


105
Глава 5. Транскрипция
5.1 Организация генетической информации
Подавляющее большинство живых клеток содержат в себе молекулы нуклеиновых кислот, отвечающих за хранение, передачу и реализацию наследственной информации. Чаще всего эту функцию выполняет ДНК, но такой способностью обладает
иРНК (как, например, у некоторых вирусов). Совокупность наследственного матери-
ала внутри одной клетки называют геномом. Изучением генома различных организмов занимается геномика, оформившаяся в отдельную область знаний с появлением
иразвитием методов секвенирования ДНК.
Ген — это структурно-функциональная единица наследственности у живых организмов. Он представляет собой участок ДНК, кодирующий некую молекулу РНК: матричную, транспортную, рибосомальную или иную (малые РНК). Число генов неодинаково у различных организмов и сильно варьирует в живом мире.
Однако не стоит связывать число генов со сложностью живого организма. Например, геном риса содержит 51 000 генов, геном лабораторной мыши — 30 000 генов, геном человека содержит 20 805 генов (по данным проекта Ensembl на апрель 2014 года), а геном кишечной палочки E. coli — 4 300 генов.
К настоящему времени успешно секвенировано почти 99,7% человеческого генома. Оставшиеся 0,3% предсталяют собой области центромер и теломер и содержат малое количество генов. Лишь 1,2% всего генома человека кодирует белки — 20 805 генов, ответственных за это, распределены между всеми молекулами ДНК (у человека
— 23 пары хромосом, а значит, 46 молекул ДНК в каждой клетке, имеющей ядро). Некоторые гены объединены в кластеры (есть как у про-, так и у эукариот). Примерно 75% генов в геноме человека имеют аналоги и у других видов. Лишь четверть генов присутствует только у позвоночных, и лишь четверть обнаружена как у про-, так и у эукариот. Как и ожидалось, человеческий геном содержит примерно столько же генов, ответственных за основные клеточные функции, сколько и геном других эукариот. Но он включает больше генов, специфичных именно для позвоночных животных: к примеру, генов, имеющих отношение к иммунной системе, а также к нервной и гормональной регуляции.
Значительная часть генома человека транскрибируется в другие виды РНК. Несмотря на то, что экзоны составляют лишь малую часть ДНК человека (примерно 1,4%), почти 80% нашего генома транскрибируется. Продукты транскрипции — различные виды РНК, кодируемые почти 4 тысячами генов: тРНК, рРНК и другие малые некодирующие РНК (нкРНК), функция которых до сих пор не выяснена. Некоторые нкРНК полиаденилированы, как и мРНК, но не связаны ни с одним из известных генов. Вероятно, такие нуклеотидные последовательности представляют собой «транскрипционный шум», возникающий при синтезе цепи на промоторном участке. Однако некоторые нкРНК довольно консервативны у разных видов и с одинаковой частотой обнаруживаются в определенных тканях. Это говорит о том, что подобные молекулы РНК выполняют регуляторные функции.
106 |
Глава 5 |
Транскрипция |
5.2 Общая характеристика транскрипции
Транскрипция — это синтез молекул РНК на матрице ДНК особыми ферментами — ДНК-зависимыми РНК-полимеразами (см. Рис. 72 ▼). Напомним, что ген — это последовательность ДНК, кодирующая молекулу РНК (продукт) и подвергающаяся транскрипции. РНК-полимераза синтезирует полинуклеотидную цепочку РНК из рибонуклеозидтрифосфатов, используя одну цепь ДНК в качестве матрицы (антисмысловая или некодирующая цепь). Клетки содержат несколько типов РНК: матричные РНК (мРНК), рибосомальные РНК (рРНК), транспортные РНК (тРНК) и малые РНК. Транскрипция начинается с промоторной последовательности (промо́тора), с которой связывается РНК-полимераза. Синтез цепи происходит в направлении 5’→3’. Некоторые гены экспрессируются конститутивно (т.е. постоянно), остальные регулируются с помощью белков-регуляторов (ингибиторов или активаторов) и экспрессируются индуцибельно (при действии определенного стимула).
Рис. 72. Общая схема транскрипции (на примере кишечной палочки E. coli).
5.3Гипотеза Жакоба и Моно
В1961 году Жако́б и Моно́ выдвинули свою теорию, которую позже назвали гипотезой Жакоба-Моно. Согласно их теории, геном прокариот содержит множество оперонов, структурно-функциональных единиц генома, кодирующих мРНК и, следовательно, белки. Часто опероны включают не один, а несколько структурных генов со связанными функциями. Оперон, как говорится в модели оперона Жакоба и Моно, содержит:
Строение РНК-полимераз 107
1.Ген-регулятор, кодирующий белок-регулятор (белок-репрессор), связывающийся с оператором;
2.Оператор — регуляторный участок ДНК;
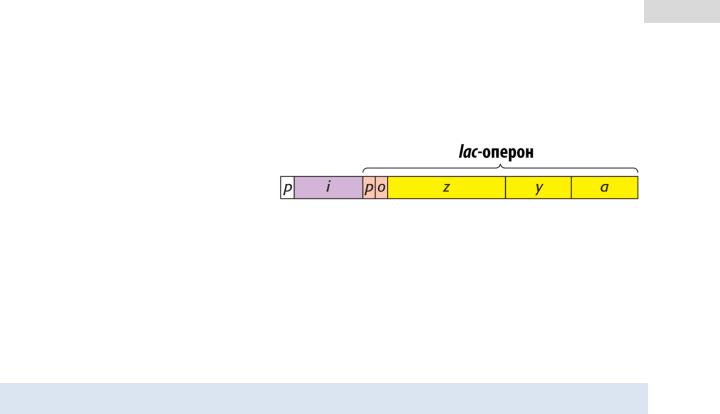
3.Структурные гены, кодирующие мРНК и, следовательно, белки. Рассмотрим строение lac-
оперона, который активно изу- |
|
|
|||
чали Жакоб и Моно. |
|
|
|||
|
В левой части рисунка по- |
|
|
||
казан |
ген-регулятор i (сирене- |
Рис. 73. |
Строение lac-оперона. |
||
вого цвета) и его промотор p (бе- |
|
|
|||
лого |
цвета) (см. |
Рис. 73 |
). |
|
|
Справа от них находится промотор p и оператор o самого lac-оперона (оранжевого цвета). За ними следуют три структурных гена z, y и a, кодирующие ферменты β -га-
лактозидазу, пермеазу и трансацетилазу, соответственно.
Регуляция работы lac-оперона рассмотрена в Теме 7.
5.4 Строение РНК-полимераз
Прокариоты. РНК-полимераза кишечной палочки E. coli состоит из 5 видов субъединиц αβ β ’ωσ и имеет массу ~449 кДа. После инициации транскрипции σ- субъединица отщепляется от кор-фермента α2β β ’ω, который и осуществляет синтез РНК на матрице ДНК.
Эукариоты. В ядре эукариот было обнаружено 3 вида РНК-полимераз, отличающихся по своей специфичности:
1.РНК-полимераза I обнаруживается в ядрышке (местах синтеза и сборки рибосом в ядре) и синтезирует большинство рРНК.
2.РНК-полимераза II находится в нуклеоплазме и синтезирует мРНК.
3.РНК-полимераза III тоже находится в нуклеоплазме и синтезирует пре-
5S-рРНК, тРНК и ряд малых ядерных и цитозольных РНК.
4.В дополнение к 3 основным видам РНК-полимераз эукариоты содержат отдельные РНК-полимеразы в митохондриях и хлоропластах.
Эукариотические РНК-полимеразы имеют молекулярную массу порядка 600 кДа, состоят из 12 субъединиц (или более). Каждая эукариотическая РНК-полимераза включает:
—Две неидентичные «большие» субъединицы (>120 кДа), гомологичные прокариотическим β и β ’-субъединицам;
—До 12 «малых» субъединиц (<50 кДа), из которых одна гомологична α- субъединице прокариот, а другая — ω-субъединице.
Степень гомологичности РНК-полимераз у людей и, к примеру, дрожжей очень высока. Настолько, что субъединица РНК-полимеразы II человека могла бы заменить аналогичную у дрожжей без потери активности.
108 |
Глава 5 |
Транскрипция |
5.5 Инициация транскрипции у прокариот
Инициация — это первая стадия транскрипции, в ходе которой происходит сборка транскрипционного комплекса, расплетение спирали ДНК и синтез короткого фрагмента РНК.
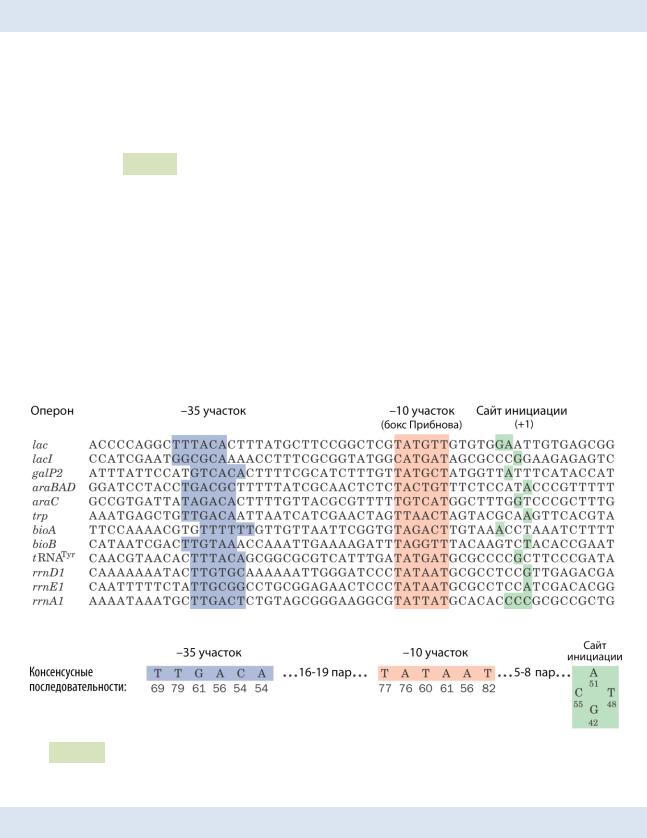
У прокариот процесс начинается с того, что прокариотический σ-фактор («сигма»-фактор или субъединица) РНК-полимеразы «узнаёт» промо́тор и связывается с ним (см. Рис. 74 ▼). Промотор — фрагмент цепи ДНК до сайта начала транскрипции, включающий два участка: –10 (бокс Прибнова) и –35. Затем к σ-фактору присоединяется кор-фермент РНК-полимеразы (комплекс субъединиц α2ββ’ω) и корректно располагается на промоторе для инициации транскрипции (σ-фактор повышает сродство РНК-полимеразы к промотору). Формируется закрытый ком-
плекс.
Далее РНК-полимераза меняет конформацию, её β’- и σ-субъединицы инициируют разделение цепей ДНК (отрезок длиной в 14 нуклеотидов), формируется тран-
скрипционный пузырёк и открытый комплекс. Затем σ-субъединица отделяется,
к РНК-полимеразе присоединяются дополнительные белки, включая NusA, и полимераза продвигается по цепи ДНК, покидая промотор.
Рис. 74. Последовательности нуклеотидов нескольких промоторов в кодирующей цепи ДНК кишечной палочки E. coli.
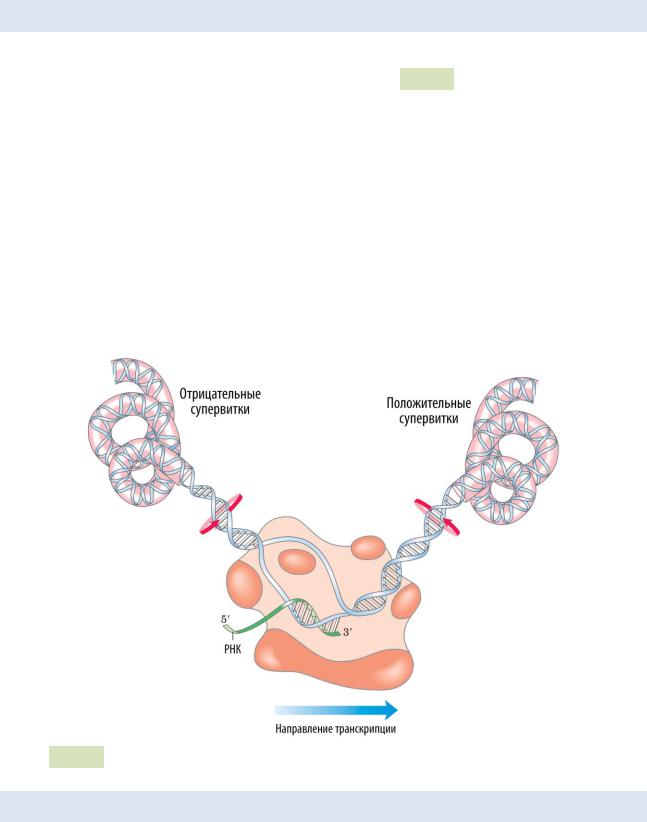
5.6 Элонгация транскрипции у прокариот
Элонгация — это вторая стадия транскрипции, в ходе которой происходит синтез цепи РНК. Он осуществляется в направлении 5’→ 3’ (подобно синтезу ДНК в репликации).