все преобразование незрелой РНК в зрелую, образование кэпов, аденилового хвоста, вырезание интронов и сшивание экзонов называется процессингом. Вырезание идет по правилу Шамбона, согласно которому интроны всегда начинаются ГУ и заканчиваются АГ.
МЕХАНИЗМ ТРАНСЛЯЦИИ.
Сразу после опубликования работы Уотсона и Крика о структуре молекулы ДНК были высказаны разные идеи о матричном синтезе белка, они предполагали взаимодействие РНК и белковой молекулы, но эта идея была отвергнута. И впервые Френсис Крик пришел к выводу, что прямого контакта между РНК и синтезируемой полипептидной молекулой нет. Он выдвинул адаптерную гипотезу, т. е. должны быть адаптерные молекулы, которые с одной стороны должны соединяться с аминокислотой, а с другой - узнавать кодоны на иРНК.
Впервые американский ученый Хогленд обнаружил, что в клетке аминокислоты соединены с низкомолекулярной РНК, при этом было установлено, что это соединение обусловлено ферментом аминоацил-тРНК-синтетазами (АРСазы).
Согласно адаптерной гипотезе, функции адаптера осуществляются в 2 этапа: 1) трансляция I
а) активация аминокислоты; б)аминоацилирование - соединение тРНК с аминокислотой при посредстве АРСазы
2) трансляция II
а) перенос аминокислоты с помощью тРНК к кодонам иРНК б) опознание кодона тРНК тРНК для этого должна обладать следующими сайтами:
1.сайтом, специфичным АРСазе
2.сайтом связывания с кодоном антикодона тРНК
3.сайтом связывания с рибосомой
4.сайтом взаимодействия с различными факторами трансляции II
Таким образом, число тРНК и АРСаз в клетке должно быть не менее 20. Нет прямой связи между тРНК и аминокислотой. Свободная тРНК и аминокислота сцеплены с помощью АРСазы.
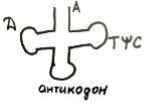
тРНК по своему строению напоминает трилистник клевера, при этом у нее есть акцепторный стебель, здесь на 3'-конце имеется кодон А АЦЦ, справа ТYC [ти пси цэ], дигидроуридиновая петля, антикодон внизу.
Впервые тРНК аланиновой аминокислоты была расшифрована Робертом Холли, а затем были расшифрованы и другие тРНК. В настоящее время известно более 300.
тРНК имеет 4 двуцепочечных и 5 одноцепочечных участков, в ней имеются т. н. минорные азотистые основания (дигидроуридин, псевдоуридин, метилинозин, метилурацил). Они не могут образовать двуцепочечные участки и содержатся в одноцепочечных.
Антикодирующая петля состоит из 7 нуклеотидов, с помощью которых тРНК взаимодействует с кодоном иРНК. Дигидроуридиновая петля способствует формированию третичной структуы. Двуцепочечные структуры делят 2 витка. Как ДНК и тРНК имеет Г-образную форму. Найдено много тРНК, несущих 1 аминокислоту, но иеющих разные антикодоны. Это изоакцепторные тРНК. Физическая конфигурация АРСаз повторяет конфигурацию родственной тРНК.
Антикодон тРНК узнает кодон иРНК путем спаривания азотистых оснований, например, дрожжевой аланин тРНК может связываться с 3 кодонами (ГЦУ, ГЦЦ и ГЦА).
Впервые Френсис Крик предположил, что основными являются только первые 2 нуклеотида, а относительно третьего возможна свобода.
2
Кодоны, различающиеся по одному из первых двух оснований должны узнаваться различными тРНК. Например, УУА и ЦУА оба кодируют лейцин, но считываются разными тРНК. Первое основание антикодона определяет, считывает ли данная тРНК один, два или три типа кодонов. Если это Ц или А, узнается только один кодон, если У и Г, то 2 кодона. Если необычный инозин, то 3 кодона.
Т. о., причина вырожденности кода - неоднозначность спаривания третьего кодона.
Весь процесс биосинтеза белка происходит на рибосомах. Рибосома - это сложная структура диаметром 2 нм, коэффициент седиминтации осаждения 50s, для маленькой - 30s, у прокариот 50s состоит из 34 видов белков, двух молекул РНК, одна 23s, а другая 25s.Маленькая частица имеет 21 белок и 16 РНК. У эукариот рибосомы крупнее. Крупная - 60S, имеет з РНК, 28, 7 и 5S, а малая (40S) имеет 18S-РНК. Все эти компоненты самособираемы. Если биосинтез не происходит, то большая и малая субчастицы плавают в цитоплазме. Отдельно при реконструкции 30s-субчастицы были сделаны выводы:
1)16s-РНК необходима для сборки и функционирования рибосомы
2)она видоспецифична
3)из 80 белков 6 строго необходимы для сборки.
Опыты Говарда-Динциса показали, что считываются от аминного конца к карбонильному, при этом на иРНК через каждые 8 нуклеотидов сидит новая рибосома, и те рибосомы, которые близко находятся к РНК-полимеразе синтезируют крупные полипептидные молекулы, а которые на конце - малые, т. е. рибосомы ближе к 5'-концу несут самые короткие полипептидные цепи, а к 3'-концу - самые длинные.
После биосинтеза субчастицы распадаются. Первый трансляционный кодон всегда находится на расстоянии не менее 25 нуклеотидов от 5'-конца.
3

Лекция 4. СТРУКТУРНАЯ ОРГАНИЗАЦИЯ ХРОМОСОМ ЧЕЛОВЕКА.
Впервые хромосомы человека в делящихся клетках обнаружил немецкий ученый Флемминг при окрашивании клеток основными красителями в 1880 г. Он их описал как хроматиновые окрашивающиеся тела. Термин «хромосома» был предложен другим немецким ученым Вальдайгером в 1888 г. Было установлено, что при делении клетки возникают палочкообразные хроматиновые тела, которые потом расходятся при
делении к разным полюсам клетки. При изучении морфологии хромосом были выявлены следующие особенности: хромосомы отличаются по размерам и по конфигурации, кроме того, в разные стадии деления хромосомы могут состоять из 1 или 2 хроматид.
У них выделяют короткое и длинное плечи, первичную перетяжку, внутри которой находится центромера, а внутри центромеры располагается кинетохорд (область, с помощью которой половинки хромосом (хроматиды) могут двигаться к разным полюсам клетки при участии особых моторных белков).
Морфологически выделяют три типа хромосом:
1)если центромера находится приблизительно посередине, короткое плечо не менее 40%,то такую хромосому называют метацентрической;
2)если короткие плечи составляют 20-45% длины хромосомы, то эти хромосомы называют субметацентрическими; 3)когда короткие плечи менее 25%, их называют акроцентрическими.
Некоторые ученые выделяют также телоцентрические хромосомы  Морфологически хромосомы могут иметь вторичную перетяжку. Например, у
Морфологически хромосомы могут иметь вторичную перетяжку. Например, у
человека 9 хромосома имеет вторичную перетяжку. Кроме того, некоторые хромосомы имеют образования, которые называют спутниками или сотелитами, чаще всего концентрические хромосомы имеют рожки на тонкой нити (как бы кусочки хромосом).
Когда смотришь на хромосомы человека под микроскопом, то видно, что спутничные хромосомы зачастую обращены друг к другу. Оказалось, что в спутниках человеческих хромосом сосредоточены рибосомальные гены, здесь синтезируется рибосомальная РНК, и в этом месте образуется ядрышко ядра, отсюда выходят в цитоплазму рибосомы. Эти хромосомы сцеплены материалом ядрышка. Вот почему они так выглядят под микроскопом.
Анализ молекулярной структуры хромосом показал, что хромосомы состоят, в основном, из ДНК и особых сильноосновных белков - гистонов (ДНК и гистонов примерно поровну в ядре клетки). Кроме того, здесь присутствуют в небольшом количестве негистонные белки, РНК и, в малом количестве, липиды.
Из хромосом выделено 5 фракций гистонов: Н1, Н2а,Н2в, Н3, Н4. Все гистоны находятся примерно в равном количестве (кроме Н1,его в 2 раза меньше, чем остальных). Гистоны образуют т. н. нуклеосомы, здесь 8 гистонов формируют "шайбу" - нуклеосому. ДНК делает 1,75 витка вокруг этой нуклеосомы, с ней контактируетирует 146 нуклеотидов.
Между двумя нуклеосомами ДНК свободна, этот участок свободной ДНК называется линкер. Как оказалось, гистон Н1 располагается над этой структурой, с одной стороны он цепляется за 1 нуклеосому, с другой - за другую нуклеосому.
Формируется т.н. нуклеосомная нить. Под влиянием особого метагенного сигнала гистон Н1 начинает сжиматься и подтягивать одну нуклеосому к другой. Нуклеосомная нить имеет толщину 11 нм.
1
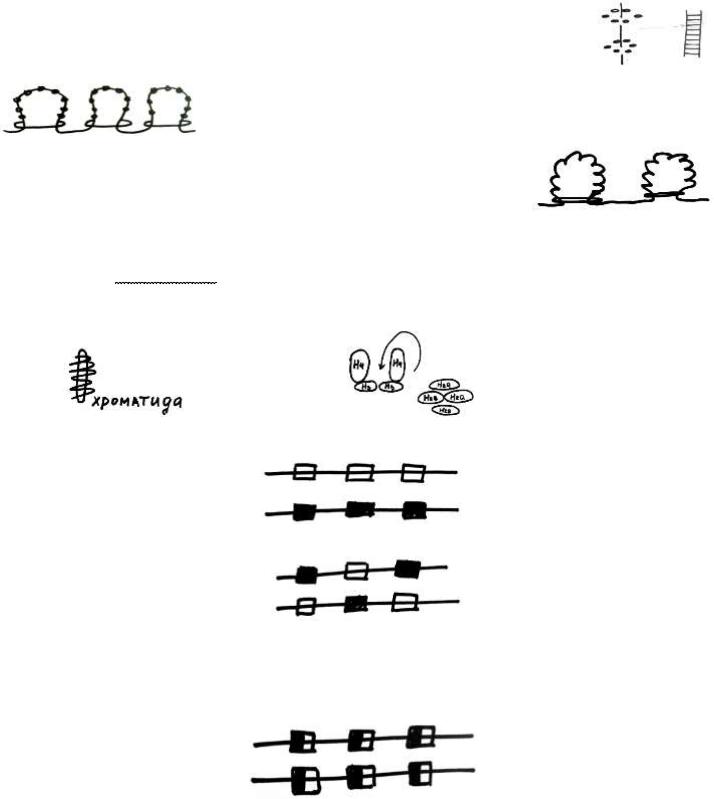
Если процессы конденсации хроматина продолжаются дальше, то формируется т.н соленоидная нить (6 нуклеосом выстраиваются в одной плоскости, образуется соленоидная нить). Толщина нити около 30 нм.
Процесс конденсации продолжается, и соленоидная нить начинает образовывать петельные домены. И сами петли удерживаются в таком состоянии специальными белками-замками, это негистонные белки.
Упаковка продолжается, образуются петельные домены второго порядка, это огромные петли по 80000 нуклеотидов, и они включают в себя петли меньшего порядка, и, опять же, все это удерживается с помощью специальных негистоннных белков.
Нить из петельных доменов 1го порядка толщиной 300 нм, а нить из петельных доменов 2го порядка имеет толщину около 700 нм. И наконец, в хроматиде хромосомы внутри имеется стержень, состоящий из металлопротеина, и здесь формируются гигантские петли, свободные концы которых зафиксированы стержнем, эту структуру назвали ламповой щеткой. При репликации молекулы ДНК было установлено, что нуклеосомы могут распадаться на полунуклеосомы.
Когда образуется дочерняя нить ДНК, возможны следующие ситуации:
1)старая материнская нить имеет старые нуклеосомы, а новая образует новые нуклеосомы;
2)обе нити содержат и старые нуклеосомы, и новые нуклеосомы;
3)нуклеосомы распадаются на полунуклеосомы, и затем формируются гибридные хромосомы: кусочек старой – кусочек новой (большинство ученых полагает так, хотя есть случаи с полным разваливанием хромосом на составные части (гистоны), с последующей сборкой из старых и новых гистонов).
Молекула ДНК в ядре может находиться в двух состояниях: плотном компактизированном состоянии, и в этом случае гены в этой ДНК не функционируют, такие районы плотно окрашиваются основными красителями, их стали называть гетерохроматиновые районы. Различают 2 типа гетерохроматиновых районов:
1) факультативный гетерохроматин, т. е. эти районы могут быть гетерохроматиновыми или нет, функционировать или не функционировать. Например, этот участок функционирует в эмбрионе, но не функционирует у старых людей. Или же эти районы функционируют в клетках печени, но не функционируют в мышечных клетках. Те районы, что не работают, заблокированы;
2
2) конститутивный гетерохроматин. Например, Х-хромосома у женщин. У женщин 2 Х-хромосомы, и они обе функционируют только в первом триместре беременности у плода, как только пол определится в женскую сторону, одна из хромосом блокируется. Функционирующие участки максимально раскручены, деспирализованы, часто одноцепочечны, здесь происходит транскрипция, их называют эухроматиновми районами. Они очень слабо окрашиваются основными красителями.
Т. о. процессы спирализации и деспирализации происходят постоянно в ядерном аппарате клеток.
Выделяют 4 типа ДНК по степени спирализации:
Тип ДНК Направление спирали Число пар нуклеотидов Диаметр спирали, нм на 1 виток
А ДНК |
правое |
10,7 |
2,3 |
В ДНК |
правое |
10 |
2 |
С ДНК |
правое |
9,3 |
1,9 |
Z ДНК |
левое |
12 |
1,8 |
Левоспиральность образуется в процессе транскрипции в тех местах, где правая спираль, и если в этом месте начинается закручивание в левую сторону, то нити разойдутся друг от друга, образуется участок одноцепочечной ДНК, и в этом месте начнется транскрипция, т. о. действует ДНК-гираза или топоизомераза, которая образует спиральность.
РЕПРОДУКТИВНАЯ ОРГАНИЗАЦИЯ КЛЕТКИ.
В организме человека деление или пролиферация обеспечивает:
1)рост организма;
2)регенерацию (физиологическую и патологическую);
3)воспроизведение потомства;
4)эмбриогенез.
Различают жизненный цикл клетки - этот весь период жизни клетки с момента возникновения до гибели. И второе, митотический цикл клетки - это цикл, связанный только с делением клетки. Все клетки разделяют на три популяции:
1)стабильная - клетки не способны к делению, наибольшая продолжительность жизни (нервные клетки, клетки сердца);
2)растущая - долгоживущие, но способные к делению (железы, печень);
3)обновляющиеся:
а) стволовые, недифференцированные, они только делятся, находятся в митотическом цикле; б) дифференцированные, короткоживущие, неспособные к делению (эритроциты)
Т. о. в жизненном цикле клеток стабильных популяций и дифференцированных клеток полностью отсутствует митотический цикл. В растущей популяции может присутствовать митоз.
Главная цель митоза или кариокинеза - это равное распределение генетического материала меду дочерними клетками. В митотическом цикле различают две стадии:
1)интерфаза
2)собственно митоз
Интерфаза подразделяется на три периода:
1)предсинтетический или G1 -период. Здесь активно синтезируется РНК, образуются структурные белки и белки - ферменты, накапливаются предшественники ДНК и энергетические молекулы АТФ.
2)синтетический или S-период. Здесь образуется дочерняя нить ДНК, происходит репликация, по существу, каждая хромосома становится двухроматидной, и формируется центромера - клеточный центр делится на две центриоли.
3
3) постсинтетический или G2-период. Здесь клетка накапливает белки, необходимые для деления (очень важный белок тубулин, он образует нити веретена деления, по которым, как по рельсам, хромосомы начнут отходить к разным полюсам), кроме этого, моторные белки, которые будут двигать хромосому (хинезин и динеин). Более того, накапливаются молекулы ГТФ (гуатинтрифосфат) и АТФ (аденозинтрифосфат), активно происходит синтез иРНК.
Всобственно митозе различают 4 фазы: профаза, метафаза, анафаза и телофаза.
1)Профаза. В профазе начинается процесс конденсации хромосом, под влиянием митотического сигнала особого белка Н1 гистон начинает сжимать нуклеосомы друг с другом, и происходит резкое укорочение хромосом в несколько тысяч раз, это в дальнейшем позволит свободно расходиться хромосомам, не переплетаясь друг с другом, кроме того, здесь исчезает ядрышко, начинает растворяться ядерная оболочка. Центриоли клеточного центра начинают движение к разным полюсам, при этом между ними формируется тубулиновый аппарат деления - особые нити в виде трубочек диаметром около 25 нм, пустых внутри, внутренний диаметр около 15 нм, в оболочке трубочки 13 протофиломентов. Один конец («+»-конец) постоянно присоединяет следующую молекулу тубулина, от другого конца («-»-конец) наоборот отщепляются молекулы тубулина. Весь этот процесс происходит при участи гуанозинтрифосфата (ГТФ) и ионов магния. При движении центросом к разным полюсам начинают формироваться дополнительные хромосомные нити. Хромосомные нити подходят к хромосоме, соединяются с кинетохором центромеры хромосомы, причем, с одного полюса к одной хроматиде, а с другого полюса - к другой хроматиде, и они начинают толкать хромосомы к центру клетки. И клетка вступает в метафазу
2)Метафаза. Хромосомы образуют экваториальную метафазную пластинку. На этой стадии особенно отчетливо видны хромосомы. Их зачастую изучают на этой стадии. Резко изменяется вязкость цитоплазмы, это, по мнению ученых, вызывает разделение центромер, и хроматиды хромосомы отходят друг от друга.
3)При этом к трубочкам присоединяются моторные белки гинеин и кинезин, они имеют головную и хвостовую части. В головной части содержится молекула АТФ. Эта часть соединяется с трубочкой, а хвостовая часть - с центромерой хроматиды, и хроматиды хромосом начинают движение к разным полюсам под влиянием моторных белков, сама трубочка начинает деполимеризоваться, укорачиваться. Эта стадия называется анафазой;
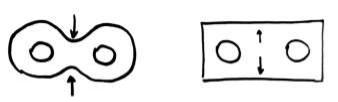
4)Когда хроматиды достигают противоположных полюсов, наступает стадия телофазы. При этом вокруг отошедших однохроматидных хромосом начинает формироваться ядерная оболочка, а между дочерними ядрами происходит образование перегородки. Если в животной клетке этот процесс происходит со сжиманием, приобретается гантелевидная форма, то в растительной клетке процесс начинается в центре и двигается к оболочке.
Т. о. в результате митоза образуются две генетически абсолютно одинаковые клетки. Хромосомы в митотическом цикле могут быть однохроматидными и двухроматидными.
Каждая хроматида - двуцепочечная молекула ДНК, обычно обозначают 2с.
Однохроматидные хромосомы в анафазе, телофазе и в G1 период. Двухроматидные в конце S- периода, профазе и метафазе. Обычно введены такие обозначения: 2с и 4с. Кроме того, введено обозначение 2n и n, все соматические клетки диплоидны.
4

Лекция 5. МЕТОДИЧЕСКИЕ ОСОБЕННОСТИ ХРОМОСОМНОГО НАБОРА ЧЕЛОВЕКА. ПРИГОТОВЛЕНИЕ ХРОМОСОМ И ИХ ИДЕНТИФИКАЦИЯ.
Первые хромосомные препараты приготовил немецкий ученый Флемминг из роговицы глаза умершего человека. Одновременно другой немецкий ученый Гертвиг изготовил препараты из семенников человека. Когда было подсчитано число хромосом, то они насчитали 26-30 хроматидных тел. Они готовили препараты гистологическим методом, т.е. ткань человека через проводку, спирты разной концентрации и спирт + парафин заключали в твердые парафиновые блоки. А затем резали на микротоме толщиной в несколько микрон. Но поскольку крупные хромосомы достигают размеров около 10 микрон, то неминуемо некоторые хромосомы разрезались. Кроме того, хромосомы плотно лежали друг около друга, и отделить их было очень сложно. В 30е гг. прошлого века самой мощной цитогенетической школой была российская, и наши ученые (Навашин, Андрес, Живаго) так же попытались изготовить хромосомы человека. Подсчитав число хромосом, пришли к выводу, что их 48. Методика была несовершенна, но было уже известно, что у человекообразных обезьян 48 хромосом.
1879 г. - Арнольд впервые визуализировал хромосомы.
Впервые в 1956 г. два шведских ученых Тио и Леван применили новую оригинальную методику. Они в исследовании взяли эмбриональную ткань плода человека. Механически измельчив ее, они с помощью фермента трипсина, который растворяет межклеточные взаимодействия, получили взвесь отдельных клеток. Они помещают эти клетки в питательную среду, состоящую из 199 компонентов - это все известные аминокислоты, нуклеотиды, витамины, соли и, кроме того, добавлялась сыворотка крови плода человека. В этом бульоне клетки начали размножаться. Первое деление было примерно через 1 сутки. Чтобы остановить митозы они добавили колхицин (алкалоид), который растворял нити веретена деления, и хроматиды хромосом не могли в анафазе отходить к разным полюсам. Хромосомы оставались двухроматидными, т. е. все фиксировалось на стадии метафазы. Далее, для того, чтобы хромосомы отошли друг от друга из клубка, они использовали гипотонический раствор солей. Клетки набухали. Хромосомы отходили друг от друга. Затем добавлялся фиксатор, который закреплял это состояние (фиксатор Карнуа, состоящий из абсолютного спирта и ледяной уксусной кислоты). Клетки наполнялись спиртом. Затем их охлаждали, помещали на предметное стекло каплю и поджигали. Набухшие клетки лопались, взрывались, хромосомы разбрасывались и приваривались к предметному стеклу. После окраски основными красителями Тио и Леван доказывают, что у человека 46 хромосом, их отлично видно, размеры от 10 до 2 микрон, они морфологически отличаются друг от друга - есть метацентрические хромосомы (центромера посередине), есть акроцентрические (в крайнем верхнем положении) и субметацентрические. Эта методика стала очень быстро распространяться, но возникла необходимость создания номенклатуры хромосом человека (кариотипа).
Понятие кариотипа ввел Левицкий, под ним он понимал упорядоченный набор хромосом в связи
с:
1)гомологичностью хромосом (одна от матери, другая от отца);
2)морфологическими особенностями (мета-, субмета-, акроцентрические хромосомы);
3)размерами;
4)положением центромеры.
В1961 г. в Денвере, США, собирается первая конференция по хромосомам человека, где ученые, занимающиеся изучением хромосом, привели первую денверскую классификацию хромосом человека. Согласно этой классификации, каждая хромосома получила свой номер. Были выделены отдельно половые хромосомы человека (хх (ж) и ху (м)). Крупные хромосомы - в начале классификации, мелкие - в конце. Но англ. ученый Патау подверг критике эту классификацию, он пришел к выводу, что некоторые хромосомы по обычным морфологическим критериям неотличимы друг от друга, и поэтому в 1963 году на лондонской конференции было принято решение разделять хромосомы по группам. Всего было выделено 5 групп.
Группа хромосомы |
Номер хромосомы |
Длина в микронах |
Центромерный индекс |
|
|
|
1 |

|
|
|
(отношение |
короткого |
|
|
|
плеча к |
длине всей |
|
|
|
хромосомы, %) |
|
А |
1 |
8,4 |
48,4 |
|
|
2 |
8,0 |
39,2 |
|
В |
3 |
6,8 |
46,9 |
|
|
4 |
6,3 |
29,1 |
|
|
5 |
6,1 |
29,3 |
|
С |
6 |
5,9 |
39,1 |
|
|
7 |
5,4 |
39,1 |
|
|
Х |
5,1 |
40,1 |
|
|
8 |
4,9 |
34,1 |
|
|
9 |
4,8 |
35,4 |
|
|
10 |
4,6 |
33,9 |
|
|
11 |
4,6 |
40,1 |
|
|
12 |
4,7 |
30,2 |
|
D |
13 |
3,7 |
17,1 |
|
|
14 |
3,6 |
18,7 |
|
|
15 |
3,5 |
20,3 |
|
E |
16 |
3,4 |
41,3 |
|
|
17 |
3,3 |
33, |
|
|
18 |
2, |
30,9 |
|
F |
19 |
2,7 |
46,5 |
|
|
20 |
2,6 |
45,5 |
|
G |
21 |
1,9 |
30,9 |
|
|
22 |
1,0 |
30,5 |
|
|
Y |
1,2 |
27,2 |
|
В кариотипе человека к крупным хромосомам относят 1 и 3; 16 – средняя; 19,20 - мелкие. К акроцентрическим хромосомам относятся D-группа 13,14,15, средние по размерам, имеют спутники (на коротких плечах особые образования, в которых находятся гены рРНК). Здесь формируется 18s-РНК и 28s-РНК, которые строго необходимы в транскрипции и трансляции белка, поэтому эти хромосомы ориентированы спутниками друг на друга и находятся близко. К ним относят 21,22. Остальные хромосомы субметацентрические разных размеров.
Дальнейшие исследования хромосом показали, что необходимы дополнительные критерии, чтобы было возможно идентифицировать все хромосомы, усилия ученых были направлены на выявление гетерохроматиновых участков хромосом. Оказалось, что хромосомы по своей длине имеют функционально активные и неактивные участки. Неактивные участки плотно упаковываются, они окрашиваются интенсивнее основными красителями. Здесь гены не работают, а светлые участки - эухроматинове зоны, где активно функционируют гены. Кроме того, оказалось, что по концентрации А- Т, Г-Ц разные районы хромосом отличаются, а между А и Т две связи, которые легко рвутся. Таким действием обладают флуоресцентные красители, они могут разорвать связи между А и Т и встроиться между ними, это явление называется интраколяция.
В 1970 г. шведский ученый Касперсон использовал акрихин-иприт (Q-метод окраски). Зона, где много А-Т поглощала много красителя, и при облучении ультрафиолетом эти места светились, четко видна была исчерченность. Недостаток - быстрое обесцвечивание.
Группа японских ученых предложила другой метод: они обрабатывали хромосомные пластинки трипсином, который разрушал белковую часть хромосом, а после это окрашивали красителем Гимзе (эозин красный и азур-2 синий). Наблюдалось окрашивание полосками, которые стали называть бендами, и в зависимости от того, на какой стадии мы анализируем клетку, количество бендов в хромосоме может
2

изменяться: чем ближе к профазе, тем больше бендов (например, первая хромосома - до 850 бендов), а чем ближе к анафазе, тем меньше бендов (около 200). Анализируют профатические хромосомы.
В 1971 г в Париже состоялась конференция по хромосомам, где принимается номенклатура, согласно которой все хромосомы человека были идентифицированы по 3 критериям :
1)длина,
2)центромерный индекс ,
3)расположение бендов в хромосоме.
Существовало направление евгеники – ставившая целью улучшение человеческой "породы". 30е
ггXX в. стерилизация людей с "плохой наследственностью". Лисенковщина (направление возглавил Лисенков).
Пинкел и Каллиониэме обнаружили, что хромосомы по некоторым районам ДНК отличаются, изготовили зонды для каждой хромосомы, к которым они прикрепили разноцветные красители, после обработки на компьютере было видно, что хромосомы отличаются.
А-группа хромосом 1-3 метацентрические самые большие, В-группа 4,5 субметацентрические, крупные, но верхние плечи короче, чем у вторых хромосом. С-группа с 6 по 12 средние субметацентрики. D-группа акроцентрики средние со спутниками, Е-группа 19,18 маленькие, 16 метацентрик, 17,18 субметацентрические, F-группа очень маленькие метацентрики, G-группа маленькие акроцентрики со спутниками, а Y-хромосома - маленький акроцентрик без спутников.
Основные окраски: 1) Q-способ
2) G-способ с помощью Гимза и трипсином
3) окрашивается гетерохроматин в основном возле центромеры Гетерохроматин - плотный компактный район хромосомы, обычно расположен в области
центромеры, в у-хромосоме есть, но это конститутивный (постоянный) гетерохроматин. Существует факультативный гетерохроматин в одной из х-хромосом, выглядит как тельце Барра, черное пятно на ядерной оболочка. Англ. ученая Лайон объяснила это так: поскольку у женщин генетического материала в клетке больше, чем у мужчин, поскольку х-хромосома больше, то обе х-хромосомы функционируют только до закладки пола, дальше одна из них инактивируется, причем, если одна из х-хромосом несет аномальный ген, то часть клеток будет нормальной, а часть патологическими. Мозаичность. Компенсаторная часть.
При дифференциальной окраске введены обозначения. Первая цифра - номер хромосомы; потом р-короткое плечо; q - длинное плечо; третий символ - цифра номер района, в который входит данный бенд; номер бенда.
Концевые участки хромосом называются теломеры, они содержат специализированные последовательности в виде ТТА ГГГ много раз повторяющиеся, их функция очень важна. С одной стороны они защищают от прикрепления хромосом друг к другу, а с другой они определяют нашу продолжительность жизни. В 70 гг. американский ученый Хакель обнаружил, что когда культивируешь в питательной среде эмбриональную ткань человека, то как бы ты не старался, больше 60 делений не выйдет, причем от эмбриона в ср 50 делений, от 18 - 20-25 делений, с возрастом уменьшается.
Вся жизнь человека - около 50 делений (короткожитель), 60 - долгожитель. Однако было непонятно, с чем это связано. Еще в 1971 Оломников впервые высказал мнение, что все дело в теломерах. Гипотеза, что при каждом делении отрезается участок теломеры пока деление не прекращается, но доказать это не смог он. Это доказали американские ученые Блэкбер, Грейде, Шостен, они установили, что отрезается примерно 10 нуклеотидов. В теломере порядка 500. В 2009 они получили Нобелевскую премию.
Ядрышкообразующие районы красят азотокислым серебром, они локализованы в 13,14,15,21,22 хромосомах. Примерно в этих хромосомах находятся около 500 (200-700) копий генов рРНК, если 700 то у человека очень активный белковый синтез, он сам активен. Если 200 - наоборот.
3
Лекция 6. МЕЙОЗ. ГАМЕТОГЕНЕЗ.
У диплоидных организмов, к которым относится и человек, возникает особое деление половых клеток, которое было названо мейоз. Цель мейоза:
1)уменьшить число хромосом с диплоидного до гаплоидного;
2)рекомбинация генетического материала.
Вотличие от митоза мейоз имеет две стадии деления:
1)редукция – уменьшительная;
2)эквационное деление – уравнительное.
Перед началом деления клетки диплоидные с двухроматидными хромосомами, т. е. они прошли G 1, S, G2 стадии интерфаз. В редукционном делении первая профаза (профаза I) состоит из 5 стадий:
1)лептонема/лептотема - стадия длинных нитей. Видны тонкие извитые нити, хромосомы на своей поверхности имеют утолщения (хромомеры), и хроматиды хромосом как бы цепляются друг за друга хромомерами. Они образуют клубок нитей, при этом свободные концы обычно направлены к клеточному центру. Особенно сильно конденсируются, уплотняются половые хромосомы X и Y.
2)зигонема, для этой стадии характерны установление связей-синапсов между гомологичными хромосомами. В результате этого эти двойные структуры из двух хромосом формируют биваленты. Х и Y хромосомы имеют очень маленькое сродство небольшими участками на концах хромосом и поэтому они образуют бивалент своими концами.
3)пахинема - наиболее длительная стадия по времени. Здесь происходит сильная конденсация хромосом, и каждый бивалент как бы распадается на 4 хроматиды, поэтому его здесь называют тетрадом. Процесс объединения или конъюгации хромосом продолжается, и в бивалентах хроматиды гомологичных хромосом начинают переплетаться друг с другом. Их называют хиазмы, т. к. напоминают греческую букву хи (χ).
4)диплонема - хромосомы резко укорачиваются, и происходит отталкивание хроматид друг от друга. В местах хиазм происходит обмен участками, это явление называется кроссинговер или рекомбинация генетического материала
5)диакинез. Здесь хиазмы сползают к концам хромосом - теломерам, и хромосомы удерживаются друг около друга своими концевыми участками. В это время происходит растворение ядерной оболочки, и центриоли клеточного центра начинают движение к разным полюсам, при этом они толкают хромосомы к экватору клетки. Далее к каждой двухроматидной хромосоме присоединяется только одна нить аппарата деления в районе кинетохора центромеры. Стадия, когда хромосомы оказываются на экваторе, называется метафаза I.
Этап анафаза I. Двухроматидные хромосомы начинают движение к разным полюсам, когда они достигнут полюса, это телофаза I. В телофазе клетки будут иметь уменьшенное вдвое число хромосом, т.е. оно будет равно n (1n). А поскольку каждая хромосома двухроматидна, то количество ДНК будет 2с. Затем короткий покоящийся период - интеркинез. Здесь нет синтеза ДНК. Клетка вступает в эквационное деление.
Профаза II. В ней наблюдается растворение ядерной оболочки, если она образовалась в телофазе, затем центриоли опять движутся к разным полюсам, хромосомы еще более конденсируются и начинают движение к экватору клетки.
Метафаза II. Хромосомы на экваторе клетки, но теперь к каждой хромосоме подходит 2 нити веретена - с одного полюса и с другого, и в конце метафазы центромера, сдерживающая хроматиды хромосом расщепляется, и наступает анафаза II. Половинки хромосом, хроматиды, начинают движение к разным полюсам. Когда они достигнут, будет телофаза II.
Таким образом, имеются следующие существенные отличия мейоза от митоза:
1)в мейозе два следующих друг за другом деления
2)в мейозе в профазе I наблюдается конъюгация гомологичных хромосом
3)в профазе I происходит рекомбинация генетического материала - кроссинговер
4)в телофазе I набор хромосом в мейозе гаплоидный, а в митозе диплоидный Значение мейоза:
1)поддержание постоянного числа хромосом, если бы не было этого явления, то в каждом поколении число хромосом удваивалось бы
1

2) рекомбинация приводит к тому, что количество вариантов генома у потомков одн ой пары родителей составляет 1012, т. е. из-за кроссинговера дети в одной семье генетически различны, и это имеет огромное эволюционное значение, позволяя работать естественному отбору.
ГАМЕТОГЕНЕЗ Гаметогенез - это процесс образования гамет (половых клеток: сперматогенез (м) и овогенез (ж)). У
человека способность к репродукции становится возможна с половой зрелостью. У девочек примерно в 14, у юношей примерно в 16. Репродуктивная возможность у женщин, как правило, до 40-45 лет, у мужчин может до глубокой старости. За весь репродуктивный период у женщин образуется 400 яйцеклеток. Чем старше женщина, тем длиннее период между редукционным и эквационным делениями, и это ведет к неправильному расхождению хромосом, поэтому у пожилых женщин велика вероятность рождения детей с несбалансированным числом хромосом. Такого явления нет у мужчин, но у пожилых мужчин, как свидетельствуют данные, увеличено потомство с доминантными генным мутациями.
Сперматогенез - образование сперматозоидов, протекает в семенных канальцах семенников. Наружный слой семенных канальцев образован диплоидными сперматогониями, они интенсивно делятся митотически, начиная от мужского эмбриона до половой зрелости, они проходят около 30 делений, где зона, в которой интенсивны митотические деления, называется зоной размножения. Часть сперматогоний вступает далее в зону роста, клетки резко увеличиваются в размерах, накапливают АТФ, РНК, белки, и эти клетки называют сперматоцитами первого порядка, они тоже диплоидны. Далее эти клетки вступают в зону созревания, которая располагается ближе к центру семенного канальца. Клетки вступают в редукционное деление мейоза. Из одного сперматоцита первого порядка образуются 2 сперматоцита второго порядка. Далее клетки вступают во второе эквационное деление, и образуется 4 сперматиды.
Овогенез (оогенез) протекает в яичнике, причем если мейотические деления у мужчин начинаются при половом созревании, то в женском организме мейоз начинается еще в женском эмбрионе примерно в возрасте 2-4 месяцев. Здесь вначале также формируется митозом овогонии, они диплоидны. Далее они увеличиваются в размерах и превращаются в овогонии 1 порядка. Затем идет редукционное деление мейоза, и здесь образуются 2 разные клетки: первая - овоцит 2 порядка с гаплоидным числом хромосом, и она забрала на себя практически всю цитоплазму и питательные вещества, а вторая - редукционная, маленькое тельце, она забрала половину ядерного материала. Далее клетки вступают во второе эквационное мейотическое деление, и история повторяется. Овоцит 2 порядка образует одну овотиду со всей цитоплазмой и одно редукционное тельце, которое содержит гаплоидный набор однохроматидных хромосом. Редукционное тельце тоже разделяется на 2 тельца. Т. о. один овоцит 1 порядка дает 1 овотиду и 3 редукционных тельца. Яйцеклетки имеют относительно крупные размеры (до 60 микрон), шарообразны, неподвижны, содержат питательные вещества, в основном, желток, и во время созревания их покрывает т. н. фолликулярный эпителий, который как бы защищает яйцеклетку. В яичники процесс образования половых клеток начинается. Можно различит кору и паренхиму яичника. В коре происходят митотические деления, а в паренхиме (3мес. возраст плода) находятся клетки в стадии лептонемы, зигонемы, в четырехмесячном возрасте есть те, что находятся в стадии пахинемы, в семимесячном возрасте клетки оказываются на стадии диплонемы и как бы застревают на этой стадии до полового созревания. Таких клеток у женщины около 400. У яйцеклеток различают первичную, вторичную, иногда третичную оболочки. Первичная оболочка внутренняя, формируется на стадии еще овоцита, она пронизана выростами фолликулярных клеток. Вторичная оболочка сформирована фолликулярными клетками, а третичная, например, у птиц, - скорлуповая оболочка, это продукт секреции желез яйцевода.
ЭМБРИОГЕНЕЗ.
Эмбриогенез - процесс образования, включающий в себя частично гаметогенез, процесс оплодотворения яйцеклетки, формирование плода и рождение. Прогенез - процесс формирования половых клеток в эмбриологии, развития. Развитие сперматозоидов происходит в семенниках и оно идет в несколько этапов. Примерно от спермогония до сперматозоида время составляет около 72 часов. За это время ядерный аппарат замещается, здесь гистоны замещаются на протамины, и ядро кристаллизуется. Клетка удлиняется, клеточный центр с ахроматиновым аппаратом, нитями ахроматинового аппарата, формирует хвостовую част ь
2
сперматозоида. Митохондрии дают энергию для движения, они перемещаются в шейку сперматозоида. В передней части образуется акросома. За счет аппарата Гольджи здесь скапливаются протеолитические ферменты, которые способны растворять оболочку яйцеклетки. Среднем в 3 мл спермы содержится 350 ион сперматозоидов. За 1,5-2 часа они достигают фаллопиевых труб, именно здесь, в фаллопиевых трубах, происходит процесс оплодотворения. Сперматозоиды сохраняют оплодотворяющую способность около 2 суток.
Эмбриогенез - это часть индивидуального развития организма, онтогенеза. Эмбриология изучает процессы развития человека от зачатия до рождения. Эмбриогенез продолжается 350 суток (10 лунных месяцев), и подразделяется на три периода:
1)1 неделя - начальный
2)зародышевый (2-8 нед)
3)плодный (с 9 нед. до рождения)
Вэмбриогенезе человека можно выделить следующие стадии:
1)оплодотворение
2)дробление зиготы - оплодотворенной яйцеклетки с образованием зародышевых листков (экто-, эндо- и мезодермы)
3)гаструляция - появление различных тканей
4)гистогенез и органогенез - появление органов
Среди разных живых организмов различают разные типы яйцеклеток:
1)изолецитальные яйцеклетки – маложелтковые (у млекопитающих и человека)
2)телолецитальные – многожелтковые (у птиц)
3) центролецитальные - желток находится в центре (насекомые) Эмбриогенез может быть:
1)личиночный, развивается с метаморфозами (амфибии - у них имеются временные, провизорные органы (у головастика лягушки есть хвост, жабры))
2)неличиночный (у птиц)
3)внутриутробный (у человека и плацентарных млекопитающих)
При этом различают следующие периоды эмбриогенеза:
1)проэмбриональный - начальный. Образование гамет. Здесь мейоз, редукция числа хромосом, образование яйцеклеток, их формирование;
2)эмбриональный / пренатальный. Начало проникновения сперматозоида в яйцеклетку, при этом различают 3 фазы;
а) дистантное взаимодействие сперматозоида и яйцеклетки, здесь важное значение имеют гормоны гиногамон и андрогамон. Эти два гормона могут активировать сперматозоид, или иногда подавить;
б) контактное взаимодействие половых клеток под влиянием сперматолизинов акросомы спермия, происходит объединение мембран сперматозоида и яйцеклетки;
в) проникновения сперматозоида собственно в цитоплазму яйцеклетки, при этом, как только сперматозоид проникает в цитоплазму яйцеклетки, тут же начинается кортикальная реакция - резкое уплотнение слоя цитоплазмы около оболочки яйцеклетки, и это не дает проникать другим сперматозоидам.
Различают оплодотворение наружное (у амфибий) и внутреннее (птицы, млекопитающие, человек). А также полиспермное, когда проникает много сперматозоидов (птицы) и моноспермное (млекопит и чел.)
Следовательно, у человека внутреннее моноспермное оплодотворение. Обычно у человека яйцеклетка окружается многими сперматозоидами, и они заставляют ее вращаться, при этом происходит капацитация - активация спермиев под влиянием клеток яйцевода и акросомные реакции спермиев, выброс ферм ентов трипсина и галуранидазы, эти ферменты расщепляют оболочку яйцеклетки. В яйцеклетке образуется два ядра - женское и мужское, их называют пронуклеусами, а такое образование называю синкарион (из 2 разных ядер). Далее происходит слияние ядер, после чего яйцеклетку называют зиготой, это новый одноклеточный организм с диплоидным набором хромосом.
Дробление яйцеклетки у разных организмов может идти по-разному. У человека, как правило, дробление идет от полюса до полюса, затем каждая половинка опять делится, образуется как бы 4 клетки, а след деление идет по экватору, так делятся изолецитальные яйца, которые имеют мало желтка. Отдельные
3
клетки называются бластомерами. У амфибий зигота дробится полностью, но бластомеры не одинаковы, яйцеклетка лежит на желтке у амфибий, и на верхнем полюсе (где нет желтка) - мелкие бластомеры, а на вегетативном нижнем - крупные бластомеры. В результате дробления на стадии 100 клеток образующийся эмбрион называется морула. По аналогии с тутовой ягодой. В телолецитальных яйцах дробление может быть полным, но на вегетативном полюсе дробление медленнее, чем на анимальном (у рыб), или неполное (у птиц), на анимальном вообще нет деления, и зародыш распластовывается по поверхности желтка, образуя диск. При этом бластомеры отличаются внешне, по периферии идут светлые бластомеры, это т. н. трофобласт, через него идет питание зародыша, а в центре темные, это и есть зародыш. Далее трофобласт отслаивается и внутри него образуется полость, которая заполнена жидкостью из матки, а зародыш как бы как диск приклеивается к внутренней поверхности трофобласта. На стадии 128 клеток формируется бластула. За бластулой идет стадия гаструлы (от слова gaster). Здесь наблюдаются сложные перемещения зародышевого материала, образование многослойности. Изолецитальный тип образует второй слой клеток за счет впячивания - инвагинации. Имеется другой способ образования двуслойности - деляминация, расслоение. Наружный слой – эктодерма, внутренний - энтодерма. Существует у птиц эпиболия - окружение (мелкими клетками). У кишечнополостных образование второго слоя идет за счет иммиграции.
Укишечнополостных на этом развитие заканчивается, а у остальных организмов процесс продолжается, образуется 3 зародышевый листок - мезодерма. Мезодерма может образоваться 2 путями: у беспозвоночных - телобластически, а у млекопитающих и человека - энтероцельно, энтодерма образует карманы, далее они отшнуровываются, из них образуется мезодерма, при этом формируется т. н. сомиты.
Например, около нервной трубки формируется дорзальный отдел сомитов. Из эктодермы у человека образуется: 1) нервная трубка; 2) эпидермис кожи, ногти, волосы, потовые и сальные железы, эмаль зубов, воспринимающие клетки глаз, слуха и обоняния.
Из энтодермы образуется эпителий пищеварительных желез, дых. сист., мочеполовой сит, желудка, кишечника, печени.
Из мезодермы образуется мезенхима - соединительная ткань, скопление мезодермы около нервной трубки дает три отдела - 1) склеротом - хрящевая часть кости, 2) миотом - скелетная мускулатура, 3) дерматом
-кожа. Вентральная часть мезодермы, здесь два отдела: 1) нефрогонотом - половая и выделительная система; 2) спланхнотом - мочевыводящие пути, кровеносные сосуды, мускулатура кишечника.
Уэмбриона имеются провизорные органы, которые функционируют только в эмбриональном состоянии. Это желточный мешок и аллантоис, амнион. В стенках желточного мешка образуются кровеносные сосуды и это первый орган кроветворения и снабжения зародыша пищей. У человека желточный мешок врастает в плаценту. Амнион создается для зародыша в виде водной оболочки. Аллантоис или мочевой мешок (пузырь) - обычно это вырост заднего отдела кишечной трубки, через него зародыш осуществляет дыхание, здесь находится скопление мочевины. У человека аллантоис слабо развит, обычно входит в пупочный канатик. Функцию наружной зарод оболочки несет хорион или ворсиночная оболочка, которая врастает в стенку матки, место наиб развития хориона называется плацента. Плацента соединена пупочным канатиком с плодом, и здесь иду кровеносные сосуды, которые снабжают плод всем необходимым от матери. Кровь матери и плода в норме не смешивается, однако бывают случаи, когда это смешение происходит, и тогда, если плод отличается иммунологически, в крови матери появляются антитела и происходит отторжение плода.
3) Третий период эмбриогенеза - постэмбриональный/постнатальный или атенатальный.
4
Лекция 7. РЕГУЛЯЦИЯ ГЕНЕТИЧЕСКОЙ АКТИВНОСТИ На протяжении длительного времени ученые задавались вопросом: кто включает и выключает
гены в нашем организме? Почему в одних тканях и органах работают одни гены, в других органах другие гены? например, когда мы что-то съедаем, как узнают клетки желудка, что надо выпускать определенные ферменты? В 1961 г. два французских ученых Франсуа Жакоб и Жак Люсьен Моно при изучении кишечной палочки добавляли в питательную среду разные сахара (глюкозу, сахарозу, лактозу), и каждый раз микробы начинали выпускать ферменты именно против этих сахаров, т. е. они разлагали данные сахара и использовали их для питания. Как только этот сахар исчезал, выпуск данных ферментов прекращался. В конце конов они пришли к выводу, что существует 2 группы генов:
1)Регуляторные гены («гены-господа»);
2)Структурные гены («гены-рабы»).
Именно главные гены решают, какие ферменты запустить. Они назвали это оперонной системой, регулирующей генетическую активность.
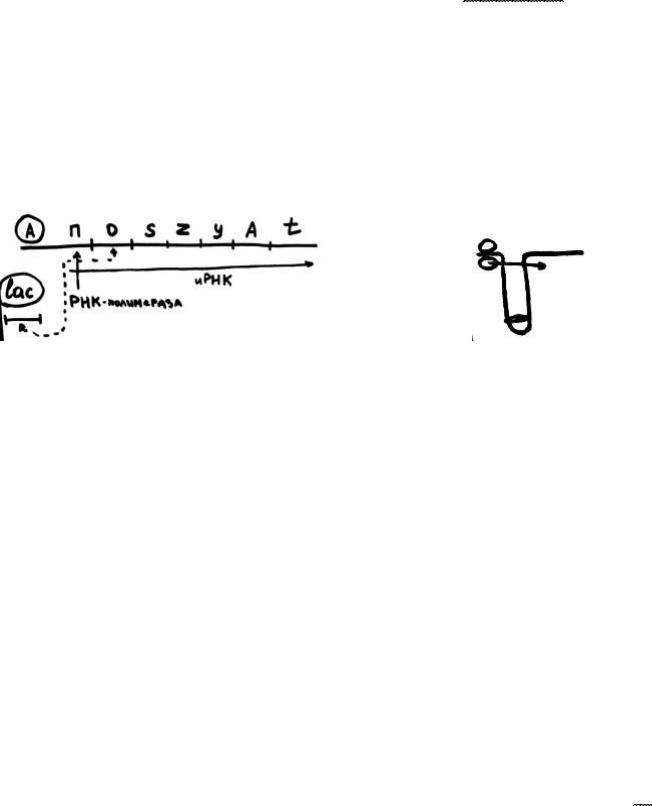
Первый оперон (lac-оперон) состоял:
активатор – промотор – оператор – спейсер – ген z – ген y – ген a –терминатор
Недалеко от него был регуляторный ген. Оператор работает так: синтезируется РНК, которая поступит на рибосомы, далее вырабатывается фермент, микроб переваривает лактозу.
Оказалось, что если в окружающей среде появилась лактоза, и ее надо съесть, к промотору присоединяется РНК-полимераза, и начинается биосинтез, образуется иРНК, образуются ферменты на рибосомах, которые переваривают лактозу. Лактоза исчезла, ферменты не нужны. Тогда с регуляторного гена постоянно образуется маленький регуляторный белок, и когда нет лактозы в окружающей среде, регуляторный белок присоединяется к оператору, теперь РНК-полимераза блокируется, дальше иРНК не образуется, ферменты также не образуются. Если в окружающей среде вновь появляется лактоза, то она имеет сродство к маленькому белку, образует конформацию, белок отпадает, РНК-полимераза вновь может считывать, лактоза вновь переваривается. Оказалось, что микроб разборчив, в смеси он начинает съедать в первую очередь глюкозу, т. к. она более энергетически ценна. Чтобы это не работало, другим оператором вырабатывается ЦАМФ (циклический аденозинмонофосфат). Присоединяется РНКполимераза, и вновь процесс запускается.
Оказалось, что опероны бывают разные: одни катаболические, т. е. предназначенные для переваривания, а другие анаболические, они должны синтезировать что-то новое. Кроме того, различают негативный контроль и позитивный контроль генетической активности, это различие связано с тем, что при негативном контроле (к которому относится лактозный оперон) регуляторный белок блокирует транскрипцию, а при позитивном контроле все наоборот – присоединение регуляторного белка резко усиливает процессы транскрипции, например, к таким оперонам относят арабинозный оперон, т. е. оперон, предназначенный для биосинтеза ферментов, способствующих катаболизму арабинозы (сложный сахар).
Долгое время искали опероны, связанные с анаболизмом, первый такой оперон был связан с биосинтезом аминокислоты триптофана. Описал его американский ученый Чарльз Яновский. Этот оперон кодирует 5 ферментов, данные ферменты превращают простое вещество хоризмата в триптофан в результате пяти биохимических реакций. Здесь стратегия работы оперона несколько иная. Если в окружающей среде уже имеется триптофан, и он не нужен для биосинтеза белка, то нужно срочно остановить биосинтез триптофана, потому что триптофан сам по себе токсичен. Поэтому когда в
1
окружающей среде имеется триптофан, он соединяется с белком регулятором, и этот комплекс присоединяется к оператору и РНК-полимераза не может двигаться вперед. Однако дальнейшие исследования показали, что за оператором имеется т. н., лидерная последовательность и участок, который стали называть аттенюатор, эти участки предназначены для очень тонкой регуляции активности этого оперона. Установлено, что поскольку процессы транскрипции и трансляции происходят одномоментно, то рибосомы, как только образовался кусочек иРНК, садятся на нее и двигаются точно вслед за тем, как движется РНК-полимераза. Если РНК-полимераза останавливается, то эти рибосомы скапливаются около РНК-полимеразы. Оказалось, что в аттенюаторе имеются стоп кодоны, в принципе, РНК-полимераза через них в норме не может пройти, но если происходит скопление рибосом, то происходит изменение конформационной структуры ДНК с образованием шпильки, и рибосома может миновать стоп кодон.
Оказывается, перед стоп кодоном друг за другом стоят кодоны УГГ, а этот кодон требует триптофан в белке, и если в окружающей среде нет триптофана, рибосомы не могут двигаться дальше, образуется «куча мала» из рибосом, стоп кодон уходит в шпильку и РНК-полимераза может двигаться вперед. Она прошла этот путь, на основе нее синтезируется 5 ферментов для синтеза триптофана, он появился в окружающей среде, рибосомы больше не останавливаются, шпилька расправляется, и следующие порции РНК-полимераз уже не могут пройти через это место.
Лактозный оперон (LAC-оперон) – негативный контроль, индуцируемый оперон, а триптофановый оперон – репрессируемый оперон с негативным контролем. Присоединение регуляторного белка в одном из случаев индуцирует транскрипцию, а в другом подавляет транскрипцию.
У эукариот в настоящее время различают несколько видов структурных генов. Во-первых, функционирующие гены разделяются на гены-ингибиторы, подавители, и гены интенсификаторы (которые наоборот усиливают), гены-интеграторы, гены, объединяющие усилия нескольких генов и гены модификаторы, которые способны к модификации продукта. Также выделены регуляторные гены. И гены-операторы, при этим регуляция генетической активности происходит на генном, транскрипционном, трансляционном и функциональном уровнях. Генный уровень связан с количеством
илокализацией генов, имеется явление, называемое амплификацией генов. Количество генов может изменяться, открыты мигрирующие гены и поэтому в зависимости от того, в какое место встроился мигрировавший ген, там и изменится регуляция генетической активности. У эукариот все клетки многоклеточного организма содержат совершенно одинаковую ДНК, однако функционально и даже морфологически они отличаются, это связано с тем, что экспрессия (включение) генов в разных органах
итканях различна. Гены, которые работают во всех клетках одинаково стали называть «гены домашнего хозяйства», а гены, которые работают только в определенных клетках – «гены роскоши». В настоящее время процессы включения и выключения генов у эукариот и влияние внешней среды на этот процесс называют эпигенетика, она изучает явления развития фенотипов организмов при наличие одного и того же генов, т. е наследуемые изменения организма, которые связаны с изменением активности генов.
Впервые американская ученая Барбара Мак-Клинток при изучении наследования у кукурузы окраски зерен обнаружила т. н. «прыгающие» гены. Если раньше ученые полагали, что гены стабильно сцеплены друг с другом, то оказалось, что некоторые гены могут перемещаться по геному, эти гены в литературе называют по разному, чаще бигль (beagle) либо цыганка (gypsy), или транспозоны и инсекции. Барбара Мак-Клинток обнаружила следующие закономерности, два генетических элемента: диссоциатор и активатор, а ген окраски - ген С.
1)Если зерно белое, то следующие закономерности: внутри гена С – диссоциатор, и пигмент не образуется из за этого диссоциатора.
2)Ген С и диссоциатор отдельно, зерно желтое.
3)Появление гена активатора и либо диссоциатор вне (коричневое пятно), либо внутри гена (белое пятно).
Вдальнейшем было установлено, что таким образом образуется
пятнистость у животных, т. е. имеются мигрирующие гены, которые определяют эту окраску у животных.
2
Активатор был в дальнейшем назван транспозоном, а диссоциатор - инсекцией. Изучение последовательностей нуклеотидов в активаторе-диссоциаторе показало, что диссоциатор похож на активатор по последовательности, только он утратил часть генетического материала, а активатор значительно больше, содержит два гена с ферментами резольваза и транспозаза. Резольваза способна вырезать диссоциатор, а транспозаза способна переместить и встроить этот участок в ген С. Активатор состоит из 4500 нуклеотидов. В диссоциаторе всего 194 нуклеотида. Последние исследования генома человека свидетельствуют, что в геноме огромное количество повторений генетических элементов, и они способны перемещаться и регулировать генетическую активность.
ГЕНЕТИКА СОМАТИЧЕСКИХ КЛЕТОК С того момента, когда было установлено, что клетки человека могут жить и размножаться вне
организма начинается отсчет генетики соматических клеток. В 1965 г. английский ученый Харрис впервые получил гибрид клетки человека и мыши, такие клетки называются гетерокарионы. В дальнейшем гибридизация соматических клеток человека сыграла определенную роль в исследовании генетики клеток человека:
1)были изучены механизмы активации и реактивации генов
2)степень фенотипического проявления отдельных генов соматических клеток
3)генетика клеточного деления
4)кротирование генов в хромосомах
5)изучены причины ракового перерождения клеток человека
Гибриды были получены с самыми разнообразными организмами. Известны цибриды, когда ядерный аппарат от одного организма и, например, митохондрии - от другого организма. Возможно создание цибридов, несущих цитоплазматические гены устойчивости к различным инфекциям. Гетерокарионы, могут существовать очень длительное время в культуре, поскольку здесь сливаются ядерные материалы двух организмов (46 хромосом чела и 40 хромосом мыши), то при дальнейшем культивировании начинается элиминация (выброс) хромосом человека, в конце концов получается т. н. синкарион, имеющий 40 хромосом мыши и, допустим, 1 хромосому человека. И возможно сравнить обычную клетку мыши с 40 хромосомами и вот этот синкарион с 1 хромосомой человека. Цитологически можно определить, какая это хромосома и посмотреть, а что лишнего продуцирует эта клетка относит нормальной клетки, какие другие ферменты продуцирует синкарион, все лишнее – продукция генов в данной лишней хромосоме, поэтому этот метод стал использоваться в котировании хромосом человека. Была разработана специальная селекционная система, с помощью которой можно было получать вот такого рода синкарион. Например, известно, что вирус полиомиелита проникает в клетки человека, прикрепляясь за рецепторы на поверхности клетки, а эти рецепторы являются продукцией гена 17 хромосомы кариотипа человека, значит, если в синкарионе есть 17 хромосома, то вирус уничтожит эти клетки. Кроме того, были получены т. н. микроклетки (клетки, содержащие небольшое количество хромосом). Эти микроклетки получали путем центрифугирования слившихся клеток при очень высоких оборотах, при этом в среду добавляли цитохолазин В, он нарушал клеточную мембрану, и клетки разваливались на маленькие фрагменты, в каждом из которых была цитоплазма и ядерный материал, затем эти микроклетки сливали с обычными клетками. Цибриды получают путем получения протопластов также при больших оборотах центрифугирования, ядро выбрасывается из цитоплазмы, и протопласт остается без ядра и его сливают с обычной клеткой.
3

Лекция 8 ОСНОВЫ КЛАССИЧЕСКОЙ ГЕНЕТИКИ – МЕНДЕЛИЗМ.
Впервые в 1865 г. Грегор Мендель, чешский ученый, выступает в обществе естествоиспытателей г. Брюнна (Чехия) со своим докладом, посвященным механизмам и закономерностям наследования признаков на примере садового гороха. Он впервые применил гибридологический метод, т. е. он отбирал константные формы, четко передаваемые по наследству, и анализировал наследование этих признаков у потомков. При этом он анализировал потомство каждого растения в отдельности и проводил точный количественный учет наследования данного признака в ряду поколений.
Первые его эксперименты были связаны с наследованием окраски гороха (зеленые и желтые горошки). Это было моногибридное скрещивание, когда родительские формы отличаются друг от друга только по одной паре альтернативных признаков. При скрещивании растений с желтыми и зелеными семенами у гибридов всегда были только желтые семена. При этом Мендель выводит первый закон, закон единообразия гибридов 1 поколения, который гласит: при скрещивании двух особей одного вида, отличающихся по одному признаку, гибриды первого поколения все будут единообразными, одинаковыми. Во втором поколении при самоопылении произошло расщепление подавленного зеленого признака, этот признак проявился у ¼ части потомков, и расщепление произошло в пропорции 3:1. Вывод: рецессивный признак у гибридов 1 поколения не исчезает, он был подавлен и вновь проявился у гибридов 2 поколения. На этом основании он сформулировал 2 закон: при скрещивании гибридов 1 поколения между собой во 2 поколении наблюдается расщепление признаков в соотношении 3:1. Выявленные Менделем закономерности не были поняты современниками. Только в 1900 трое ученых: Гуго де Фриз, Карл Корренс и Эдвард Чермак обнаружили на разных растениях те же закономерности. Это положило начало такому направлению как менделизм.
Одновременно была выведена гипотеза чистоты гамет, согласно ей в каждую из гамет при преобразовании попадает только один из двух элементов наследственности (аллельные гены), отвечающий за данный признак. Зачем нужна данная гипотеза? До Менделя наследственность представляли как жидкость, и считалось, что жидкости мужчин и женщин сливаются, и возникает нечто среднее, поэтому были такие понятия как «чистокровка», «полукровка» и др. (у лошадей). Мендель впервые четко доказал, что никакого смешения не происходит.
Основные понятия классической генетики.
Признак или фен – некоторое качество или свойство, по которому можно отличить один организм от другого;
Доминирование – преобладание признака какого-либо родителя у гибрида;
Рецессивный признак – не проявляющийся в первом поколении;
Доминантная аллель определяет доминантный признак (А, а˖);
Рецессивная аллель определяет рецессивный признак (а, а¯);
Генотип – совокупность всех генов организма, но мы генотипом часто называем только те гены, которые рассматриваем;
Фенотип – совокупность признаков организма, зависящая от генотипа и особенностей внешней среды. Часто рассматриваемые некоторые внешние признаки называем фенотипом.
Виды скрещивания
1)Моногибридное
2)Дигибридное
3)Полигибридное
|
Генетическая запись скрещивания. |
|
|
|
|
|
|
Дано: |
|
Р: |
♀АА х |
|
♂аа |
|
|
||||||||||||
|
|
|
|
|
|
|
|
|
|
|
А – желтые семена |
|
|
|
|
|
|
|
|
|
|||||||
|
|
|
|
|
|
|
|
|
G: |
А, а |
|
|
|
|
|
||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
(гаметы) |
|
|
|
|
||
|
Правило расщепления гибридов первого поколения |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||
|
|
|
|
|
|
|
а – зеленые семена |
|
|
|
|
|
|
||||||||||||||
|
Аа |
х |
Аа |
|
|
|
|
|
|
|
F1: |
Аа |
|
|
|
|
|
|
|||||||||
|
|
|
|
|
|
|
Р: ♀АА, |
♂аа |
|
|
|
|
|
|
|
|
|
||||||||||
|
(гибриды 1го поколения) |
|
|
|
|
|
|
|
|
|
(желтые семена) |
|
|||||||||||||||
|
Результаты: из 8023 семян гороха 6022 желтого цвета, а 2001 зеленого. |
|
|
|
|
Найти F1 - ? |
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Дано: |
|
|
|
|
Р: |
♀Аа |
х |
♂Аа |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
А – желтые семена |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
G: |
А, а |
а, А |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
а – зеленые семена |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||
|
|
F: |
АА, Аа, Аа, аа |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||
|
Р: ♀Аа, |
♂Аа |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||
|
|
|
|
ж : ж : ж : з |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||
|
Найти F1 - ? |
|
|
|
3:1 по фенотипу; 1:2:1 по генотипу |
|
|
|
|
|
|
|
|
|
|
||||||||||||
|
|
Решетка Пеннета – графический метод, предложенный английским генетиком Р. Пеннетом для наглядного |
|||||||||||||||||||||||||
|
представления о сочетании различных типов половых |
клеток (гамет) при скрещивании. Типы женских |
гамет |
||||||||||||||||||||||||

располагаются вдоль одной, а мужских гамет – вдоль смежной стороны квадрата. Внутри решетки вписывают получившиеся при сочетании гамет генотипы. Метод облегчает расчет возможных сочетаний разных типов гамет.
|
|
|
♀ |
А |
а |
|
|
|
|
|
|
|
|
|
|
|
|
|
♂ |
|
|
|
|
|
А |
|
|
АА - ж |
Аа - ж |
|
а |
|
|
Аа - ж |
аа - з |
|
|
|
|
|
|
Наследственность – способность организма передавать свои признаки.
Изменчивость – свойство приобретать новые признаки.
Генотип – совокупность наследственных типов (генов).
Фенотип – совокупность проявившихся внешних признаков.
Доминантный – признак, который проявляется в первом поколении.
Рецессивный – непроявившийся признак (подавленный).
Гетерозготный – организм, несущий разные аллельные признаки.
Гомозиготный – организм, несущий одинаковые аллельные признаки.
Bb – генотип особи – гетерозиготный организм. Одна пара альтернативных признаков. Два типа гамет.
BB – генотип особи – гомозиготный организм по доминантному признаку. Оба аллельных гена одинаковы. Один тип гамет. Нет альтернативных признаков.
bb – генотип особи – гомозиготный организм по рецессивному признаку. Оба аллельных гена одинаковы. Один тип гамет. Нет альтернативных признаков.
Основные генетические символы. х – гибридизация или скрещивание Р – родительские формы
♀ - женский организм
♂ - мужской организм
F1, F2 и т. д. – потомки или гибриды
Анализирующее скрещивание используется для определения генотипа особи (гомозигота или гетерозигота), для этого ее скрещивают с рецессивной гомозиготой (аавв). Если в первом поколении наблюдается расщепление 1:1:1:1 или 25%:25:25%:25%, значит изучаемая особь дигетерозиготна. Если расщепления не происходит, то изучаемая особь гомозиготна.
Возвратное скрещивание – скрещивание одного из родителей с его потомком.
ВЗАИМОДЕЙСТВИЕ ГЕНОВ.
Взаимодействие генов
|
|
|
|
Взаимодействие аллельных генов |
|
|
|
|
Взаимодействие неаллельных генов |
|
|||||||||||||
|
|
полное |
|
|
|
кодоминирование |
|
|
|
неполное |
|
|
|
|
эпистаз |
|
|
|
полимерия |
|
|
||
|
доминирование |
|
|
|
|
|
доминирование |
|
|
|
|
|
|
комплементарность |
|
||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Кодоминирование (наследование группы крови у человека, гены группы А и гены группы В являются кодоминантными). Когда встречаются два доминантных гена и ни один из них не подавляет действие другого, возникает новый признак, такое взаимодействие обозначается термином «кодоминирование». Встречается в иммунологии и биохимии
в формировании признака у гетерозиготного организма участвуют обе аллели;
расщепление признаков по фенотипу в F2 – 1:2:1
|
Р: |
IᴬIᵒ (2 группа) |
x |
IᴮIᵒ (3 группа) |
|
||
|
G: |
Iᴬ, Iᵒ |
Iᴮ, Iᵒ |
|
|
|
|
F:IᵒIᵒ (1 группа), IᴬIᵒ (2 группа), IᴮIᵒ (3 группа), IᴬIᴮ (4 группа)
Примером кодоминирования служит IV группа крови человека в системе АВ0: генотип - IᴬIᴮ, феноти – АВ, т. е. у людей с IV группой крови в эритроцитах синтезирутся и антиген А (по программе гена Iᴬ) и антиген В (по программе Iᴮ)

Взаимодействие аллельных генов связывают с именем ученого Карла Эриха Корренса. Ярким примером неполного доминирования может служить окраска цветков у горошка, примулы и т. д.
Р: |
♀АА (красные цветки) х |
♂аа (белые цветки) |
|
G: |
А, а |
||
F: |
Аа (розовые цветки) |
||
В дальнейшем при скрещивании F1 происходит расщепление по фенотипу и по генотипу в соотношении 1 (красн.): 2 (роз.): 1 (бел.)
Морган впервые 1714 установил, что наследование некоторые признаков не подчиняется законам Менделя. Он впервые доказал, что это связано со сцеплением генов. Например, 1 хромосома у дрозофилы.
коричневое тело |
– |
желтое тело |
волосатое тело |
– |
гладкое тело |
толстое тело |
– |
тонкое тело |
слияние жилок |
– |
сплетение жилок |
красные глаза |
– |
розовые глаза |
зачаточные крылья |
– |
длинные крылья |
центромера
Другой пример: ген глухоты и ген голубых глаз сцеплены у кошек практически всегда.
Постепенно накапливались факты, показывающие, что взаимоотношения между генами и признаками носят более сложный характер. Выяснилось, что один и тот же ген может оказывать влияние на развитие нескольких признаков. Один и тот же признак может развиваться под влиянием многих генов.
Известно несколько типов взаимодействия неаллельных генов:
1) комплементарность (комплементарное взаимодействие). Комплементарными называют гены, обусловливающие при совместном сочетании в генотипе в гомозиготном или гетерозиготном состоянии новое фенотипическое проявление признака.
Классическим примером комплементарного взаимодействия генов является наследование формы гребня у кур. При скрещивании кур, имеющих розовидный и гороховидный гребни, все первое поколение имеет ореховидный гребень. При скрещивании гибридов первого поколения, у потомков наблюдается расщепление, по форме гребня на 9 ореховидных приходится 3 розовидных, 3 гороховидных и 1 листовидный гребень
Р: ♀ааBB (гороховидный) х ♂AAbb (розовидный)
F1: AaBb (ореховидный)
F2: А_В_ (9/16 – ореховидный), ааВ_ (3/16 - розовидный), A_bb (3/16 – гороховидный), aabb (1/16 – листовидный)
Выводы:
1)Если первое поколение единообразно, значит родители – гомозиготы.
2)Если во втором поколении расщепление в соотношении 9:3:3:1, значит первое поколение – двойные гетерозиготы, и гены расположены в разных парах гомологичных хромосом и имеют генотип AaBb.
3)9/16 с ореховидным гребнем имеют геноти A_B_, 3/16 с гороховидным и 3/16 с розовидным могут иметь генотипы A_bb и aaB_, особи с листовидным гребнем имеют генотип aabb.
4)Родители имеют генотипы aaBB и AAbb.
Упопугаев цвет перьев определяется двумя парами генов. Сочетание двух доминантных генов определяет зеленый цвет. Рецессивные по обеим парам генов особи имеют белый цвет. Сочетание доминантного гена А и рецессивного гена b определяет желтый цвет, а сочетание рецессивного гена а с доминантным геном В – голубой цвет.
Задача. При скрещивании между собой двух зеленых особей получили попугаев всех цветов. Определите генотипы всех потомков.
|
|
|
♀ |
AB |
aB |
Ab |
ab |
|
|
|
|
|
|
|
|
|
♂ |
|
|
|
|
|
|
|
AB |
|
AABB (зеленый) |
AaBB (зеленый) |
AABb (зеленый) |
AaBb (зеленый) |
|
|
aB |
|
AaBB (зеленый) |
aaBB (голубой) |
AaBb (зеленый) |
aaBb (голубой) |
|
|
Ab |
|
AABb (зеленый) |
AaBb (зеленый) |
AAbb (желтый) |
Aabb (желтый) |
|
|
ab |
|
AaBb (зеленый) |
aaBb (голубой) |
Aabb (желтый) |
aabb (белый) |
|

Эпистаз. Эпистатичным называют такое взаимодействие генов, при котором аллель одного гена подавляет действие аллелей других генов. Ген-подавитель называется эпистатичным, а подавляемый – гипостатичным. При доминантном эпистазе доминантная аллель гена является эпистатичной, а при рецессивном – рецессивная.
Задача. При скрещивании кур двух пород белого цвета – леггорнов с плимутроками все потомство F1 имеет белую окраску. Если же гибридов F1 скрестить между собой, то во втором поколении F2 происходит расщепление по окраске в соотношении 13/16 белых и 3/16 окрашенных. Запишите генетическую схему скрещиваний и объясните полученное расщепление.
Р: |
♀ССII (белая) х |
♂ссii (белый) |
|
F1: СсIi (белые) |
|||
F2: |
С_I_ ccI_ ccii (13/16 - белые); С_ii (3/16окрашенные) |
||
Выводы из анализа родителей и потомства:
Родители гомозиготны, т. к. F1 единообразно. Если во втором поколении расщепление на 16 вариантов, то F1 – двойные гетерозиготы и гены расположены в разных парах хромосом.
Генотипы F1: CcIi
Окрашенными могут быть только особи 3/16 с генотипами C_ii или 3/16 с генотипами ccI_. 9/16 с генотипом C_I_ белые (из-за гена-подавителя).
1/16 ccii белые.
F1 белые, т. к. имеют генотип CcIi. Генотипы родителей: CCII и ccii
Задача. Ген W обуславливает белый цвет плодов тыквы, причем при его наличии ген Y не проявляется. При генотипе wwYY или wwYy плоды имеют желтый цвет. Наконец, если оба гена рецессивны, то плоды зеленые. Каково будет расщепление по фенотипу в F2 при скрещивании тыкв с генотипами WWYY (белые) и wwyy (зеленые)?
Р: |
♀WWYY (белая) |
х |
♂wwyy(зеленая) |
|
F1: WwYy (белая) |
|
|
|
|
F2: |
W_Y_ (9/16 – белые) |
W_yy (3/16 – белые) wwY_ (3/16 – желтые) wwyy (1/16 – зеленые) |
||
|
Расщепление в соотношении 9:3:3:1 |
|||
Задача. От скрещивания тыквы с белыми плодами с тыквой, имеющей зеленые плоды с генотипом wwyy, в F2 ½ плодов имеют белую окраску, ¼ желтую и ¼ зеленую. Каковы генотипы гибридов и родителей? Объясните полученное расщепление.
Р: |
♀ WwYy (белая) х |
♂wwyy(зеленая) |
|
F1: |
W_Y_ (9/16 – белые) |
W_yy (3/16 – белые) wwY_ (3/16 – желтые) wwyy (1/16 – зеленые) |
|
Вывод: это анализирующее скрещивание и второй из родителей двойная гетерозигота с генотипом WwYy и образует 4 типа гамет, т. к. в F1 4 варианта генотипов.
Особи с генотипами WwYy и Wwyy имеют белые плоды. Особи с генотипом wwyy – зеленые, особи с генотипами wwYy – желтые.
Следовательно, ген W – подавитель, ген Y отвечает за желтую окраску, а ген y – за зеленую.
Задача. Окрашенность шерсти кроликов (в противоположность альбинизму) определяется доминантным геном Взаим неал генов. У человека вместе с альбинизм появл ослабленный иммунитет, светобоязнь. Один ген может несколько призн
От количества обр зерен меланина зависит окраска кожи. Меланоциты защищают отУФ лучей. В африке у многих белокожий высокий риск заболевания раком кожи. Под влиянием УФ лучей обр витамин Д. Постепенно светлый цвет кожи на севере стал преобладать. Произошел отбор в сторону белокожих. Поэтому на севере имеют преимущественно белый цвет кожи. У темнокожих высокий риск заболеванием рахитом, особенно у детей.
Чем больше генов, тем интенсивнее окраска. Аддитивная полимерия. Накопит. Неаддит, ненакопительная
Серповидно кл анемия. Гемоглобин склеивается в столбики, эритроциты не могут нормально переносить кислород. Заменена всего одна аминокислота и поэтому молекулы электростатически притягиваются. В норме же распределены диффузно.

Лекция 9. СОСТАВЛЕНИЕ ГЕНЕТИЧЕСКИХ КАРТ ХРОМОСОМ ЧЕЛОВЕКА.
Впервые в 1911 г. американский ученый Томас Морган выдвинул хромосомную теорию наследственности. Он в качестве объекта исследования использовал муху-дрозофилу. Это оказался отличный генетический объект. Репродуктивный период этого организма составляет 10-12 дней, т. е. из отложенного яйца вылупляется муха и уже через 10 дней дает следующее потомство. Одна самка может дать около 100 потомков. Живет на минимальной питательной среде, питаясь, в основном, дрожжами. Морган при изучении наследования признаков у дрозофилы впервые обнаружил расщепление, не соответствующее 3 закону Менделя (закону независимого комбинирования признаков у потомков). Как оказалось, часть генов наследовалась вместе, сцеплено, передаваясь из поколения в поколение. Бельгийский ученый Янсенс в это время изучал хромосомы дрозофилы, Морган установил, что у мухи 4 группы сцепления, а Янсенс - что у нее 4 пары хромосом, так была доказана хромосомная теория наследственности.
Основные законы:
1.гены, находящиеся в 1 хромосоме образуют одну группу сцепления;
2.гены располагаются в хромосоме линейно друг за другом;
3.поскольку гомологичные хромосомы в мейозе конъюгируют и образуют кроссинговер, то сцепленные гены могут наследоваться врозь и чем чаще происходит между генами кроссинговер, тем чаще происходит процесс расцепления генов, и чем ближе, тем частота кроссинговера будет
ниже.
Число групп сцепления генов равно гаплоидному набору хромосом, т. е. у человека 23, у мыши 20, дрозофилы 4 и т. д. Морган доказал, что гены находятся в хромосоме в строго определенном участке.
Огромное значение для картирования генов сыграло предположение о том, что расстояние между 2 генами можно охарактеризовать частотой рекомбинаций, кроссинговеров. Цитологический обмен между хромосомами происходит в хиазмах, переплетениях. Однако, как оказалось, не каждая хиазма приводит к кроссинговеру. Число хиазм, ведущих к рекомбинации в два раза больше числа рекомбинантов, т. е. к кроссинговеру приводит только каждая вторая хиазма.
Единицей хромосомной карты (Θ, тэта) служит расстояние, на котором в среднем происходит 1 кроссинговер на 100 гамет, это стали называть сантиморганиды (сМ), т. е. 1% рекомбинации – это одна сантиморганида. Если, например, два гена удалены друг от друга на 20 единиц хромосомной карты (20 сМ), то 20 гамет из 100 обнаруживают кроссинговер, а на цитологическом уровне можно увидеть 40 хиазм между этими участками. Исследуя частоту хиазм можно определить генетическую длину хромосом. Например, длина х-хромосомы от 160 до 250 сМ, общая длина всех аутосом 2650 – 3333 сМ. Такие колебания связаны с тем, что частота кроссинговеров у женщин в 2 раза выше, чем у мужчин. Этот феномен распространяется практически на весь животный мир, причина непонятна. У самцов дрозофилы вообще нет кроссинговера.
Моргановское картирование хромосом. Представим женский организм, у которого все доминантные гены в одной хромосоме, а рецессивные – в другой. Скрещиваем с самцом – рецессивной гомозиготой.
|
P: |
♀ ABC |
|
|
x |
|
|
|
|
|
♂ abc |
|
|
|
|
abc |
|
|
|
|
|
|
|
|
abc |
|
|
|
G: |
ABC |
abc |
I: Abc |
aBC |
II: |
ABc |
дв.: aBc AbC |
|
abc |
|
||
|
|
|
|
|
|
abC |
|
|
|
|
|
|
|
|
F: |
ABC |
abc |
Abc |
aBC |
ABc |
abC |
aBc |
AbC |
|
|
|
|
|
|
abc |
abc |
abc |
abc |
abc |
abc |
abc |
abc |
|
|
|
|
|
|
(88%) |
|
(6%) |
|
(4%) |
|
(2%) |
|
|
|
|
|
I AB (расстояние между участками): 6+2=8сМ
II BC: 4+2=6 сМ
На этом принципе Морган создал генетические карты дрозофилы. Большая часть генов дрозофилы была создана на основе этого метода. Естественно, ученые попытались создать нечто похожее для человека. Но здесь оказалось много трудностей:
1

1)нельзя в нужном направлении провести скрещивание;
2)очень длительный репродуктивный период;
3)нужно знать, в каком положении находятся гены человека (цисили транс-);
4)большое количество хромосом.
AB |
Ab |
ab (цис-положение) |
aB (транс-положение) |
Если в цис-положении АВ и аb не кроссоверные, то в транс-положении все наоборот. |
|
Наиболее хорошо |
на первом этапе была изучена х-хромосома человека. Впервые |
американский ученый МакКьюсик предложил особый метод по составлению генетических карт х- хромосом. Во-первых, он попытался составить генетическую карту расстояния для дальтонизма и гемофилии. Эти два гена давно известны. Если женщина имеет расположение в цис-фазе, а у мужчины есть обе патологии, тогда в потомстве один сын нормальный, некроссоверный, у второго есть обе аномалии, следующий сын кроссоверный с нормальным зрением и гемофилией, еще один следующий кроссоверный дальтоник без гемофилии.
|
P: |
♀DG x |
|
♂dg |
|
|
|
|
|
|
|
dg |
|
|
|
|
|
|
|
|
F: |
DG (обе аномалии) |
dg (здоровый) |
<некроссоверные> ; Dg (дальтоник) |
dG (болен |
||||
|
гемофилией) |
<кроссоверные> |
|
|
|||||
Мы не знаем, в каком положении находятся гены у матери, если мать будет иметь гены в трансположении, то все будет наоборот, одни кроссоверные, другие нет. Это метод МакКьюсика получил название метода ДЕДА. Метод заключается в том, что надо смотреть на отца этой матери, если отец будет dG или Dg, то дочь в цис-положении, а если отец будет DG или dg, то дочь в транс-положении. В связи с этим можно точно говорить, какие потомки кроссоверные, а какие нет. Всего он посмотрел около 107 семей, где встречаются обе аномалии и выявил следующие закономерности, что из 100 семей - с кроссинговером 5 человек, следовательно, расстояние между генами гемофилии и дальтонизма 5 сМ. Он картировал х-хромосому у человека, определяя расстояние между генами дальтонизма и гемофилии, но для этого требовалось установить, как эти гены расположены: в цис- (в одной и той же хромосоме) или в транс- (в разных хромосомах). После чего было определено количество кроссоверных потомков у матерей и установлено расстояние между генами дальтонизма и гемофилии.
Дальнейший этап картирования хромосом связан с изучениями хромосомных перестроек в кариотипе человека, особенно перестроек, связанных с удвоением числа хромосом, дубликацией участков хромосом, утратой хромосомы и делецией (утратой участка хромосомы). В таких клетках у такого человека определяют, что исчезает и что добавляется? какие ферменты исчезают? И видя в хромосоме утрату, устанавливают, что в этом участке находится ген, который определяет синтез того или иного фермента. Особенно много исследований в этом направлении появилось после того, как возникла дифференциальная окраска хромосом, и стало возможным сегментирование участков хромосом.
Следующий этап – это появление метода гибридизации соматических клеток, когда происходит слияние клеток человека и мыши либо хомяка, при этом образуется гибридная клетка синкарион, в которой происходит утрата человеческих хромосом, постепенно возникают клетки, содержащие все хромосомы мыши и 1 хромосому человека. И опять же определяют, что лишнее продуцирует эта клетка относительно нормальной мышиной клетки. Такое слияние проводят с помощью вируса Сендай или полиэтиленгликоля. Затруднение заключается в том, что мы можем установить, какие ферменты, вернее гены каких ферментов находятся в данной хромосоме. Обычно это состояние называют синтенность, приуроченность к какой либо хромосоме.
Установить расстояние между генами предложили американские ученые Госс и Харльз. Они назвали свой метод методом дробления. Такие гибридные клетки, содержащие 1 хромосому человека,
2
облучали мощным рентгеновским пучком, и хромосома начинала распадаться на куски. И далее смотрели, какие ферменты утрачиваются в клетке с делетированной хромосомой, когда видят, что утрачиваются эти 2 фермента, говорят, что эти ферменты синтенны, они сцеплены друг с другом. Т. о. были проанализированы практически все хромосомы человека, была установлена локализация более 1,5 тыс. генов.
С 1990 года началась новая эпоха картирования хромосом человека, был запущен проект «Геном человека». Был использован совершенно новый метод. В основе метода лежит вариант метода Сэнгера или метод секвенирования ДНК, т. е. метод определения последовательности нуклеотидов в молекуле ДНК. Если взять от 46 хромосом у человека молекулы ДНК, то длина будет около 2 метров, было установлено, что в этой ДНК 3,2 млрд пар нуклеотидов и нужно было по одному нуклеотиду определить всю последовательность, как они сцеплены друг с другом.
В методе Сенгера одна из цепочек ДНК выступает в качестве матрицы, на ней с помощью ДНКполимеразы синтезируется комплементарная цепь, т. е. сперва нужно раскрутить молекулы ДНК, обычно это делают при повышении температуры. Цепи ДНК отходят друг от друга. Реакцию обычно проводят в 4 пробирках. Здесь нужен праймер – небольшой участок ДНК, который комплементарен началу молекулы ДНК. ДНК-полимераза не может начинать считывание с пустого места, она только присоединяет нуклеотиды к 3'-гидроксильной группе, это затравка. Необходим дезоксинуклеотид, меченый радиоактивно, часто метят Р32, это позволяет в дальнейшем определить, где произошел синтез. Кроме того, в эту смесь вводят 4 дезоксинуклеотида, чтобы протекал синтез. На сегодняшний день синтезирование по Сенгеру полностью автоматизировано, используются приборы секвенаторами, которые могут за 1 раз прочитать последовательность около 1000 нуклеотидов. Т. о. в настоящее время как бы существует три типа генетических карт хромосом человека:
1)генетические карты, созданные с помощью метода Моргана, измерение в сМ, но оказалось, что в разных местах хромосомы (в гетерохроматине или эухроматине) кроссинговер идет по-разному, поэтому это измерение чисто физиологическое, а не физическое;
2)физические карты хромосом создаются на основе изучения под микроскопом дифференциально окрашенных хромосом, когда определенной зоне хромосом присваивается номер и устанавливается, какие гены находятся в этом участке;
3)секвенцерные хромосомы, когда участок ДНК разбивается на отдельные нуклеотиды, измерение при этом идет в т.н. Мb (мегабазах), 1 Мb – это примерно 1 млн нуклеотидов. Метод секвенциса позволяет проводить определение участков до 1 нуклеотида. Однако ученые не могут эту
последовательность приурочить к определенному гену, известно только 3% В настоящий момент у человека насчитывают около 30000 генов. В настоящее время использование
новых устройств для синтезирования позволяет расшифровывать со скоростью около 500 Mb в год и стоимость этой работы 0,25 $ за нуклеотид. Результаты завершающей части проекта показали, что большая часть генов ацитилированных связаны с устройством и работой головного мозга, поэтому очевидно, что практически любое нарушение в генетическом аппарате приводит к резкому снижению интеллекта. Из 10 тысяч известных болезней половина – это наследственные болезни, они вызваны изменениями в генетическом аппарате, поэтому одно из направлений – это выявить, что сломано в генетическом аппарате и, может быть, появятся методы, с помощью которых можно будет помочь больному.
Кроме этого, данные по бактериям, животным и прочие помогли построить эволюционную лестницу существ. До последнего времени к прокариотам относили еще более примитивные археобактерии которые резко отличаются от других микробов, однако стало ясно, что это совершенно другая ветвь эволюции. Кроме того, данный метод позволил по-новому взглянуть на эволюцию человека. Стали создавать генетические паспорта людей (особенно в США), где обязательно прописаны те гены, изменение которых может приводить к заболеваниям.
3
Лекция 10. ГЕНЕТИКА ОПРЕДЕЛЕНИЯ ПОЛА Уже на уровне бактерий имеются механизмы, которые можно назвать генетическое определение
пола у бактерий. Различают бактерии F+, имеющие особый половой фактор, и бактерии F-, которые этого полового фактора не содержат. F+-плазмида может разрывать кольцевую хромосому у бактерий, встраиваться в нее, и после этого бактерия подходит к F--бактерии и между ними образуется эндоплазматический мостик, после чего плазмида-F начинает толкать через этот плазматичекий мостик хромосому в F—бактерию. Когда этот процесс завершается, между двумя хромосомами происходит рекомбинация генетического материала, после чего такая рекомбинированная хромосома прикрепляется к клеточной стенке, и начинается процесс репликации, который американские ученые называют «по типу рулона туалетной бумаги». Заключается этот процесс в том, что точка репликации начинается от места разрыва, где прикрепилась хромосома, и по кругу синтезируется кольцевая хромосома одна, вторая и т. д. Около отделившейся хромосомы обосабливается своя цитоплазма, формируется клетка и начинается процесс деления, размножения бактерий.
У многоклеточных организмов различают 4 типа определения пола:
1.эпигамное определение пола,
2.прогамное,
3.сингамное,
4.иногда выделяют эусингамное определение пола.
Итак, эпигамное определение пола - когда пол определяется условиями окружающей среды, например, морской червь Bonellia, у которого самец имеет микроскопические размеры и как симбионт обитает в половых протоках самки, самка же откладывает оплодотворенные яйца, их них развиваются личинки. Если личинка обнаруживает виргинную (девственную) самку, проникает в ее половые протоки, то превращается в самца, если этого не происходит, то прикрепляется ко дну и развивается в самку. У крокодилов, черепах, некоторых ящериц из отложенных яиц появляются самки или самцы в зависимости от того, какая температура окружающей среды. Часто если она понижена, то развиваются самцы, если повышена, то развиваются самки. Причем такие различия могут быть даже внутри гнезда, те особи, которые оказались ниже (а там более прохладно) – самцы; а те, что выше – самки. Или у черепахи – ближе к берегу самцы (более холодная среда). Некоторые ученые полагают, что это причина вымирания динозавров, когда появились в определенный период только самки или только самцы. Другой пример – это улитка-блюдечко (Crepidula fornicata), особи поселяются друг на друге, вначале это самцы, которые постепенно, опускаясь ниже, превращаются в самок. Факторы внешней среды, определяющие формирование пола у позвоночных (пресмыкающихся) могут действовать через изменение баланса половых гормонов. Мужские половые гормоны – андрогены (тестостерон и ряд других), женские половые гормоны – эстрогены. Показано, что фермент Р450 ароматаза, катализирующий превращение тестостерона в эстроген, у пресмыкающихся способен менять свою активность в зависимости от температуры окружающей среды. Температура инкубации яиц в последнюю треть развития пресмыкающихся (черепах, крокодилов, аллигаторов) является определяющей для формирования пола выводящихся из яиц животных.
Прогамное определение пола. Пол определяется еще на уровне яйцеклеток. Например, у тлей и коловраток, когда из крупных яиц появляются самки, а из мелких самцы.
Сингамное определение пола. Пол определяется в момент оплодотворения, зачатия.
Различают генное и хромосомное определение пола. Генное, - например, у растения брионии (относится к тыквинным), у них Аа – это мужское растение, аа – женское, при скрещивании в потомстве 50% Аа и 50% аа.
Хромосомное определение пола. Впервые половые хромосомы были обнаружены еще в 20х гг прошлого века у водяных клопов рода Protenor, оказалось, что у их самок 12 хромосом, а у самцов 11. У самца была одна необычная хромосома, ее назвали X-хромосома. Пол в этом случае у самок XX, а у самцов X0, т.е. в сперматогенезе у самца 1 сперматозоид содержит X-хромосому, а второй вообще не содержит половых хромосом. Позднее у водяных клопов рода Lуgaeus обнаружили, что самец содержит тоже 12 хромосом, здесь обнаружили маленькую Y-хромосому.Y-хромосома генетически очень бедна, в ней большая часть представляет из себя гетерохроматин. При мейозе X и Y-хромосомы должны конъюгировать, чтобы потом четко расходиться к разным полюсам. Это очень маленький район и в Y-хромосоме заблокированы гены от кроссинговера, наложен блок, иначе бы гены, которые определяют мужской пол, перекочевали бы в X-хромосому (что очень редко, но всетаки происходит). Определение пола по типу Ligaeus разделяют:
1.по типу дрозофилы
2.по типу человека
Впервые еще в 1922 г. американский ученый Бриджес при изучении дрозофил получил разные мутации, изменения по числу хромосом. У дрозофилы 4 пары хромосом, у самок ХХ, у самцов XY,но как доказал Бриджес, Y-хромосома не определяет мужского пола, здесь нет генов, которые бы определяли мужской пол, она определяет только плодовитость самца. Гены, определяющие пол, находятся в аутосомах, а их три пары. Пол определяется балансом между аутосомами и X- хромосомами. Если число X-хромосом будет не 2 хромосомы, а меньше, например одна, то пол будет мужской, самец. Если аутосом будет больше, чем X-хромосом, то тоже пол будет мужской. Бриджес вывел индекс соотношения X-хромосом и аутосом. У нормальных самок соотношение будет 1:1, у самцов 0,5,если соотношение будет 1,5 и даже больше, то этих мух он назвал сверхсамками, а если в соотношении 0,33 то это суперсамец, и при соотношении 0,67 – промежуточный пол (интерсекс). В отличие от дрозофилы, у человека Y-хромосома содержит особый ген SRY, который определяет развитие пола в мужскую сторону. На первых этапах развития эмбриона у него закладывается два протока – мюллеров и вольфов. Если эмбрион мужской и у него имеется в Y-хромосоме ген SRY, начинает вырабатываться антимюллеров гормон, это приводит к деградации мюллерова протока, он постепенно исчезает. Этот гормон у мужчин функционирует до полового созревания, а из вольфова протока развивается все, что характерно для мужской половой сферы. Если у эмбриона нет этого гена, предположим, что это женский эмбрион XX, то начинает развиваться мюллеров проток, и развитие идет в женскую сторону, вольфов проток при этом деградирует. У человека существуют разнообразные аномалии в числе половых хромосом
Синдром Шершевского-Тернера. Всего 45 хромосом, одна половая X-хромосома, женский фенотип, как правило, это женщины низкого роста, интеллект немного снижен. Полового хроматина нет, при анализе он не проявляется. Детей быть не может, практически нет яичников, стерильны.
Синдром Клайнфельтера. 47,48,49 хромосом XY, XXY, XXXY, мужчины, интеллект ослаблен, проявляют часто во время призыва в армию, как правило, высокого роста, стерильны. Определяется половой хроматин.
Синдром суперженщины, или трисомия по X-хромосоме. 47 хромосом, но может быть 3, 4 и даже 5 X-хромосом. Внешне отклонений нет, однако половина детей рождается с аномалиями из за неправильного расхождения хромосом.
Синдром агрессивности – лишняя Y-хромосома. 47 хромосом, XYY. Отличаются ярко выраженным агрессивным поведением. Могут иметь детей, но половина рождается с аномалиями.
Эусингамное определение пола характерно для некоторых насекомых (пчел, муравьев), самцы имеют гаплоидный набор хромосом, а самки имеют диплоидный набор хромосом, к ним относятся матка и рабочие пчелы. Матка оплодотворяется один раз в жизни, и семя содержится в специальном семяприемнике, часть откладываемых яиц проходит через этот семяприемник, и из них образуются рабочие пчелы или матки, а другая часть минует семяприемник, и из гаплоидных яиц, неоплодотворенных по существу, развивается трутень, т. е. самец. Такое определение пола самца является партеногенетическим. Партеногенетическое определение пола распространено среди некоторых насекомых, например, среди тлей. У них в летний период самки откладывают крупные яйца, и из них без оплодотворения развиваются только самки. Осенью с наступлением холодов самки начинают откладывать крупные и мелкие яйца, которые перезимовывают и весной из крупных появляются самки, а из мелких яиц самцы. Как только условия становятся благоприятными, происходит развитие только самок. Существует такое явление, которое называют гиногенезом, например, у серебристого карася, при гиногенезе самцов не существует, а сперматозоиды других рыб проникают в икринки и стимулируют их к развитию, при этом никакой гибридизации не происходит. Другое явление называется андрогенез, при андрогенезе два сперматозоида проникают в яйцеклетку, превращаются в пронуклеусы и сливаются вместе, при этом ядро яйцеклетки деградирует, вся наследственность становится мужской у этого организма.
Пол – совокупность морфологических, физиологических, биохимических и других признаков организма, обусловливающих воспроизведение себе подобных.
Половые хромосомы. Х-хромосома связана с более чем 300 болезнями (дальтонизм, аутизм, гемофилия, умственное развитие, мускульная дистрофия). Х-хромосомы могут затрагивать особей мужского пола, т. к. они не имеют другой Х-хромосомы, чтобы дать компенсацию за мутации. Гоносомное определение пола. Y-хромосома гораздо меньше, чем Х-хромосома; содержит меньшее количество генов. Известно несколько признаков, гены которых в Y-хромосомах и передаются от отца всем сыновьям, внукам и т. д.
Существует понятие признаков сцепленных с полом, это в случае, когда гены этих признаков находятся в половых хромосомах. Если аномальный ген находится в Х-хромосоме, то обычно страдают сыновья матерей, имеющих аномалию, т. к. мужчины являются гемизиготами, когда признак представлен только одной аллелью, например у мужчин все гены в Y-хромосоме гемизиготны.
Гены, содержащиеся в Y-хромосоме, передаются только сыновьям – голандрическое наследование. Признаки, наследуемые с половыми хромосомами Х и Y получили название сцепленных с полом. Y-хромосому называют генетически инертной, или генетически пустой, так как в ней очень мало генов. На Y-хромосоме у человека находится ряд генов, регулирующих сперматогенез, проявления генов гистосовместимости, влияющих на размер зубов и т. д. Известны аномалии, связанные с Y-хромосомой, которые от отца передаются всем сыновьям (чешуйчатость кожи, перепончатые пальцы, сильное оволосение на ушах – трихопатия). Известно более 300 болезней, сцепленных с Х-хромосомой. Поскольку у особей мужского пола 1 Х-хромосома, то все локализованные в ней гены, даже рецессивные сразу проявляются в фенотипе.

Лекция 11. ГЕНЕТИКА ПОПУЛЯЦИЙ Популяция – это совокупность особей одного вида, длительно населяющих определенную территорию
относительно изолированную от других территорий, свободно скрещивающихся друг с другом и имеющих плодовитое потомство. Термин «популяция» впервые был использован в 1903 г. датским генетиком Вильгельмом Иогансеном. Он понимал под термином «популяция» группу особей, неоднородных в генетическом отношении. В переводе с латинского populus – население. Иогансен впервые вводит понятие «чистая линия», он экспериментирует на самоопыляющейся фасоли и устанавливает, что у тех организмов, которые длительно скрещиваются внутри себя, возникает генетически чистая линия. Он проводил отбор на вес семян в этих линиях. Вес семян колебался от 150 до 750 мг, но независимо от того , брал ли он в потомство 150 мг семена или 750 мг, в потомстве все равно идет колебание 150-750. И он доказывает, что отбор в чистых линиях ни к чему не приводит, потому что эти колебания не генетические, а модификационные, отбирать можно только в гетерогенных популяциях.
На Земле насчитывается около полутора миллионов видов животных и около 500 тыс. видов растений. Различают мегаэволюцию (на уровне типов и классов), макроэволюцию (на уровне родов и семейств) и микроэволюцию (на уровне видов и популяций). Новое видообразование происходит на уровне видов и популяций. Вид и популяция обладают двумя противоположными свойствами:
способность к стабильной передаче своих свойств;
способность к изменчивости.
В1859 г., когда Ч. Дарвин опубликовал свою книгу, его противником был известный ученый-математик Дженкенс, и он впервые задал вопрос: представим себе, что вдруг возникло изменение, но это изменение возникло у единственной особи, и она неминуемо будет скрещиваться с обычными особями, следовательно, это изменение в следующем поколении сократится пополам, затем еще и еще, и в конце концов оно должно исчезнуть. Такое понимание связано с тем, что большинство ученых того периода полагали, что наследственность (как жидкость) смешивается. И только Мендель доказал, что это константные единицы.
Впервые в 1908 г. англ. математик Годфри Харди сформулировал понятие «панмиксия» - свободное скрещивание между организмами - и выдал математическую модель для описания генетической структуры панмиктической популяции. В это же время нем. врач-антрополог Вильгельм Вайнберг независимо создает сходную модель панмиктической человеческой популяции. Эти ученые панмиктическую популяцию
представили как уравнение Харди-Вайнберга: ( + )2 = 2 + + 2, где 2 − доминантные гомозиготные аллели АА, 2 − рецессивные аллели аа, pq –гетерозиготы. Они пришли к выводу, что в идеальной популяции несмотря на длительный период жизни ее должно наблюдаться равновесие гамет и особей.
В идеальной популяции частоты ген и генотипов находятся в равновесии и не изм еняются в ряду поколений. Различают следующие человеческие популяции по численности:
большие – не менее 4000 человек;
малые делятся на демы (число людей от 1,5 до 4 тыс, внутригрупповые браки 80-90%, приток генов из других групп 1-2%) и изоляты (до 1,5 тыс. человек, внутригрупповые браки выше 90%, приток генов
менее 1%)
Человеческие популяции характеризуются следующими демографическими показателями:
размерами;
рождаемостью и смертностью, разница между которыми определяет прирост населения;
возрастной структурой
родом занятий
экологией среды
климатическими условиями
экономическим положением
Популяции называются панмиксными, если в них случайные неограниченные скрещивания и случайный выбор партнера. В идеальной популяции - панмиксия, нет мутаций и нет естественного отбора. Очевидно, что в малых популяциях все это проявляется, и только в очень больших популяциях эти факторы оказывают незначительное действие.
Популяции характеризуются 1. Численностью. Существует нижний предел численности, ниже которого популяция не может
существовать.
2. Плотностью. Плотность определяется как среднее число особей на единицу площади (среднее число особей).
Низкая плотность сокращает шансы на воспроизведение, но в некоторых условиях увеличивает шансы на выживание, т. е. должна быть оптимальная плотность.
3.Популяционный ареал. В идеальной популяции можно выделить ядро – это место, где плотность наибольшая, дальше выделяют субпериферию и периферию.
4.Рождаемость. Число новых особей, появляющихся за единицу времени – абсолютная рождаемость. Отношение числа новых особей к числу имеющихся в этой популяции называют удельной или
относительной рождаемостью. Относительная рождаемость рассчитывает или на одну особь или на 1000. В ходе размножения численность популяции изменяется, поэтому введено понятие мгновенной удельной рождаемости, т. е. рождаемость в пересчете на одну особь за бесконечно малый промежуток времени, а малый промежуток зависит от вида. Например, для человека абсолютно малым считается один год. Существуют моноциклические организмы, которые размножаются один раз в жизни, и полициклические, которые размножаются много раз. У раздельнополых диплоидных организмов для размножения требуется пара родителей, поэтому здесь, чтобы упростить, учитываются только женские особи, но с точки зрения генетики мужские и женские особи в равной степени передают свои гены потомкам, поэтому различают
плодовитость самок
коэффициент воспроизводства в расчете на одну особь, независимо от пола. Например, популяция состоит из 500 самцов и 500 самок. За ед. времени появилась 1000 особей нулевого возраста. Удельная рождаемость составит 1 новорожденный на 1 особь, однако каждая самка как бы дала 2 потомков и каждый самец дал тоже 2 потомков.
Численность популяции может увеличиваться не только за счет рождаемости, но и за счет иммиграции особей из других популяций. Существуют зависимые и полузависимые популяции, которые
поддерживают свою численность именно за счет иммиграции.
5.Смертность. Понятие противоположное рождаемости. Различают абсолютную смертность – это количество особей, погибших за единицу времени. А также относительную или удель ную смертность
– это количество особей, умерших за единицу времени в расчете на 1 особь или 1000 особей. Характер смертности обычно описывается статистиками в виде кривых выживаемости, они показывают, какая часть новорожденных дожила до определенного возраста. Введено понятие мгновенная удельная смертность аналогично мгновенной удельной рождаемости.
Численность популяции уменьшается не только за счет смертности, но и за счет эмиграции.
ГЕНЕТИЧЕСКАЯ СТРУКТУРА ПОПУЛЯЦИИ.
По способу воспроизведения различают следующие типы популяций:
амфимиктические, здесь нормальное половое воспроизводство;
амфимиктические инбредные – когда брачные пары формируются из родственных особей. Различают а) инбридинг – близкородственное скрещивание у животных, б) инцухт – у растений в) инцест – у человека;
амфиктические панмиктические – когда происходит свободное скрещивание;
апомиктические – есть лидер партеногенеза без оплодотворение, гиногенеза (сперматозоиды только стимулируют оплодотворение) и андрогенеза (проникают два сперматозоида, ядро погибает и диплоидный организм образуется из 2 сперматозоидов);
клональные – отсутствие полового процесса, вегетативное размножение (полиэмбриония).
Различают такие понятия как аллелофонд – это совокупность аллелей в популяции, а также понятие генофонд – это совокупность всех генов данного вида. Сейчас существует термин фенофонд – совокупность всех фенотипов данного вида.
Закон Харди-Вайнберга применим только к идеальной популяции. В норме в популяциях происходит отбор, мутации, иммиграции и эмиграции. В свое время Ч. Дарвин выделяет несколько типов отбора:
движущий отбор – когда происходит изменение окружающей среды, и эти изменения приводят к отбору среди живущих особей. Например, бабочка - березовая пятница в районе Манчестера. Три формы: белая, серая, черная. Ее поедают воробьи, к примеру. Стволы берез покрываются сажей, белые видны лучше всего, следовательно, их поедают воробьи лучше всего выживают серые. Местность темнеет, загрязняется сажей, теперь серые лучше видны на темном фоне, чем черные , выживают черные.
стабилизирующий отбор
Шмальгаузен выделяет еще дизруптивный или разрывающий тип отбора. Та же бабочка, Манчестер, в сельской местности остаются светлые формы, в городе – черные. Образуются две популяции, которые со временем начинают отличаться друг от друга.
Серповидная клеточная анемия у человека – генетическая мутация, когда аминокислота валин заменилась на другую, при этом гемоглобин определенным образом заряжается, и молекулы начинают притягиваться, эритроцит сгибается в серп. Если человек гомозигота по этой мутации, то чаще всего он погибает после рождения. В таком случае мутация могла исчезнуть полностью, однако в Средиземноморье, например, раньше свирепствовала малярия, и оказалось, что гетерозиготы-носители этой мутации устойчивы к малярии. Это отбор в сторону гетерозиготности.
Отбор в сторону гомозиготности – группы крови. Если рассматривать количество людей, то окажется, что около 40% - I, 20-25% II или III и 5-7% четвертая (в Томске). В сторону Англии соотношения несколько другие – под 80% I, а вот в стороне Индии первой почти нет. В Америке у индейцев только первая гр. крови.
Как оказалось, это связано с тем, что происходит отбор по чувствительности к инфекциям (холера, оспа, чума
– к ним особо чувствительны люди с I группой крови). У антигенов на поверхности эритроцитов существуют предшественники, h-антиген, с помощью гена А или B, превращающийся в антиген (у людей с II, III или IV гр. крови). А у I так и остается h-антиген, и он оказался близок к антигенам возбудителей упомянутых выше болезней. Чем дальше от Индии, Гималаев, тем больше сохранялась I группа крови. Существует внутриутробный отбор у человека, когда нет антигенов на поверхности эритроцитов, соответственно у матери с I группой крови не происходит конфликта с плодом наподобие резус-конфликта.
Впервые ам. ученый Райт и отечественный ученый Н. П. Дубинин выдвинули идею, которую назвали генетико-автоматический процесс или дрейф генов. Эти ученые впервые доказали, что при образовании новых видов обязательно должна быть изоляция части популяции. Что происходит при изоляции? она м ожет быть нескольких типов:
географическая изоляция, например, в долине гор или на острове;
генетическая;
экологическая;
этологическая (у человека), когда существуют предрассудки в заключении брака по национальным, материальным и др. особенностям.
Вэтот изолят попадают не все аллели генов, характерные для этого вида, т. е. в изоляте обеднен генофонд. В изоляте наблюдается инбридинг либо инцест. При инбридинге возникает появление гомозиготных ситуаций по рецессивным генам. Некоторые рецессивные гены резко снижают жизнеспособность у человека, и при этом наблюдается гибель этих людей. Постепенно происходит генетическое обеднение изолята. В изоляте мутации очень быстро захватывают всю популяцию из -за близкородственных браков и малого количества особей, постепенно изолят может начать отличаться генетически от материнской популяции, и дальше может произойти генетическая нескрещиваемость между особями материнской и изолятной популяций. Постепенно может возникнуть новый вид. Впервые отеч. ученый С. С. Четверяков высказал мысль относительно того, что в человеческой популяции происходит накопление генетического груза, т. е. вредных мутаций в гомозиготном состоянии, и он считал, что это очень опасно для человеческой популяции. А почему происходит этот процесс? Он полагал, что в человеческой популяции снизился естественный отбор, возникающие мутации в виде рецессивных аллелей скрываются в гетерозиготном состоянии и не подвергаются отбору. И еще, благодаря прогрессу медицины сохраняются те особи в человеческой популяции, которые в принципе несут патологические гены. Происходит постепенное накопление таких людей. В настоящее время 15% новорожденных генетически дефектны, а с врожденными пороками развития более 25%. Он считал, что сохраняя этих людей и позволяя им рожать детей , человечество может привести себя к гибели или ужасным последствиям.
Вчеловеческой популяции частота мутаций 105-107 на поколение, т. е у каждого человека 6-10% гамет мутантно изменены.
Обычно при кровнородственных браках говорят об инбредной депрессии у потомков. Если, например, частота рождения детей с фенилкетонурией составляет 1 случай на 15000, то при близкородственном браке эта величина увеличивается в 100 раз. Мерой генетических последствий инбридинга служит коэффициент инбридинга. У детей одной супружеской пары вероятность одинаковых аллелей в одном локусе равна 1/2. Если вступят в брак их дети, то 1/4. Двоюродные 1/16. В изоляте огромную роль играет т. н. эффект родоначальника. Например, в США есть секта мормонов, выходцев из Голландии, из-за инбридинга в этой секте несколько тысяч карликов, склонных к дистрофии. Описано поселение в горах Швейцарии, там на 5000 жителей 50 глухонемых.
Аутбридинг – неродственные браки. При этом аутбридинг поддерживается иммиграцией и эмиграцией. Когда браки совершаются между людьми, живущими очень далеко друг от друга, то проявляется эффект который называется акселерацией, ученые полагают, что акселерация возникает в том случае, когда по большинству генов у человека гетерозиготные состояния.
Интенсивность естественного отбора у современного человека довольно высока, примерно 50% зачатий оканчиваются спонтанными абортами, 3% - мертворождениями, ранняя детская смертность - 2%. Если смотреть хромосомы у абортированных плодов, то более чем в 30% случаев обнаруживаются цитогенетические аномалии, а почти в 50% случаях – генные точковые мутации. Примерно 75% людей не вносят вклад в генофонд будущих поколений, в брак не вступает примерно 20%, около 15% бесплодны, неполноценных тоже часто не выносят. Помимо естественного отбора в человеческих популяциях действует еще и контротбор – это сохранение некоторых типов мутаций (серповидная клеточная анемия).
Вэволюции человеческая популяция проходила несколько раз т. н. бутылочное горлышко (эпидемии, катастрофы, ЧС), когда от человечества оставалось малое количество особей. Это явление довольно часто приводит к отсеканию других генотипов, все начинает развиваться иначе. Ледниковый период явился одним из бутылочных горлышек.
Лекция 12. ИЗМЕНЧИВОСТЬ.
Изменчивость является вторым свойством, рассматриваемым в генетике. Генетика – наука о наследственности и ее изменчивости.
1809 г. – французский ученый Жан Батист Ламарк в своей книге «Философия зоологии» впервые приводит два фактора изменчивости, которые ведут к эволюции организмов:
1.все живое стремится к самоусовершенствованию;
2.тренировка органов ведет к их развитию, а отсутствие – к деградации, исчезновению.
1859 г. – Чарльз Дарвин в книге «Происхождение видов путем естественного отбора» приводит пять факторов, ведущих к эволюции:
1.все живое обладает наследственностью;
2.наследственность может изменяться;
3.имеет место избыточное воспроизводство потомства;
4.между потомками идет борьба за существование;
5.выживают наиболее приспособленные – это естественный отбор. Дарвин выделил следующие типы изменчивости:
1.Групповая или определенная изменчивость. Этот тип изменчивости адаптивный, возникает как адаптация к условиям окружающей среды. Не передается по наследству. В настоящее время данный тип называется фенотипической или модификационной изменчивостью.
2.Индивидуальная или неопределенная изменчивость. Возникает внезапно, зачастую неадаптивна, и только случайным образом она может стать тем фактором, который дает преимущества этому организму.
Ученый Дженкинс подверг критике Дарвина, сказав, что этот тип изменчивости в ряду поколений должен исчезать. И только Мендель впервые доказывает, что наследственность – это дискретная единица, которая возникла, не исчезая.
Внастоящее время изменчивость подразделяют на 2 типа:
1.модификационная или фенотипическая; различают следующие понятия: норма реакции модификационной изменчивости. Модификационная изменчивость может быть в широких пределах и зачастую это не жизненно важные признаки (например, загар тела у человека), и в узких пределах, часто жизненно важные признаки (например, содержание сахара в крови).
2.генотипическая;
комбинативная
мутационная
В 1927 ученый Тимофеев-Ресовский вводит понятия:
1.Экспрессивность признака – степень его выраженности, например, брахидактилия (укорочение фаланг пальцев). При наличии одного и того же генотипа брахидактилия может быть как резко выражена, так и мало отличима от нормы.
2.Пенетрантность признака или гена – это процент особей, у которых данный доминантный ген проявляется, т. е. не всегда доминантный ген может проявляться как признак. Например, подагра (отложение солей мочевой кислоты на суставах). У мужчин пенетрантность 100%, у женщин 20%. У человека можно ясно выявить модификационную изменчивость при изучении однояйцевых, двуяйцевых близнецов. Те изменения, которые встречаются чаще у однояйцевых близнецов , чаще являются генотипическими и наоборот.
Термины генотип и фенотип был впервые предложен в 1911 датским ученым Иогансеном. С этого
момента стали рассматривать отдельно фенотипическую и генотипическую изменчивость.
Учение Ламарка можно было бы забыть, однако периодически появляются идеи, являющиеся, по сути, ламаркистскими. Они заключаются в следующем: условия окружающей среды вызывают изменения, которые являются приспособительными и передаются потомству. По Дарвину же это не так, изменения возникают случайно и борьба за существование, а также естественный отбор отбирают только те, которые адаптивны.
В 30х гг. прошлого века у нас появилось учение Лысенко, которого поддержало руководство страны, он предлагал, изменяя условия окружающей среды, получать новые сорта и породы животных. Например, телят выпаивали сливками, чтобы получить жирномолочную породу, и действительно, выращенная корова давала жирное молоко, но если делать это не каждый раз, то жирномолочность исчезала. Борьба лысенковцев с генетиками привела к тому, что практически все генетики были репрессированы. Первый учебник по генетике вышел только в 1961 г. Ныне существуют и новые попытки по этому поводу. Есть данные, что внешняя среда все-таки оказывает влияние.
Генотипическая изменчивость комбинативна, связана с перекомбинацией ген материала у потомков, т.е сами гены не изменяются, но потомки получают разную комбинацию генов, это происходит из за следующих процессов:
равная вероятность расхождения гомологичных хромосом в будущих половых клетках;
перекомбинация ген материала в результате конъюгации и кроссинговера гомологичных хромосом;
равная вероятность слияния в зиготе мужской и женской половых клеток, приводит к тому, что разнообразие потомков только в одной семье (муж и жена) составляет 24^12.
Мутационная изменчивость связана с изменениями в структуре наследственного материала, и
возникающие изменения передаются потомкам, это могут быть следующие поколения клеток или же следующие поколения особей. Различают варианты генных мутаций, первое такое описание по морфологическим критериям сделал в 1932 г. американский ученый Меллер. Он, изучая дрозофил, выделил следующие морфологические мутации:
гипоморфные мутации – происходит ослабление выраженности признака;
аморфные мутации – признак вообще исчезает;
антиморфные мутации – происходит появление противоположного признака;
неоморфные мутации – новые признаки;
гиперморфные – чрезмерное развитие признака.
Кроме этого различают прямые и обратные мутации, т. е. возникшая мутация может вернуться в
исходное состояние.
Выделяют соматические и генеративные мутации. Соматические происходят в неполовых клетках тела, они не передаются по наследству потомкам, но передаются следующим поколениям клеток организма, накопление их резко снижает жизнеспособность, наблюдаются аномалии функционирования органов, где они накапливаются. Соматические мутации часто приводят к раковой трансформации. Генеративные мутации происходят в половых клетках, чаще всего, на первых стадиях деления в сперматогониях, овогониях. Они могут вызывать появление генетически аномального потомства.
Иногда вводят такие понятия как мутации биохимические, морфологические, физиологические и т. д. Спонтанные и индуцированные мутации. Спонтанные или самопроизвольные мутации могут
возникнуть от самых разных причин, дело в том, что колебательные движения любых молекул в организме, в т. ч. и ДНК иногда спонтанно могут привести к изменению структуры. Этому может способствовать простое повышение температуры. Кроме того, в естественных условиях мы подвергаемся естественному радиоактивному воздействию от Солнца, разных горных пород, космического излучения и др. И это может вызвать нарушения в генетическом материале. Индуцированные мутации возникают под влиянием:
физических факторов;
химических факторов;
биологических факторов.
В1925 г. отечественный ученый Надсон и Филиппов впервые в экспериментах на дрожжевых клетках установили, что лучи радия могут вызывать мутации у этих дрожжевых клеток. В 1927 Меллер обнаруживает, что рентгеновские лучи вызывают мутации у дрозофилы. Различают
электромагнитные или волновые физические мутагены – это рентгеновские лучи, гамма-лучи (γ-лучи), космические лучи;
корпускулярные физические мутагены – это β-часицы (электроны и позитроны), протоны, нейтроны
(быстрые и тепловые) и α-частицы (ядра атома гелия).
Помимо этого сюда же относятся УФ лучи. Различают ионизирующее излучение и неионизирующее, ионизация, как правило, происходит в молекулах воды, образуются ионы ОН- и Н+, и возникают перекиси Н2О2 и НО2 . Эти перекиси способны разрушать молекулы ДНК. Под влиянием УФ лучей возникают т. н. тиминовые димеры: между двумя тиминами в одной цепи ДНК формируется особая связь, и это нарушает процесс репликации ДНК.
Химические мутагены. Впервые супермутагенное действие химических веществ было обнаружено в 1949 г. английской ученой Шарлоттой Ауэрбах и нашим ученых Ильей Рапопортом.
Наиболее сильное мутагенное действие оказывает т. н. алкилирующие соединения, которые способны внедрить в ДНК алкильные группы, например СН3- или С2Н5-. К таким соединениям относятся диметилсульфанат, диэтилсульфанат, иприт, этиленимин. К супермутагенным соединениям относится нитрозаалкилмочевина, нитрозаметилмочевина и этилметансульфанат. Последние три применяются в медицинской практике для лечения раковых больных.
Вторая группа – аналоги азотистых оснований, т. е. вещества, которые химически похожи на стандартные азотистые основания А, Т, Г, Ц и при репликации вместо нормального азотистого основания в ДНК попадает аналог, это в дальнейшем приводит к мутационным изменениям. К ним относятся 5-бромурацил, 5-фтордезоксиуридин, 8-азогуанин, кофеин.
Акридиновые красители. Акридиновые красители под влиянием УФ лучей начинают особо светиться, испускать собственный свет, как оказалось, они способны разрывать связь между аденином и тимином, это называется интеркаляция, и встраиваться между нитями ДНК. И естественно, при репликации эти места повреждаются. Сюда относится акридиновый желтый, акридиновый оранжевый, профлавин и 5-аминоакридин.
Сборная. Например, колхицин - алкалоид в луковицах колхикума, это вещество способно разрушать нити ахроматинового аппарата, что приводит к неправильному расхождению хромосом. Формальдегид и все соединения, связанные с азотной кислотой.
Современная классификация мутаций связана с типами мутаций в генетическом аппарате, различают генные или точковые мутации, хромосомные мутации и геномные мутации. Генные мутации обусловлены изменениями в структуре ДНК, чаще всего это вставки или выпадение нуклеотидов, замена одного нуклеотида на другой. Различают следующие типы:
1.трансверсии – пуриновое азотистое основание замещается на пиримидиновое, и наоборот;
2.трансзиции – пурин замещается на пурин, пиримидин замещается на другой пиримидин;
3.миссенс-мутация – один значимый кодон замещается на другой тоже значимый кодон, это приводит при биосинтезе белка к замене одной аминокислоты на другую, например, серповидная клеточная анемия, здесь в α-цепи гемоглобина в 6 положении (всего 141 аминокислота) глутаминовая кислота замещается на валин;
4.сейсменс мутация – один вырожденный кодон замещается на другой вырожденный. Вырожденными называют те кодоны, которые определяют одну и ту же аминокислоту в молекуле белка, причем это не приводит ни к каким изменениям в белковой молекуле;
5.нонсенс-мутация. Имеется три нонсенс стоп-кодона, которые определяют конец считывания информации, это УАА, его называют охра, УАГ его называют амбер, УГА – опал. Если появляется такой кодон, то в этом месте обрывается синтез, если это происходит в самом начале, то дефект гораздо сильнее, чем если в конце гена;
6.сдвиг рамки считывания. При вставке или выпадении одного или нескольких кодонов порядок считывания кодонов может резко измениться и изменить всю белковую молекулу;
7.сплайсинговые мутации. Сплайсинг – это процесс вырезания из РНК интронов и сшивание экзонов вместе, после чего с этой РНК начинается синтез белка, если сплайсинг произойдет неверно, то может измениться весь смысл этой РНК и белка соответственно;
8.регуляторные мутации. Из положений оперонной системы Жакома Моно существуют генырегуляторы и оперонные системы, если мутация произойдет в регуляторном гене, и нарушится система регуляции, то биосинтез этого оперона может происходить безостановочно или наоборот полностью блокируется;
9.динамические мутации или мутации-экспансии. В последнее время было открыто, что наши гены отличаются у разных людей по т. н. тринуклеотид-повторам ЦГЦ. Пока не совсем понятно, как это происходит, но порой у человека связаны с этим некоторые заболевания, например, такое заболевание как ломкость Х-хромосомы человека. Как правило, эти люди отстают в умственном развитии, имеют проблемы в генеративном развитии. При исследовании была обнаружена область, где ЦГЦ у нормального человека около 20, а у больного – тысячи, это обуславливает ломкость Х-хромосомы.
10.транспозоны или прыгающие гены. Впервые Барбара МакКлинток обнаружила на кукурузе генетические элементы, которые могут перемещаться по геному, они встраиваются в другие гены и
блокируют их. Она это установила на примере окраски зерна кукурузы Оказалось, что существует второй генетический элемент (транспозон), который заставляет
инсекцию «прыгать» то в ген, то из гена. У него есть два фермента – транспозаза и резольваза, которые как бы вырезают эту инсекцию и перемещают ее, встраивая в ген, такие генетические элементы были открыты и у человека. Барбара МакКлинток получила Нобелевскую премию за свое открытие в 98 лет.

Лекция 13. ХРОМОСОМНЫЕ МУТАЦИИ.
Хромосомные мутации или хромосомные аберрации связаны с изменение структуры хромосом, при этом тип хромосомной мутации зависит от того, в какую стадию клеточного цикла произошло нарушение структуры хромосом.
На стадии G1 (пресинтетический период) хромосома
однохроматидна, пройдя через стадию синтеза, в G2-стадии хромосома двухроматидна, и затем в митозе происходит разделение хроматид. Клетка вновь вступает в G1-период.
Если разрыв структуры хромосомы произошел в G1-период, то после стадии синтеза образуются двойные фрагменты. Такой тип называется хромосомным типом аберрации.
Если разрыв произошел в конце S-стадии или в G2, когда хромосома двухроматидна, то возникает иной тип, когда только одна хроматида будет повреждена, а вторая будет целой, в этом случае имеет место хроматидный тип аберрации.
Редко в одном и том же месте у двухроматидной хромосомы возникают повреждения, очень напоминающие хромосомный тип аберрации, в этом случае их называют изохроматидный тип аберрации.
Среди аберраций хромосомного и хроматидного типа выделяют простые и обменные аберрации. Нарушение одной или нескольких хромосом, когда в результате одного разрыва появляются фрагменты, и эти фрагменты различаются. Фрагмент, который содержит центромерную область, называется центрическим, он может нормально расходиться в митозе и мейозе, потому что имеется центромера, а фрагмент, который не имеет центромеры, зачастую сперва образует собственное микроядро, а затем нуклеазы клетки его растворяют (ядро), т.е. происходит утрата этого хромосомного материала.
Различают терминальные, концевые и интерстициальные фрагменты (средний участок хромосомы). Обменные аберрации очень разнообразны. Различают:
межхромосомные обмены, когда обменивается материал между разными хромосомами;
внутрихромосомные обмены.
Кроме того, обмены могут быть симметричными и асимметричными. Асимметричные обмены
приводят к образованию ацентрических фрагментов, которые потом лизируются. Симметрические обмены – когда две хромосомы меняются друг с другом участками, и ацентрические фрагменты не образуются. Внутрихромосомные обмены могут быть внутри одного плеча хромосомы или между верхним и нижним плечом хромосомы. Внутриплечевые и межплечевые обмены. Обмены могут быть простыми и сложными. Кроме того, они могут быть полными (реципрокными), когда все фрагменты успешно соединились и неполными (нереципрокными). Кроме этого, выделяют стабильные и нестабильные обмены. При стабильном обмене перестроенные хромосомы успешно проходят митоз и мейоз, а при нестабильном наблюдается потеря хромосом.
Среди структурных аберраций хромосом выделяют следующие:
1.дефишенси – утрата концевого участка хромосомы;
2.делеции – утрата внутреннего участка хромосомы;
3.инверсии – переворот участков хромосомы на 180°. Различают
a)парацентрические инверсии, когда инверсия происходит внутри какого-то плеча и не затрагивает центромерную область, в этом случае конфигурация хромосомы не меняется (мета-, субмета- и акроцентрические хромосомы);
b)перицентрические инверсии, когда происходит межплечевой обмен и в этом случае может измениться конфигурация хромосомы, т. е. метацентрическая хромосома может превратиться в акроцентрическую и наоборот.
4.дупликации – удвоение участка хромосомы;
5.транслокационные обмены (транслокации) – могут быть внутри- и межхромосомными. Отдельно среди транслокаций выделяют т. н. роберсоновские транслокации, когда акроцентрические хромосомы (13, 14, 15, 21, 22, у). Когда, например, 21 хромосома садится на 13, формируется субметацентрик. В эволюции человека такая перестройка сыграла огромную роль. У приматов (шимпанзе, гориллы, орангутанга) 48
1
хромосом, а у человека – 46. И у них имеются дополнительные две акроцентрические хромосомы, а у нас есть вторая хромосома, которой нет у обезьян. Доказано, что она произошла благодаря роберсоновской транслокации акроцентрических хромосом. И такая транслокация у какой-то группы древних обезьян привела к генетической изоляции. Скрещивание было возможным, но потомки были бесплодны.
Ученые считают, что эволюционные процессы происходят следующим образом: у низших организмов много акроцентрических хромосом, затем начинаются процессы слияния с образованием метацентриков, у метацентриков происходят инверсии, и они превращаются в большие метацентрики, и чем дальше в эволюции, тем хромосомы становятся крупнее.
Аберрации хромосомного типа могут быть:
1.парными или интерстициальными, они могут быть точечными или очень длинными. Они лишены центромеры, образуются в G1-период;
2.кольцевые хромосомы, как правило, интерстициальны, т. е. внутренний фрагмент может замкнуться на себя и образовать кольцо, эта хромосома имеет центромеру, но для этого обязательно должно быть два разрыва, должны оборваться два терминальных участка.
3.дицентрические хромосомы, когда образуется определенная конфигурация в результате
нарушений в G1-период.
Как правило, кольцевые и дицентрические хромосомы образуются в результате радиационных воздействий, подсчитав у человека количество в его клетках таких хромосом, можно четко сказать, какую дозу облучения он получил, это т. н. биологический метод детекции радиационного воздействия
МУТАГЕНЕЗ И РЕПАРАЦИЯ ДНК Большая часть повреждений молекулы ДНК способна восстановиться с помощью
специальных ферментов, этот процесс называется репарация (что значит «зашиваю»). Различают три основные возможности формирования предмутационных повреждений:
1.мутаген может включиться в ДНК вместо нормального азотистого основания. Так мутаген 2- аминопурин является аналогом аденина, и когда этот мутаген встраивается, он начинает спариваться с тимином или цитозином. Это ведет к появлению транзиции: А-Т замещается на Г-Ц или наоборот;
2.мутаген может модифицировать азотистое основание. Пример: такой мутаген как гидроксиламин. Он модифицирует цитозин, превращая его в урацил. Алкилирующие мутагены вызывают модификацию гуанина;
3.мутаген может повредить азотистое основание, и это затруднит спаривание этого азотистого основания. Примером служит появление тиминовых димеров, когда тимины образуют друг с другом особую связь.
Все известные в настоящее время виды репарации ДНК можно подразделить на:
a)конститутивные, т. е. постоянно действующие,
b)индуцируемые, т. е. возникающие под влиянием какого-то фактор.
Возможность репарации ДНК была установлена в 1949 году независимо тремя учеными – Дюльбекко, Кельнер, Ковалев. Они установили, что мутации, которые возникают у простейших под влиянием УФ-лучей восстанавливаются, если этих простейших поместить на видимый свет. Это было названо фотореактивацией.
В 1964 г. трое ученых Сетлоу, Бойс и Кэрриер обнаружили темновую репарацию ДНК, т. е. восстановление ДНК в темноте.
Всего в организме насчитано около 50 генов, которые ведают репарацией ДНК. Выделяют т. н. дорепликативную репарацию, т. е. происходящую в G1-период. Она включает как фотореактивацию, так и темновую репарацию, при этом происходит вырезание поврежденного участка, это явление называется эксцизией поврежденного участка, и на основе неповрежденной нити ДНК под влиянием особых ферментов происходит правильное восстановление поврежденного участка.
2
Кроме этого имеется и пострепликативная или внутрирепликативная репарация, т. е. происходящая в S или G2-период. Считается, что этот тип репарации основан на процессах рекомбинации, т. е. восстановление идет путем кроссинговера и перекомбинации генетического материала.
Третий тип репарации это индуцируемая репарация, имеются особые гены, которые включаются при повреждениях ДНК, особенно если этих повреждений много. Иногда называют эту репарацию S.O.S.-репарацией. При этом восстановление может идти неправильно, хаотично, что приводит к образованию мутаций.
Наибольшее количество мутаций восстанавливается с помощью дорепликативной репарации, она или одноэтапна (проходит с помощью одного фермента) или многоэтапна, т. е. здесь задействовано несколько ферментов, наиболее хорошо изучен фермент пурин-инсертаза и ДНКлигаза, которые восстанавливают пуриновые поврежденные сайты. Многоэтапные репарации – это, в основном, темновая эксцизионная репарация. Один фермент удаляет поврежденный участок, а другой фермент восстанавливает его от другой нити ДНК.
Фотореактивация связана с ферментом дезоксиориботидпиримидинфотолиазой. Этот фермент присоединяется к тиминовым димерам, разрывает образовавшуюся между тиминами связь и восстанавливает поврежденные участки. Как правило, эта система может восстановить до 90% поражений ДНК, однако 10% могут миновать эту систему, и появятся генетически дефектные молекулы, их судьба может быть разная, все зависит от степени повреждения. У нормального здорового человека 2-3% генетически измененных клеток. Эти клетки могут функционировать, но у них начинает изменятся внешняя поверхность клеточной оболочки, и особые клетки-киллеры, образующиеся, в основном, в тимусе человека их воспринимают как чужие и с помощью особых ферментов уничтожают. Кроме того известна система апоптоза клеток. Во время апоптоза включаются особые гены, и образуются нуклеазы, растворяющие ядро клетки. Т. о. при функционировании этих систем организм постепенно очищается от генетических изменений.
3

Лекция 14. ГЕНОМНЫЕ МУТАЦИИ У ЧЕЛОВЕКА Геномные мутации связаны с изменениями в числе хромосом. Различают следующие типы
геномных мутаций:
Анеуплоидии – увеличение или уменьшение числа хромосом на одну или несколько, среди них выделяют гипоплоидии (уменьшение числа хромосом) и гиперплоидии (увеличение числа
хромосом). Гипоплоидии встречаются как:
a)моносомии – когда отсутствует одна из пары гомологичных хромосом;
b)нуллисомии – отсутствуют обе гомологичные хромосомы.
Среди гиперплоидий встречаются:
a)трисомии – когда имеется три гомологичные хромосомы;
b)тетрасомии – четыре гомологичные хромосомы.
Полиплоидии – кратное увеличение числа гаплоидных наборов хромосом. Различают:
a)триплоидии (три гаплоидных набора хромосом, у человека это 69 хромосом),
b)тетраплоидии (4 набора, у человека это 92 хромосомы).
Полиплоидия в соматических клетках у человека встречается довольно часто. Полиплоидными являются клетки печени и клетки сердца. Полиплоидия образуется в печени, в анафазе хромосомы расходятся к разным полюсам, образуется два ядра в клетке, однако в телофазе не происходит цитотомии, т. е. не образуется перегородка. Результат – двуядерная клетка. Затем два ядра снова вступают в митоз, их хромосомы объединяются (92 хромосомы), снова образуется два ядра по 92 хромосомы. И вновь нет цитотомии. Клетки печени могут содержать 16 или даже 32 набора гомологичных хромосом. Это необходимо потому, что печень является детоксикационным органом, через нее проходят практически все поступающие в организм мутагены. И даже если произошла мутация, то она ни к чему не приводит, так как каждый ген представлен 16 или 32 вариантами. Мутация не может проявиться. В животном и растительном мире полиплоиды встречаются на границах существования данного вида (в пустынях, далеко на Севере), так как полиплоидия дает преимущества перед другим организмами. У человека полиплоидия среди новорожденных также встречается (особенно триплоидия, т. е. имеющие 69 хромосом, дети). Часто это связано с тем, что в оплодотворенной яйцеклетке не произошло расхождение хромосом, и после того как происходит оплодотворение спермием, плод будет иметь 69 хромосом. Реже встречаются тетраплоиды, они чаще всего результат оплодотворения аномальным диплоидным спермием диплоидной яйцеклетки.
ХРОМОСОМНЫЕ БОЛЕЗНИ ЧЕЛОВЕКА Хромосомные болезни обусловлены изменениями числа и структуры хромосом. Частота среди
новорожденных около 1%. Не по всем хромосомам встречаются аномальные новорожденные, некоторые изменения летальны, их можно встретить среди выкидышей спонтанных абортов. Около 20% выкидышей имеют аномальный хромосомный набор. Изменение числа хромосом происходит при неправильном расхождении в мейозе, чаще это бывает у матерей, которым более 35 лет. Нерасхождению хромосом способствует:
Ионизирующее излучение
Химические вещества
Вирусные инфекции. Например, этому способствует вирус краснухи, вирус гепатита, токсоплазма и некоторые другие биологические мутагены. К ним относится также изменение гормонального фона человека.
Хромосомные мутации подразделяются на:
1)Болезни, обусловленные нарушениями числа аутосом;
2)болезни, связанные с изменениями числа гоносом, т. е. половых хромосом Х и Y;
3)болезни, связанные с появлением полиплоидного набора хромосом;
4)болезни, связанные с изменениями структуры хромосом и с перестройками хромосом. Структурные нарушения бывают
транслокационные;
дупликационные;
делеционные;
1
появление изохромосом, когда верхние и нижние плечи одинаковы;
возникновение кольцевых и дицентрических хромосом.
Трисомии, когда хромосома представлена тремя гомологами, моносомии когда гомолог только один. Типы наследственности:
1. Аутосомно-доминантный признак
обнаруживается в каждом поколении
этот признак наследуется половиной детей
мужские и женские потомки наследуют одинаково
родители в равной степени могут передать этот признак потомкам.
2. |
Аутосомно-рецессивный |
|
признак передается чаще через поколение |
|
признак может не проявляться у родителей, чаще всего это гетерозиготы |
|
обнаруживается у ¼ детей, рецессивных гомозигот, если оба родителя больны, то все дети |
|
больны, если один из родителей болен, а другой гетерозиготен, то больны половина детей, |
|
потомки мужского и женсого пола наследуют признак одинаково |
|
оба родителя в равной степени могут передавать этот признак |
3. |
Наследование, сцепленное с Х-хромосомой при рецессивном признаке |
|
мужчины наследуют чаще, чем женщины |
|
девочки как носители могут унаследовать от отца |
|
если оба супруга здоровы, но при этом мать гетерозиготна, 50% сыновей больны, а дочери |
|
здоровы |
|
прослеживается чередование больных мужчин в поколении |
4. |
Наследование, сцепленное с Х-хромосомой, когда признак доминантен |
|
мужчины наследуют реже, чем женщины |
|
если признак есть только у супруги и мать гомозиготна, то наследуют все дети, если мать |
|
гетерозиготна, то половина |
|
если признак есть только у супруга, то наследует только его дочь |
5. |
Наследование, сцепленное с Y-хромосомой |
страдают только сыновья, если болен отец.
Среди наследственных заболеваний у человека выделяют следующие типы:
хромосомные болезни
болезни обмена веществ
нарушения иммунитета
болезни с преимущественно поражением эндокринной системы
болезни крови
нарушение функций посек
болезни НС
поражения глаз
болезни ПВС
1.Синдром Шершевского-Тернера. Установлено, что Y-хромосома особо чувствительна к мутагенным факторам, если Х-хромосома содержит 4000 генов, то Y-хромосома – 33 гена. Имеется тенденция к уменьшению Y-хромосомы. У некоторых народов, например, у японцев Y-хромосома очень мала, с этим некоторые ученые связывают распространение гомосексуализма у людей. Дегенерация Y-хромосомы выявлена у 10-15% бесплодных мужчин. Данный синдром встречается как среди женщин, так и среди мужчин. В отличие от Y-хромосомы Х-хромосома более стойкая, при синдроме Шершевского-Тернера пол определяется в системе х0 определения пола.
2.Синдром Патау или трисомия по 13-й хромосоме. Встречается с частотой 1:25000 живорожденных детей, риск увеличивается с возрастом женщины. Около 20% аномалии связано с транслокацией избыточной хромосомы. Если в семье появляется такая аномалия, то велика вероятность появления людей с этим синдромом, поэтому необходимо генетическое
2

консультирование и обследование. Симптомы: маленькая голова, уменьшенные глаза, поражения кожи, аномальные уши, заячья губа, волчья пасть, шестипалость, аномалии сердца и мочеполовой системы. Большинство детей умирает в течение года, выжившие резко отстают в физическом и умственном развитии, лечение не предусмотрено.
3.Синдром Эдвардса или трисомия по 18-й хромосоме. Встречается с частотой 1:6600 живорожденных. Почти 80% девочки. Мальчики часто не доживают до рождения. Также чаще
рождают женщины после 35 лет. Встречается три типа:
a)регулярная форма трисомии, когда у человека 47 хромосом, три 18-х хромосомы
b)транслокационная форма, примерно 5% случаев, когда 18 хромосома присоединяется к какойто другой хромосоме
c)мозаичная форма, когда поражена только часть клеток организма, при этом если часть клеток
нормальны, то тяжесть заболевания будет другой.
Симптомы: чаще дети рождаются переношенными, но с низким развитием плода, удлиненная голова, т. н. долихоцефалия и у основании находятся как бы выпячивания черепа, маленькие глаза, микрогнатия, маленький рот, подбородок, деформированные уши, короткая грудина. Характерный признак – сжатый кулак и указательный палец перекрывает другие пальцы. Большой палец часто отсутствует. Пороки сердца, почек, легких, грыжи. Сильная умственная отсталость, живет часто не больше года, редко доживают до подросткового возраста, лечения не существует.
4.Синдром Дауна или трисомия по 21-й хромосоме. Один случай из 7-10 тыс. живорожденных, зависит от возраста матери и отца. Существует регулярная классическая форма (во всех клетках тела три 21-е хромосомы, всего 47 хромосом), 4% имеют транслокационную форму, если обнаруживается транслокационная форма, то необходимо смотреть хромосомы родителей, у одного из них 45 хромосом – сбалансированая транслокация – 21 хромосома прицепилась к какой-то другой хромосоме. В этом случае прогноз на других детей неблагоприятный, в 50% случаев последующие дети могут быть с синдромом Дауна. Если это регулярная форма, то чаще всего это случайное явление, и следующие дети могут быть совершенно нормальными. Существует также мозаичная форма, когда не все клетки поражены, она возникает не у родителей, а у эмбриона при первых делениях клеток может произойти неправильное расхождение хромосом, в одной 47 в другой 45 хромосом, все зависит от того, какое количество клеток поражено, при этом иногда человек с синдромом Дауна может учиться в ВУЗе. Для определения унаследованной транслокации необходимо обследовать хромосомы обоих родителей. Симптомы: характерное плоское лицо, около глаз имеется эпикан (складка), макроглассия, плоская переносица, косоглазие, открытый рот, пигментные пятна на радужке, увеличен поперечный размер головы, плоский затылок, низко посаженные уши, зубные аномалии, короткая складчатая шея, очень подвижные суставы, воронковидная грудь, на ладонях обезьянья складка, умственная отсталость, которая может быть выражена по-разному. Дауны склонны к болезням слуха и дыхательных путей, лейкемии. У них нарушен иммунитет, и чаще всего они погибают от инфекций. Хромосомные нарушения нельзя устранить, иногда применяют препараты для стимулирования умственной деятельности (наподобие глицина), создают специальные группы для их адаптации.
5.Синдром «кошачьего крика» – делеция короткого плеча 5-й хромосомы. Частота 1:45000 новорожденных, характерен плач, напоминающий кошачье мяуканье, лунообразное лицо, мышечная гипотония, микроцефалия, деформированные уши, т. н. антимонголоидный разрез глаз, косоглазие, атрофия зрительного нерва, у них изменена глотка с необычной складчатостью, уменьшен надгортанник что обуславливает своеобразный плач. Живут обычно не более 10 лет.
6.Синдром Клайнфельтера. 47 хромосом, ХХУ определение пола, бывает больше Х-хромосом. Частота 1:(500-750) новорожденных мальчиков. В присутствии У-хромосомы определяют развитие мужского пола. Как правило, до пубертатного возраста они ничем не отличаются, далее наступает недоразвитие семенников и вторичных половых признаков, стерильны, сперматогенез нарушен, у них повышен женский фолликулостимулирующий гормон, который обычно можно определить в моче. Характерна гинекомастия – увеличенные млечные железы, (но это не молочная железа как таковая, там находится соединительная ткань). Складываются евнухоидные пропорции тела: узкие плечи, широкий таз, слабое оволосение лица, лобка,
3
подмышек. Обычно имеют высокий рост, безынициативны, поддаются внушению, их обычно привлекают к себе антисоциальные коллективы, могут часто попадать в тюрьмы. Может быть выражена дебильность, чем больше Х-хромосом, тем больше дебильность. Обычно этим людям вводят метилтестостерон (мужской гормон), начинают курс с 10 лет, однако чаще выявляют, когда призывают в армию.
7.Синдром дубль-У или синдром агрессивности. 47 хромосом ХУУ. Впервые описали среди заключенных в Дании в 1960, частота 1:1000(250). Средний рост 186 см, высокие, характерно агрессивное поведение. Часто неправильное строение зубов, увеличение нижней челюсти, часто увеличен уровень андрогенов, тестостеронов, могут иметь детей, интеллектуальное недоразвитие, часто попадают в тюрьмы за антисоциальное поведение
8.Трисомия Х, частота 1:1000. Часто попадают в психиатрические больницы, но мало чем отличаются от нормальной женщины, иногда неглубокая умственная отсталость (75%), часта шизофрения, высокое твердое небо, гортанный голос, у мизинцев выраженная клинодактилия, эпикант, встречаются высокого роста. Могут иметь детей, но может быть и бесплодие. Может быть 4 и даже 5 Х-хромосом, чем больше Х-хромосом, тем ниже интеллект.
4

Лекция 15. МЕДИКО-ГЕНЕТИЧЕСКОЕ КОНСУЛЬТИРОВАНИЕ. ОСНОВНЫЕ МЕТОДЫ МЕДИЦИНСКОЙ ГЕНЕТИКИ.
Основная цель МГК – это предупреждение рождения генетически больного ребенка. Главные методы МГК:
1.точное установление наследственной патологии;
2.дородовая (пренатальная) диагностика врожденных и наследственных заболеваний различными методами:
ультразвуковыми
цитогенетическими
биохимическими
молекулярно-генетическими
3.определение типа наследования заболевания:
рецессивное аутосомное
рецессивное, сцепленное с полом
доминантное аутосомное
доминантное, сцепленное с полом
гоносомное;
4.основное определение, оценка величины риска рождения больного ребенка и оказание помощи родителям в принятии решения;
5.пропаганда медико-генетических знаний среди населения.
Поводом для обращения в МГК могут быть
1.Рождение ребенка с врожденными пороками развития, умственной, физической отсталостью, слепотой, глухотой и другими отклонениями от нормы. Как правило, родители хотят иметь здорового ребенка, и поэтому они должны решить сохранить ребенка или нет.
2.Спонтанные аборты, выкидыши, мертворождения.
3.Близкородственные браки.
4.Неблагоприятные течения беременности.
5.Работа родителей во вредном производстве.
6.Несовместимость супружеской пары по резус конфликту.
7.Возраст родителей: матери более 35лет, отец – более 40.
МГК врача включает 4 этапа:
1.постановка диагноза
2.установление прогноза для следующих поколений
3.заключение
4.совет родителям
Точный диагноз – необходимое условие для любой консультации, если это хорошо изученное заболевание, часто встречаемое, например, синдром Дауна, то диагноз ясен сразу. Но часто необходимо прибегнуть к более точным современным методам: биохимическим, иммунологическим, генетическим.
Один из первых методов, который использует врач-генетик – это генеалогический метод, т. е. он собирает досье о своих пациентах. В первую очередь, это относится к тому из супругов, в родословной которого отмечались наследственные патологии. Более сложный случай, например, при рождении ребенка с множественными патологиями, правильный диагноз может быть поставлен только с использованием специальных методов. Родословная составляется зачастую либо со слов обследуемых родителей, либо путем анкетирования. При этом врачу нужно не забывать психологическую сторону этой проблемы. Очень часто один из родителей скрывает, что у него были наследственные аномалии. И поэтому иногда очень сложно составить родословную.
Врач-генетик может прогнозировать вероятность двумя методами:
1.Априорная вероятность, когда врач высказывает заключение только на основе изучения родословной родителей без учета изучения их детей
2.Апостериорная вероятность, когда обследованы дети и плюс априорная вероятность.
Пробантом или пропозитом называют человека, с которого врач начинает составлять родословную. Лучше, если врач лично обследует большую часть родственников. Следует учитывать, что иногда на первый взгляд болезнь обусловлена действием различных генов, но нужно помнить, что существует плейотропия, когда один ген может вызвать разные болезни.
1
Консультирование может быть ретроспективным, когда уже событие рождения больного ребенка произошло, и родители хотят знать вероятность рождения следующего нормального ребенка. Проспективное консультирование – ребенок еще не родился, хотят узнать вероятность рождения нормального ребенка.
Сбор семейных данных может быть ограничен, так как многие не знают причин смерти своих родственников, нет семейного регистра, может быть скрываемое усыновление. Часто консультируемые обладают низким интеллектом, и очень сложно внушить, что определенному пациенту нельзя иметь детей. Кроме того, часто сознательно искажаются факты. Нужен большой такт, потому что моральноэтическая сторона проблемы очень велика, и метод анкетирования на практике дает больше сведений, чем непосредственная беседа с пациентом.
При составлении родословных часто используется история болезни, протоколы о патологоанатомических вскрытиях, справки из лечебных учреждений. Обычно опрос начинают с родственников матери – бабушка, дед, их дети, данные о выкидышах, мертворождениях или у какой-либо пары отсутствие детей.
Сама анкета состоит из следующих пунктов:
ФИО, для женщин девичья фамилия
Год рождения
В каком возрасте умер
С возрастом не появились ли такие болезни как подагра, диабет и т д.? какие либо заболевания, связанные с понижением интеллекта?
Когда пошел в школу/армию/универ?
Национальность (так как, к примеру, серповидная клеточная анемия распространена у армян)
Место жительства (так как есть места эндемичные по развитию зоба), прерванное место жительства
Кровнородственные браки
Профессия, вредность профессии
Наличие хронических заболеваний
После установления диагноза врач определяет прогноз для потомства, при этом используются особые таблицы эмпирического риска, по которым врач рассчитывает этот риск.
Передача наследственных заболеваний возможна несколькими путями. Например, если у ребенка имеется заболевание как у одного из родителей, то это указывает на доминантный тип, если оба родителя здоровы – на рецессивный. В настоящее время у человека четко описаны 789 рецессивных заболеваний и 944 доминантных. Патология может быть сцеплена с полом, причем вероятность для мальчиков 50%. Возможно, это мультфакториальное заболевание – с одной стороны у этого человека комплекс генов, которые обуславливают предрасположенность к возникновению заболевания, но еще должны быть особые факторы окружающей среды. В этом случае для расчета ген риска созданы таблицы эмпирического риска. Риск до 5% считается низким и нет противопоказаний к рождению ребенка. При риске 6-10% (средний) требуется всестороннее обследование, выше 20% (высокий риск) рождение детей не рекомендуется. Однако нужно помнить, что все зависит от того, какая это патология, если, предположим, что у определенного человека шестипалость, то опасность невысока, но если это тяжелая патология – наоборот.
При хромосомных болезных вероятность повторного рождения больного довольно низка, не превышает 1%. Например, при транслокационной форме болезни Дауна важно определить, кто из родителей имеет сбалансированную транслокацию. При этом вычислено, например, если 21 хромосома «сидит» на 14, риск равен 10% (если носителем является мать, 45 хромосом у матери), если же отец, то 2,5%. Потому что существует селекция сперматозоидов, и если в неком сперматозоиде дисбаланс, то движение его ограничено, вероятность аномалии резко снижается. При транслокации 21 хромосомы на другую 21 хромосому, риск рождения составит 100% независимо от того, кто родитель.
Для определения риска повторного рождения ребенка с патологией при генных заболеваниях важно установить, кто из родителей гетерозиготный носитель патологии. Особенно это относится к аутосомнорецессивным заболеваниям и при наследовании, сцепленном с полом. В настоящее время некоторые ненаследственные заболевания устанавливаются с помощь ДНК диагностики. Заключение МГК и советы родителям часто объединены вместе. Врач-генетик дает заключение о диагнозе, вероятности возникновения болезни у будущих детей, дает рекомендации. При этом учитывается не только вероятность рождения, но и тяжесть болезни и дает также рекомендации о возможности пренатальной
2
(дородовой) диагностики, и если у родителей появляется аномальный плод, рекомендуют его абортировать.
СОВРЕМЕННЫЕ МЕТОДЫ ПРЕНАТАЛЬНОЙ ДИАГНОСТИКИ.
Эффективность МГК значительно возрастает, если использовать возможности пренатальной диагностики. Она позволяет задолго до рождения ребенка определить заболевание, и при необходимости прервать беременность. Особенно это показано в тех случаях, когда лечения нет. Основными показаниями пренатальной диагностики служат:
1.Наличие в семье точно установленного наследственного заболевания
2.Возраст матери от 35 лет, отца от 40 лет
3.Гетерозиготность родителей по рецессивному заболеванию
4.Наличие у одного из родителей транслокации, инверсии
5.Наличие в анамнезе беременной работы на вредном производстве или проживание на
радиационно загрязненных территориях Принято законодательство о рентгенологическом исследовании женщин лишь в течение 10 дней после менструаций
Косновным методам пренатальной диагностики относятся
1.Определение α-фетопротеина.
Проводят в крови и амниотической жидкости беременной. α-фетопротеин – это белок, который вырабатывается клетками печени плода. Оптимальный срок его определения 15-16 неделя беременности. Уровень этого белка резко повышается при таких заболеваниях как спинномозговая грыжа, врожденный нефроз, дефект нервной трубки, дефект брюшной стенки. Следует отметить, что уровень этого белка может повыситься и при других заболеваниях, например, опухоль печени, гепатит, цирроз, но если это синдром Дауна или Эдвардса, уровень этого α-фетопротеина, наоборот, резко снижен. Обычно при этом назначают цитогенетическое исследование клеток плода.
2. УЗИ Обычно его назначают, чтобы посмотреть врожденные пороки развития. Оптимальные сроки 17-23
неделя беременности. При некоторых патологиях, например, задержка роста плода, назначают и на более ранний срок. При этом методе исследования можно посмотреть строение головы, туловища, конечностей, половых органов, определить, мальчик это или девочка. В некоторых странах данную процедуру проводят всем беременным, что предупреждает появление детей с врожденными уродствами
3. Биопсия ворсин хореона и плаценты Это перспективный метод, проводится на сроках 7-9 недель с помощью особого шприца с гибким
катетером, при этом как бы отщепляют ворсинки хореона и далее в них проводят цитогенетический анализ хромосом, биохимические и молекулярно-генетические исследования. Если имеется патология, то беременность прерывают, но относительно высока вероятность абортирования, выкидышей плода при этом методе, поэтому используется очень редко, примерно в 3% случаев. Имеются данные, что данный метод на ранних сроках может вызвать ампутацию конечностей, поэтому ее не применяют ранее 8 недель.
4. Аммниоцентез Через брюшину матери прокалывают околоплодную оболочку и забирают 8-10 мл околоплодной
жидкости. В ней имеются слущенные клетки плода, которые можно подвергнуть цитогенетическому и биохимическому анализу. Проводят на 15-18 неделе. Риск 0,2%. Проводят под контролем УЗИ. С помощью этого метода диагностируют многие хромосомные нарушения, болезни обмена веществ.
5. Кордоцентез.
При этом методе берут образцы крови из пуповины плода и там смотрят цитогенетические, биохим и молекулярно-генетические патологии. Проводят на срок 18-22 недели. При этом хорошо диагностируются гемаглобинопатии, энзимопатии, иммунодефициты и др., а также синдром ломкости Х- хромосомы
6. Фетоскопия Данный метод основан на прямом рассматривании плода через фетоскоп, тонкий эластичный зонд с
оптической системой, этот зонд вводят через брюшину в плоскость амниона, проводят на 18-23 неделе беременности. Этот метод используется редко, только при особых показаниях, так как может вызвать осложнения: выкидыши в 8%.
ГЕНЕТИЧЕСКИЕ ЗОНДЫ.
3
Генетические зонды – это небольшие отрезки одноцепочечной молекулы ДНК, которые по своей последовательности нуклеотидов повторяют определенные участки генов у человека. Чаще всего это область промотора у гена. Такие зонды получают несколькими способами:
1.[предложил индийский ученый Хар Гобинд Корана] последовательно друг за другом соединяют особую последовательность нуклеотидов.
2.известны ферменты рестриктазы, которые могут разрезать ДНК в строго определенных районах, и эти районы можно выделить
3.используют явление обратной транскрипции, когда фермент ревертаза на основе иРНК строит молекулу ДНК. Если удается выделить такую РНК, например, в эритроцитах крови (только с белками во главе), то можно построить на этой иРНК молекулу ДНК.
4.определить последовательность нуклеотидов ДНК возможно, изучив последовательность аминокислот в определенном белке и на это основе построить участок, кусочек ДНК Как идет определение наличия или отсутствия определенного гена у человека, например, есть ли
такой патологический ген у данного человека? У данного человека надо взять материал (кровь или же соскоб слизистой оболочки рта). Далее из полученных клеток выделяют ДНК. Следующим этапом полученную ДНК надо сделать одноцепочечной, для чего эту ДНК нагревают при температуре 95-100°С. Двуцепочечная молекула ДНК распадается на две одноцепочечные нити. После этого в смесь добавляют зонд на определенный ген. Если этот зонд отыщет комплементарный участок, то соединится с ним, и образуется маленький двуцепочечный участок, эта операция идет на особых фильтрах. Следующим этапом ДНК вместе с фильтром помещают в специальную ферментную смесь нуклеаз. Эти нуклеазы разрушают все одноцепочечные нити ДНК, а двуцепочечный фрагмент разрушить не могут, и если такой двуцепочечный фрагмент существует, значит, этот патологический ген присутствует у человека. Чтобы точно определить, к зонду прибавляют радиоактивную или флуоресцентную метку. Если зонд не найдет свой комплементарный участок, то на фильтре ничего не будет (не будет светиться), зонд саморазрушится. Это самый современный метод по определению у человека наличия определенных генов.
4
Лекция 16. ЭКОЛОГИЯ ЧЕЛОВЕКА.
Экология – наука, изучающая взаимоотношения между организмом и средой. Термин (гр. Oikos – дом, logos – наука) предложил немецкий ученый Геккель. Это сложная комплексная дисциплина, здесь задействованы различные области знаний: биология, химия, физика и даже социология. Основным объектом изучения являются экосистемы. Экосистема – это комплекс живых организмов и среды обитания.
Экология разделяется на несколько направлений:
1.Аутэкология – изучает взаимоотношения отдельного организма и среды
2.Демоэкология – взаимоотношения популяции и среды
3.Синэкология – взаимоотношения экосистем и среды
В экологии человека отдельно выделяется социальная экология.
Главной задачей экологии является изучение механизмов выживания, адаптации и устойчивости экосистем.
Основные прикладные задачи:
1.Прогнозирование последствий влияния человека на природу
2.Сохранение воспроизводства и рациональное использование природных ресурсов Так же как и в биологии имеются несколько уровней: молекулярный, клеточный, органный,
организменный, популяционный, биогеоценотический, биосферный.
На Земле существуют 4 среды обитания: водная, наземно-воздушная, почвенная и живого организма (внутри).
Экологические факторы подразделяются на три основные группы:
абиотические (температура, солнечный свет, климат, влажность),
биотические факторы (влияние живых организмов друг на друга)
антропогенные факторы (влияние человека на природу).
Популяция – это минимальная самовоспроизводящаяся группа особей одного вида на
определенной территории. Биоценоз – это совокупность популяций в данном регионе или ареале. В биоценозах идут процессы борьбы за существование и естественного отбора, в биоценоз могут входить виды массовые, многочисленные, малочисленные и редкие виды.
Типы экологических взаимодействий
1.Нейтрализм, организмы не влияют друг на друга
2.Антибиоз, виды отрицательно влияют друг на друга
3.Комменсализм – взаимоотношения, приносящие выгоду
4.Аменсализм – одному виду это взаимоотношение положительно, а для другого отрицательно
5.Симбиоз – оба партнера получают выгоду
6.Хищничество – один вид поедает другой
7.Паразитизм – один вид живет за счет питательных веществ другого. Паразитизм характеризуется следующими особенностями:
чаще всего паразит нападает только на одну особь (но есть исключения)
паразит поедает только часть питательных веществ, редко приводя к гибели хозяина
паразит меньше хозяина
истинный паразитизм характеризуется узкой специализацией, что резко увеличивает выживаемость паразита
существует несколько форм паразитизма:
-временные паразиты (кратковременно нападают на человека): слепни, комары, блохи, клопы,
-постоянные паразиты (простейшие – плазмодий малярийный, круглые черви, вши)
обычно с гибелью хозяина погибает и паразит
Вэкосистему входят организмы, отличающиеся по способу питания
1.Автотрофы – сами производят органическое вещество из неорганического (фотосинтез, хемосинтез)
2.Гетеротрофы – потребляют готовое органическое вещество, они подразделяются на:
консументы, потребляющие органическое вещество живого организма (могут быть растительноядными и плотоядными, а также паразитами или симбионтами),
детритофаги или сапрофаги, питающиеся органическим веществом мертвого организма (гнилостные бактерии или жуки-копрофаги),
1
редуценты, которые завершают деструкцию органического вещества до полной минерализации (бактерии и низшие грибы).
Экологическая сукцессия – изменение состояния биогеоценозов. Различают
1.Сингенез – сукцессия, протекающая вследствие размножения растений и конкуренции между ними без изменения места обитания
2.Эндоэкогенез – изменения в результате нарушения условий обитания
Сукцессии происходят путем замещения одного вида другим видом.
Термин «биосфера» был предложен австрийским ученым Зюссом, но развил это понятие Вернадский. Он считал, что живые организмы изменяют облик Земли, например, зеленые растения, используя энергию Солнца вовлекают неорганические вещества Земли в непрерывный круговорот и, второе, он развил представление о роли человека в эволюции Земли. Ввел понятие «ноосфера» – сфера разума.
Вернадский ввел несколько биогеохимических принципов:
1.Миграция химических атомов всегда стремится к максимальному своему проявлению, эта закономерность проявляется в способности живого к распространению
2.Эволюция видов приводит к появлению устойчивых форм жизни и усиливает миграцию атомов
3.Живое вещество непрерывно создается энергией Солнца, поэтому этот процесс имеет космическое значение
ЭКОТОКСИКОЛОГИЯ Экотоксикология изучает загрязнение условий окружающей среды человека. Количество
загрязняющих веществ огромно, в настоящее время известно около 9 млн химических веществ и соединений, 600 тыс. – это промышленные яды, ядохимикаты, отравляющие вещества, бытовая химия и лекарства.
Химические вещества-загрязнители называются поллютанты. По степени опасности выделяют 4
класса:
1.Чрезвычайно опасные вещества. Предельно допустимая концентрация – менее 0,1 мг/м3
2.Высокоопасные – до 1 мг
3.Умеренные – до 10 мг
4.Малоопасные – более 10 мг
Наибольшую опасность представляют тяжелые металлы, среди них особенно опасны свинец, кадмий и ртуть.
Свинец интенсивно рассеивается, потому что входит в состав этилированного бензина. При сгорании 1 л бензина выделяется 0,5 г. свинца. Каждый год рассеивается более 2 млн тонн. Особенно большая концентрация наблюдается около автострад, кроме того, свинец выделяется при сжигании каменного угла. Известны свинцовые отравления, при которых поражается ЦНС, имеются генетические последствия.
Кадмий в природных химических соединениях – это рассеянный металл, его добыча невелика, в мире добывается около 13 тыс. тонн в год. Это спутник цинка. Кадмий рассеивается вместе с минеральными удобрениями и при сжигании мусора, может накапливаться в организме, поражая почки, костную ткань. Опасный канцероген, вызывающий рак.
Ртуть. Большую часть выделяют электростанции, другой источник – цементное производство. В атмосфере ртуть адсорбируется с пылевыми частицами. Вызывает поражения НС и обладает тератогенным действием.
Кроме тяжелых металлов опасность представляют пестициды или синтетические яды (это инсектициды - уничтожение насекомых, фунгициды – грибков, гербициды – сорняков). Ежегодно производится 2,5 млн тонн пестицидов, существует около 50 тыс. видов пестицидов. Многие из них обладают устойчивостью к разрушению, например, дуст ДДТ не производится уже 50 лет, однако в печени любого человека и даже пингвина можно обнаружить до 1 грамма этого вещества.
Нефть, основной компонент которой – углеводороды. Нефть, попадая в воду, образует нефтяную пленку и, попадая в водоносные горизонты, в дальнейшем вызывает загрязнение питьевой воды. На кафедре СибГМУ изучали канцерогенное действие нефтяных продуктов. Было обнаружено, что если кролику сбрить шерстку на спине и смазывать чистой нефтью ту часть кожи, то через 10 дней разовьется рак кожи. Таким образом, чистая нефть представляет большую опасность для живых существ.
2
Лекция 17. ГЕНЕТИЧЕСКАЯ ИНЖЕНЕРИЯ И ГЕНОТЕРАПИЯ.
Впервые в 1965 г. отечественный ученый Гершензон провел эксперимент по переносу генов окраски яиц у тутового шелкопряда, сделано это с помощью вируса полиэдра. Опыт оказался удачным, т.е. гены работали, окраска изменилась. Перенос генов с помощью вируса и бактериофагов впервые был сделан в 1952 г. Циндером и Геденбергом от одной бактерии к другой бактерии и это явление было названо трансдукцией, но эти ученые просто наблюдали такой перенос у бактерий, сами при этом не экспериментировали.
В50-х гг. прошлого века впервые появились инструменты в виде ферментов рестриктаз, с помощью которых возможно было вырезать определенный ген, и это резко стимулировало исследования по генетической инженерии. Американские ученые Мерилл, Ганьер и Петричани смогли с помощью бактериофага перенести ген, определяющий фермент галактозо-1- фосфатуридилтрансферазу. Из кишечной палочки был взят ген и внедрен в клетки человека в условиях вне организма. Человек болел галактоземией, у него была мутация по данному гену, гену фермента. Внедренный ген стал работать и продуцировать этот фермент, но только на протяжении 41 дня. Больные с таким дефектом не могут усваивать молочный сахар, и если это вовремя не определить, то ребенок погибнет.
В70-х гг. прошлого века начался активный период холодной войны. Ученые с той и с другой стороны стали работать над бактериологическим оружием. Группа американских ученых под руководством Коэна работали над бактерией кишечной палочки, это наш симбионт, дающий человеку витамины группы В. У нас нет иммунитета против кишечной палочки. Эта группа внедрила
вкишечную палочку гены устойчивости к антибиотикам, затем были внедрены гены агрессивности золотистого стафилококка, далее начались работы с генами чумы, холеры, оспы. Возникла опасность неконтролируемого распространения такой кишечной палочки и в 1974 г. на конгрессе молекулярных биологов было принято общее решение советских и американских генетиков наложить мораторий на исследования такого рода. Однако никто не прекратил эти работы.
Было разработано три класса опасностей:
1.работа с клетками человека и теплокровных животных и с генами вирусов и бактерий, которые могут паразитировать у теплокровных организмов. Чрезвычайные меры предосторосжности, особая система контроля, пониженное атмосферное давление, в подземных бункерах, специальные бассейны
2.работают с клетками холоднокровных животных
3.работы с растительными генами
ГЕННАЯ ТЕРАПИЯ Первые успехи в 1990 г. ученые попытались внедрить гены врожденного иммунодефицита в
клетки больного человека, у этого человека брали лимфоциты крови, в условиях вне организма размножали эти лимфоциты, а затем выделяли из здоровых клеток другого человека этот нормальный ген, присоединяли его к ретровирусу, заражали клетки больного человека. Затем эти клетки вводили, когда ген начинал работать. Успех был потрясающий. Наступали значимые улучшения. Другой случай (1999 г.) был предпринят с другим геном. 18-летний юноша страдал мутацией гена орнитинтранскарбамилазы, ему на основе аденовируса был введен нормальный ген, но здесь ему ввели, резко поднялась температура, тромбоз сосудов, юноша погиб. Данный инцидент приостановил исследования в то время.
Генная терапия предполагает введение ДНК в клетки-мишени с какой-то аномалией. В первом случае вводят просто нормальный ген и пытаются добиться излечения, а во втором случае вводят ДНК для того, чтобы уничтожить пораженную клетку, например опухолевая или пораженная ВИЧ. В генной терапии существует проблема эффективности введения генов, нужно не просто ввести ген, но и чтобы он встроился туда, куда надо и работал. Существуют физические, химические и биологические методы доставки генов в клетку. Есть два пути
1.берем клетки от больного человека, их исправляем и вводим больному человеку
2.вводим ДНК непосредственно больному человеку
Существует три основных способа доставки ДНК в клетку
1.с помощью вируса
2.физические способы: с помощью электропорации, баллистическая трансфекция
1
3. использование невириусных носителей Самый примитивный способ – это перенос «голой» ДНК в организм человека. В 1992 г.
немецким ученым Гольфманом успешно был инъецирован ген β-галактозидазы в мышцы. И было установлено, что мышечные волокна обладают эндоцитозом, такая же способность у эпителиальных клеток (другие клетки эндоцитозом не обладают). Преимущество такого способа в дешевизне.
Другой путь назвали методом гидродинамического шока. Внутривенно вводят физиологический раствор, в котором находится ДНК данного гена, причем его вводят до 10% от массы тела. При этом происходит трансфекция примерно 20% клеток. Эксперимент проводился на макаках, но 1% животных погиб.
Вирусы. Используют следующие вирусы: ретровирусы, аденовирусы, вирус герпеса, вирус СПИДа и вирус ветряной оспы.
Аденовирусы – наиболее эффективная система доставки генов, они не способны внедряться в геном клеток человека и не способны реплицироваться, можно внедрить большие отрезки ДНК (до 30 тыс. нуклеотидов), но есть и недостатки – слишком короткая экспрессия генов, а также у организма вырабатывается иммунная реакция на этот вирус, и клетки, пораженные им, уничтожаются.
Ретровирус. Часто используют лейкемии Молони мышей, который вызывает лейкемию у мышей, с его помощью в 1990 г успешно были излечены люди с иммунодефицитом. Здесь длительность экспрессии больше, т.к. этот вирус способен внедриться в ДНК, принести с собой нужный ген и остаться там. Недостаток: проникает только в активно делящиеся клетки и может нести не очень большие отрезки (всего 8 тыс. нуклеотидов).
Лентивирусы. Интегрируются в геном. Вирус способен внедриться не только в делящиеся, но и в неделящиеся клетки. Наиболее везуч в этом отношении ВИЧ. Из него делают гибрид, чтобы не инфицировать в полной мере человека. Берут оболочку вируса везикулярного стоматита, а внутри находится ДНК этого ВИЧ. Однако такая гибридизация очень сложна, что тормозит данное направление.
Вирус простого герпеса. Имеет сродство к определенным тканям человека, в основном, к нервной ткани, поэтому он может принести новые ДНК только туда, в нервные клетки. Используют для лечения опухолей НС, таких заболеваний как болезнь Паркинсона и Альцгеймера.
Невирусные способы доставки ДНК. Чаще всего применяется т.н. «генное ружье» или метод баллистической трансфекции. Получают золотые наночастицы, на них осаждают ДНК. Генным ружьем со сжатым газом, по существу, стреляют по клеткам, пробивается клеточная мембрана и ДНК оказывается в клетках.
Применяют и безигольного введения. Тоже осаждают ДНК на золотые частицы, используют физиологический раствор под большим давлением и чаще всего это делают на клетках кожи и мышцах, вводят эти частицы. Это еще называют генными вакцинациями.
Метод электропорации. С помощью электрических разрядов делают поры в мембране и вгоняют ДНК. Эти два метода применяют для лечении опухолей, кроме того используют метод микро-инъекций, вгоняя ДНК в яйцеклетку, так получают трансгенных животных, предпринимаются попытки и на человеческих яйцеклетках, но в литературе не описаны положительные случаи.
В последнее время стали применять метод использования липосом. Изготавливают липидный пузырек, в него помещают ДНК, и этот пузырек активно прикрепляется к мембране, сливается с ней, таким способом обычно лечат легочные генетические патологии, например, муковисцидоз и бронхиальную астму. Но показано, что из 1000 липосом только 1 доставляет ДНК в нужное место, и эта ДНК начинает работать. Перспективными в настоящее время являются вирусные методы, хотя они представляют значительную опасность.
2