60. Биологическое значение глюконеогенеза
Биосинтез В-глюкозы — необходимый жизненно важный процесс для всех высших животных. Постоянное поступление глюкозы в качестве источника энергии необходимо для нервной системы, а также эритроцитов, почек, всех тканей эмбриона. Мозг человека потребляет более 120 г глюкозы в сутки. При понижении концентрации глюкозы в крови ниже критического уровня происходит нарушение функционирования мозга; тяжелая гипогликемия приводит к коматозному состоянию. Очевидно, глюкоза играет существенную роль в поддержании эффективных концентраций интермедиатов цикла Кребса. Она является необходимым метаболитом липидного обмена, так как служит источником глицерола — структурного элемента глицеридов.
Биосинтез глюкозы в организме животных постоянно осуществляется благодаря совокупности строго регулируемых реакций. Предшественниками В-глюкозы являются лактат, пируват, глицерол, промежуточные продукты цикла трикарбоновых кислот и большинство аминокислот. У животных синтез глюкозы из неуглеводных предшественников называют глюконеогенезом, что означает образование "нового" сахара. Глюкоза служит предшественником других физиологически важных углеводов. В этой связи в первую очередь следует отметить биосинтез гликогена, осуществляющийся главным образом в печени и мышцах. Гликоген печени выполняет резервную функцию: глюкоза, отщепляемая от цепей гликогена, поступает в кровь. Распад мышечного гликогена обеспечивает потребности в АТР в ходе мышечного сокращения.
В растениях глюкоза, а также другие углеводы образуются в процессе фотосинтеза за счет восстановления диоксида углерода.
У позвоночных глюконеогенез протекает главным образом в печени, значительно менее интенсивно — в почках (в корковом веществе). Если центральным путем катаболизма углеводов является превращение глюкозы в пируват, то центральным путем глюконеогенеза принято считать превращение пирувата в глюкозу. В этой связи гликолиз иногда называют "путем, ведущим вниз", а глюконеогенез — "путем, ведущим вверх". Большинство реакций глюконеогенеза представляют собой обращение соответствующих реакций гликолиза. И гликолиз, и глюконеогенез представляют собой необратимые процессы.
69. Роль фосфолипидов.
К фосфолипидам относятся 1) фосфатидная кислота и фосфатидилглицеролы, 2) фосфатидилхо-лин, 3) фосфатидилэтаноламин, 4) фосфатидилино-зитол, 5) фосфатидилсерин, 6) лизофосфолипиды, 7) плазма л огены и 8) сфингомиелины.
Фосфатидная кислота и фосфатидилглицеролы
Фосфатидная кислота является важным промежуточным соединением в ходе синтеза триацилгли-церолов и фосфолипидов, но в тканях содержится в незначительных количествах.
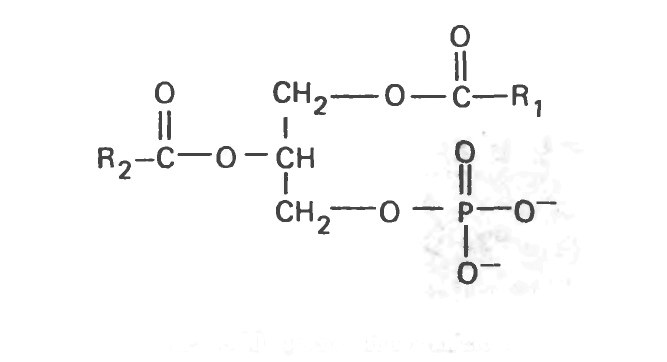
Рис. 15.10. Фосфатидная кислота.
Кардиолипнн — фосфолипид, содержащийся
в мембранах митохондрий. Он образуется из фосфа-тидилглицерола (рис. 15.11).
Фосфатидилхолин (лецитин)
Лецитины, как и простые жиры, содержат глицерол и жирные кислоты, но в их состав еще входят фосфорная кислота и холин. Лецитины широко представлены в клетках различных тканей, они выполняют как метаболические, так и структурные функции в мембранах. Дипальмитиллецитин — очень эффективный поверхностно-активный агент, снижающий поверхностное натяжение и тем самым препятствующий слипанию внутренних поверхностей дыхательных путей в легких. Его отсутствие в легких
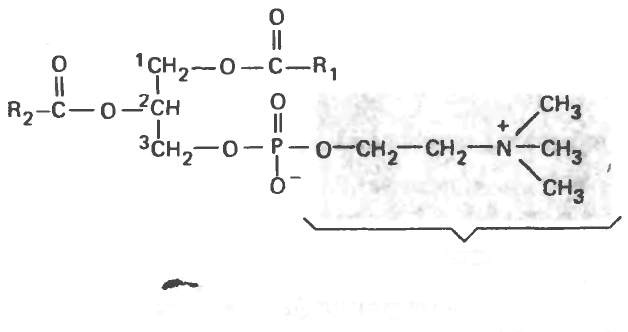
Рис. 5.12. 3-фосфатидилхолин.'
недоношенных новорожденных приводит к развитию синдрома дыхательной недостаточности. Большинство фосфолипидов содержит насыщенный аци-льный радикал в положении С, и ненасыщенный радикал в положении С, (рис. 15.12).
Фосфатидилэтаноламин (кефалин)
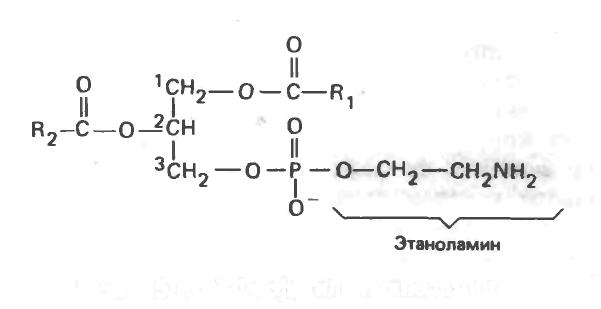
Рис. 15.13. 3-Фосфатидил этаноламин.
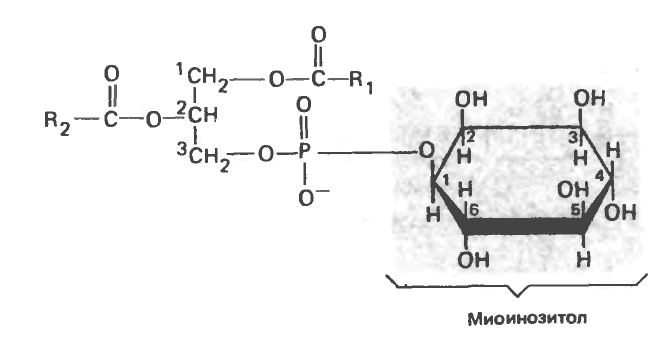
Фосфатидилинозитол
Инозитол в этом соединении представлен одним из стереоизомеров — миоинозитолом (рис. 15.14). Фосфатидилинозитол-4,5-бисфосфат является важным компонентом фосфолипидов, входящих в состав клеточных мембран; при стимуляции соответствующим гормоном он расщепляется на диацилглицерол и инозитолтрифосфат действуют как внутриклеточные, или вторые посредники.
Фосфатидилсерин
В тканях находится также родственный кефалину фосфолипид. содержащий вместо этаноламина остаток серина (рис. 15.15). Кроме того, были выделены фосфолипиды, содержащие остаток треонина.
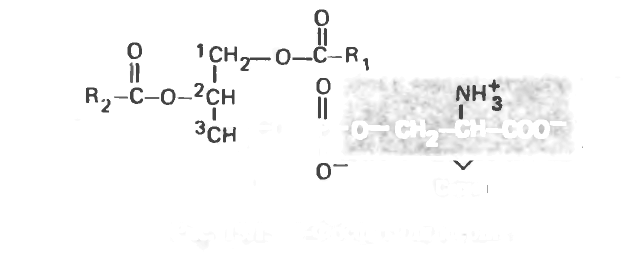
Рис. 15.15. 3-Фосфатидилсерин.
Лизофосфолипиды
Эту группу соединений образуют фосфоацилглицеролы, содержащие только один ацильный радикал. Примером служит лизолецитин, играющий важную роль в метаболизме фосфолипидов
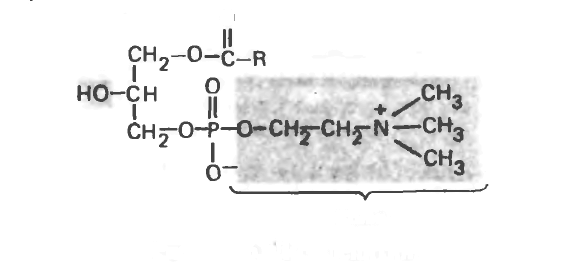
Рис. 15.16. Лизолецитин
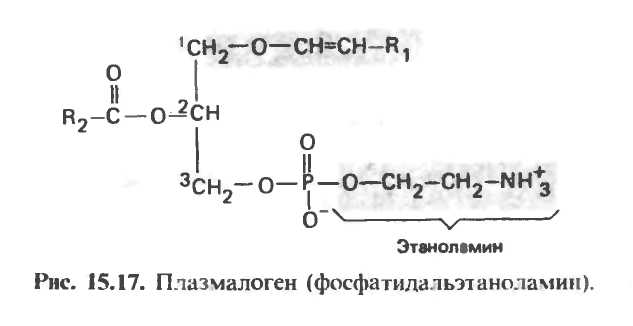
Плазмалогены
На долю этих соединений приходится до 10% фосфолипидов Мозга и мышечной ткани. В структурном отношении они родственны фосфатидилэтано-ламину, но имеют при атоме углерода С, простую эфирную связь, а не сложноэфирную, как большинство других ацилглицеролов. Алкильным радикалом в плазмалогенах обычно является ненасыщенный спирт (рис. 15.17).
В некоторых случаях этаноламин замешают холин, серии или инозитол.
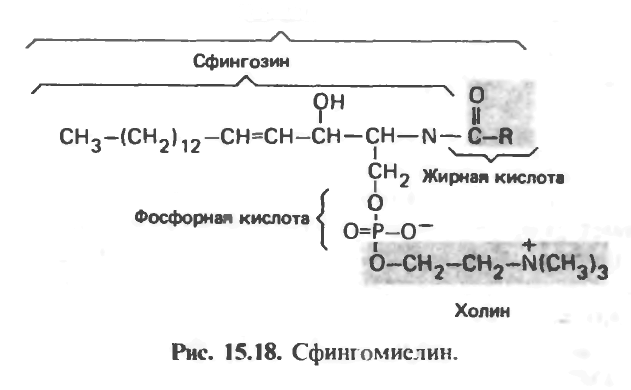
Сфингомиелины
Сфингомиелины в больших количествах встречаются в нервной ткани. При гидролизе сфингомиелинов образуются жирная кислота, фосфорная кислота, холин и сложный аминоспирт сфингозин (рис 15.18). Глицерола в составе этих соединений нет. Соединение сфингозина с жирной кислотой получило название церамид, он обнаружен в составе гликолипидов (см. ниже).