

Биохимия._Углеводы
.pdfжания гликогена в мышцах отмечается в период восстановления при приеме богатой углеводами пищи. В печени гликоген накапливается после еды.
Такие отличия печени и мышц обусловлены наличием различных изоферментов гексокиназы, фермента, который фосфорилирует глюкозу в глюкозо- 6-фосфат.
Для печени характерен изофермент, получивший собственное название – глюкокиназа. Отличия-ми этого фермента от гексокиназ других тканей являются:
•в низком сродстве к глюко-зе, что ведет к захвату глюкозы печенью только при ее высокой концентрации в крови (после еды);
•продукт реакции – глюкозо-6-фосфат – не ингибирует фермент, в то время как в других тканях гексокиназа чувствительна к такому влиянию.
Благодаря этим отличиям гепатоцит может эффективно захватывать глюкозу после еды и метаболизировать ее в любом направлении.
Например, при переполнении запасов гликогена накапливающийся глюкозо-6-фосфат не подавляет глюкокиназу и усвоение глюкозы, а просто идет на
окисление до ацетил-S-КоА и в пентозофосфатный цикл, что в целом увеличивает синтез липидов.
Регуляция глюкокиназы: активация – андрогены и инсулин, подавление – глюкокортикоиды и эстрогены.
МЕТАБОЛИЗМ ГЛИКОГЕНА
МОБИЛИЗАЦИЯ ГЛИКОГЕНА
Мобилизация (распад) гликогена или гликогенолиз в норме активируется при недостатке свободной глюкозы в клетке, а значит и в крови (голодание, мышечная работа) . При этом уровень глюкозы крови поддерживают только печень и почки, ткани в которых имеется фермент глюкозо-6-фосфатаза, гидролизующий фосфат-ный эфир глюкозы. Образуемая свободная глюкоза выходит через плазматическую мембрану в кровь. Остальные органы используют гликоген только для собственных нужд.
В гликогенолизе непосредственно участвуют три фермента:

1.Фосфорилаза гликогена – расщепляет α-1-4-гликозидные связи с образованием глюкозо-1-фосфата. Фермент работает до тех пор, пока до точки ветвления α(1-6) не останется 4 остатка глюкозы.
2.α(1-4)–α(1-6)-глюкантрансфераза, фермент переносящий трисахаридный фрагмент на другую цепь с образованием новой α-1-4-гликозидной связи. При этом
на прежнем месте остается один остаток глюкозы и «открытая» доступная α-1-6-гликозидная связь.
3.Амило-α-1-6-глюкозидаза, «деветвящий» фермент – гидролизует
α-1-6-гликозидную связь с высвобождением свободной глюкозы. В результате обра-зуется цепь без ветвлений, служащая субстратом для фосфорилазы.
АКТИВАЦИЯ ФОСФОРИЛАЗЫ
Скорость гликогенолиза лимитируется только скоростью работы фосфорилазы гликогена. Ее активность может изменяться трямя способами.
Ковалентная модификация
Происходит фосфорилирование фермента при действии гормонов на клетку через аденилатциклазный механизм. Он является так называемым каскадным регулированием:
1.Молекула гормона взаимодействуя со своим рецептором, активирует фермент
аденилатциклазу.;
2.Аденилатциклаза превращает АТФ в циклический АМФ (цАМФ) – вторичный посредник или мессенджер;
3.цАМФ аллостерически активирует фермент протеинкиназу А;
4.Протеинкиназа А фосфорилирует различные внутриклеточные белки. Одним из этих белков является киназа фосфорилазы, которая при фосфорилировании активируется;
5.Киназа фосфорилазы фосфорилирует фосфорилазу "b" гликогена, послед-
няя в результате превращается в активную фосфорилазу "а"; 6. Активная фосфорилаза " а" гликогена расщепляет α-1-4-гликозидные связи в
гликогене с образованием глюкозо-1-фосфата.

Активация ионами кальция
Второй способ регуляции заключается в активации киназы фосфорилазы не
протеинкиназой, а ионами Ca2+ и кальмодулином. Этот путь работает при инициа-ции
кальций-фосфолипидного механизма. Такой способ оправдывает себя, например, при мышечной нагрузке, когда гормональные влияния недостаточны, зато в ци-
топлазму под влиянием нервных импульсов поступают ионы Ca2+. Также некоторые гормоны влияют на углеводный обмен посредством этого механизма.
Активация с помощью АМФ
Третий способ – аллостерическая активация благодаря присоединению АМФ к молекуле фосфорилазы "b". Способ работает в любой клетке – при увеличении рас-
хода АТФ и накоплении продуктов его распада.
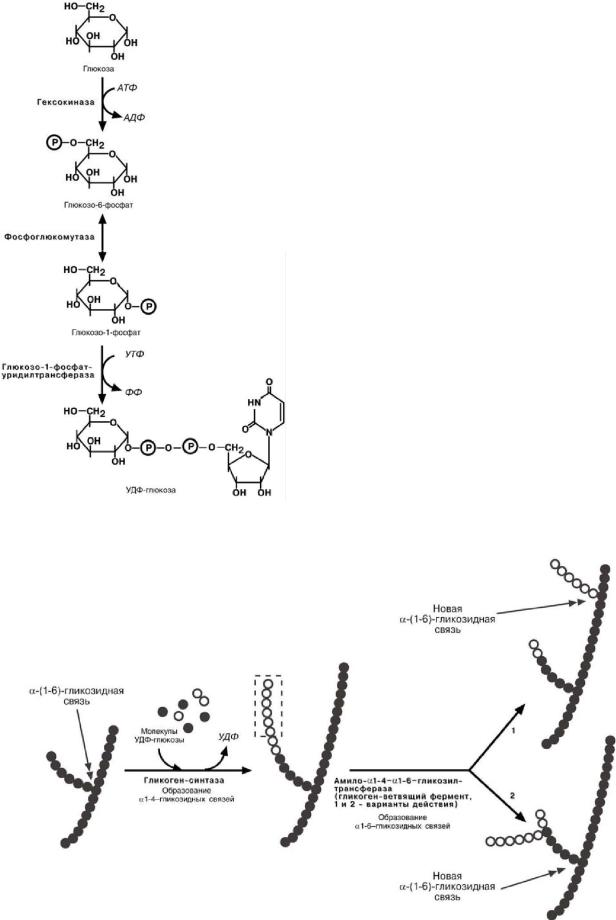
СИНТЕЗ ГЛИКОГЕНА
Синтез гликогена начинается с обра-
зования глюкозо -6 -фосфата под действи-ем глюкокиназы в печени или других гек-
сокиназ в остальных тканях. Как уже говорилось, глюкокиназа обладает низким сродством к глюкозе и в гепатоцитах глюкоза будет задерживаться только при высоких концентрациях ее в клетке .
Непосредственно же синтез гликогена осуществляют следующие ферменты:
1.Фосфоглюкомутаза – превращает глюкозо-6-фофат в глюкозо-1-фофат;
2.Глюкозо-1-фосфат-уридил-трансфе- раза – фермент, осуществляющий ключе-вую реакцию синтеза. Необратимость этой реакции обеспечивается гидролизом образующегося дифосфата;
3.Гликогенсинтаза – образует α-1-4- гликозидные связи;
4.Амило-α-1.4-α-1,6-гликозил-транс- фераза, гликоген-ветвящий фермент – переносит фрагмент с минимальной длиной в 6 остатков глюкозы на эту же или соседнюю цепь с образованием α-1-6-гликозидной связи.

РЕГУЛЯЦИЯ ОБМЕНА ГЛИКОГЕНА
Ферменты метаболизма гликогена активны либо в фосфорилированной, либо в дефосфорилированной форме:
•фосфорилаза гликогена активируется
после присоединения фосфатной группы (см выше),
•синтаза гликогена после присоединения фосфата инактиви-
руется.
Таким образом, осуществ-
ляется реципрокная (взаи-
моисключающая) регуляция:
•при работе клетки и/или гормональных влияниях активируется протеинкиназа и, как следствие, акти-
вируется фосфори-
лаза гликогена и ингибируется гликогенсинтаза. Идут реак-ции катаболизма глюкозы, и образуется энергия.
•при отдыхе или в по-
кое работают протеин-фосфатазы, которые освобождают ферменты от фосфорной кислоты: в результате гликоген-фосфорилаза дефосфорилируется и
становится не активной, активируется гликоген-синтаза. Начинается за-пасание глюкозы в виде гликогена.

ГЛИКОГЕНОВЫЕ БОЛЕЗНИ
Это наследственные заболевания, обусловленные недостаточностью каких-либо ферментов, отвечающих за метаболизм гликогена. Средняя
частота встречаемости составляет 1:40000.
Гликогенозы
Ранее гликогенозы классифицировались по номерам, однако в связи с открытием новых видов этих болезней появилось много разночтений. В настоящее время глико-
генозы делят по патогенетическому признаку на печеночные, мышечные и сме-шанные формы. Следует отметить, что при гликогенозах количество гликогена не всегда изменено, изменения могут быть только в структуре его молекулы.
Печеночные гликогенозы
Самый частый гликогеноз I типа или болезнь фон Гирке обусловлен аутосом- но-рецессивным дефектом глюкозо-6-фосфатазы. Из-за того, что этот фермент есть только в печени и почках, преимущественно страдают эти органы, и болезнь носит еще одно название – гепаторенальный гликогеноз. Даже у новорожденных детей
наблюдаются гепатомегалия и нефромегалия, обусловленные накоплением гликоге-на не только в цитоплазме, но и в ядрах клеток. Кроме этого, активируется синтез липидов с возникновением стеатоза печени. Так как фермент необходим для де-фосфорилирования глюкозо-6-фосфата с последующим выходом глюкозы в кровь, у больных отмечается гипогликемия, и, как следствие, ацетонемия, метаболический ацидоз, ацетонурия.
Гликогеноз III типа или болезнь Форбса-Кори или лимит-декстриноз – это аутосомнорецессивный дефект амило-α-1-6-глюкозидазы, «деветвящего» фермен-та, гидролизующего α-1-6 -гликозидную связь. Болезнь имеет более доброкачествен-ное течение и частота ее составляет примерно 25% от всех гликогенозов. Для боль-ных характерна гепатомегалия, умеренная задержка физического развития, в подро-стковом возрасте возможна небольшая миопатия.
Еще два печеночных гликогеноза – гликогеноз IV типа (болезнь Андерсена),
связанный с дефектом ветвящего фермента и гликогеноз VI типа (болезнь Херса), связанный с дефицитом печеночной фосфорилазы гликогена встречаются довольно редко.

Мышечные гликогенозы
Для этой группы гликогенозов характерны изменения ферментов мышечной ткани. Это приводит к нарушению энергообеспечения мышц при физической нагрузке, болям в мышцах, судорогам.
Гликогеноз V типа (болезнь Мак-Ардля) – отсутствие мышечной фосфорила-
зы. При тяжелой мышечной нагрузке возникают судороги, миоглобинурия, хотя лег-кая работа не вызывает каких-либо проблем.
Смешанные гликогенозы
Эти заболевания касаются и печени, и мышц, и других органов.
Гликогеноз II типа (болезнь Помпе) – поражаются все гликогенсодержащие клетки из-за отсутствия лизосомальной α-1-4-глюкозидазы. Происходит накопле-
ние гликогена в лизосомах и в цитоплазме. Заболевание составляет почти 10% всех гликогенозов и является наиболее злокачественным. Больные умирают в грудном возрасте из-за кардиомегалии и .тяжелой сердечной недостаточности.
Агликогенозы
Агликогенозы – состояния, связанные с отсутствием гликогена.
В качестве примера агликогеноза можно привести наследственный аутосомнорецессивный дефицит гликоген-синтазы. Симптомами является резкая гипогликемия натощак, особенно утром, появляется рвота, судороги, потеря сознания. В результа-те гипогликемии наблюдается задержка психомоторного развития, умственная от-сталость. Болезнь несмертельна при адекватном лечении (частое кормление), хотя и опасна.
ГЛИКОЛИЗ
Путь, по которому глюкоза окисляется до пировиноградной кислоты для получения энергии, называется гликолизом. В зависимости от дальнейшей судьбы пирува-
та различают аэробный и анаэробный гликолиз.
Ваэробном процессе пировиноградная кислота превращается в ацетил-S-КоА и
далее сгорает в реакциях тканевого дыхания до СОB2B и НB2BО.
Ванаэробном процессе пировиноградная кислота восстанавливается до молоч-ной кислоты (лактата), поэтому в микробиологии анаэробный гликолиз называют молочнокислым брожением. Лактат является метаболическим тупиком и далее ни во что не превращается, единственная возмож-ность утилизовать лактат – это окислить его обратно в пируват.
Практически все клетки организма способ-ны к анаэробному гликолизу. Для эритроцитов он является единственным источником энер-
гии. Клетки скелетной мускулатуры за счет бескислородного расщепления глюкозы способны выполнять мощную, быструю, интенсив-
ную работу, как, например, бег на короткие дистанции, напряжение в силовых видах спорта.
Анаэробный гликолиз локализуется в цито-золе
ивключает 2 этапа из 11 ферментатив-ных
реакций.

Первый этап – подготовительный, здесь происходит затрата энергии АТФ, активация глюкозы и образование из нее триозофосфатов.
Первая реакция гликолиза, строго говоря, к гликолизу не относится. Это гексокиназная реакция о которой ранее уже говорилось ("Метаболизм гликогена"). Ее роль сводится к превращению глюкозы в реакционно способное соединение за счет фосфорилирования 6-го, не включенного в кольцо, атома углерода.
Для печени характерен изофермент гексокиназы – глюкокиназа. Низкое сродство этого фермента к глюкозе обеспечивает ее захват печенью только после приема пищи, когда создается высокая концентрация глюкозы в крови. При обычных концентрациях глюкозы в крови пе-
чень ее не потребляет и та достается другим тканям. Вторая реакция – реакция изомеризации – необхо-
дима для выведения еще одного атома углерода из кольца для его последующего фосфорилирования. В ней образуется фруктозо-6-фосфат.
Третья реакция – фосфорилирование фруктозо-6- фосфата с образованием почти симметричной молекулы фруктозо-1,6-дифосфата.
В четвертой реакции фруктозо1,6-дифосфат разрезается пополам с образование двух фосфорилирован-ных триоз-изомеров, альдозы глицеральдегида (ГАФ) и кетозы диоксиацетона (ДАФ).
Пятая реакция подготовительного этапа – переход глицеральдегидфосфата и диоксиацетонфосфата друг в друга. Равновесие реакции сдвинуто в пользу диоксиацетонфосфата, его доля составляет 97%, доля глицеральдегидфосфата – 3%. Эта реакция, при всей ее простоте, является вершителем судьбы глюкозы:
•при нехватке энергии в клетке и активации окисления глюкозы диоксиацетонфосфат превращается в глицеральдегидфосфат, который далее окисляется на втором этапе гликолиза;
•при достаточном количестве АТФ, наоборот, глицеральдегидфосфат изомеризуется в диоксиацетонфосфат, и последний
отправляется на синтез жиров (см "Синтез триацилглицеридов").

Второй этап гликолиза – это освобождение энергии, содержащейся в глицеральдегидфосфате, и запасание ее в форме АТФ.
Шестая реакция гликолиза – окисление глицеральдегидфосфата и присоединение к нему фосфорной кислоты приводит к образованию макроэргического соединения 1.3-дифосфоглицериновой кислоты.
В седьмой реакции энергия фосфоэфирной связи, заключенная в 1,3-дифосфоглицерате тратится на образование АТФ. Реакция получила до-
полнительное название –реакция субстратного фосфорилирования, что уточняет источник энергии для получения макроэргической связи в АТФ (суб-
страт) в отличие от окислительного фосфорилиро-вания (электрохимический градиент ионов водорода на мембране митохондрий).
Подобных реакций в клетке всего три – 1) ука-занная реакция, 2) пируваткиназная реакция, деся-тая реакция гликолиза (см ниже), 3) тиокиназная реакция цикла трикарбоновых кислот.
Восьмая реакция – синтезированный в предыдущей реакции 3-фосфоглицерат изомеризуется в 2-фосфоглицерат.
Девятая реакция – отрыв молекулы воды от 2-фосфоглицериновой кислоты приводит к образованию еще одной макроэргической фосфоэфирной связи.
Еще одна реакция субстратного фосфорилирования – десятая реакция гликолиза – заключа-
ется в переносе макроэргического фосфата с фосфоенолпирувата на АДФ.
Последняя реакция , одиннадцатая, образова-ние молочной кислоты из пирувата под действием лактатдегидрогеназы. Важно то, что эта реакция осуществляется только в анаэробных условиях. Эта реакция необходима клетке, так как НАДН, образующийся в 6-й реакции, в отсутствие кислорода не может поступать и окисляться в митохондриях.
При наличии кислорода пировиноградная кислота превращается в ацетил-S-КоА.
ГЛИКОЛИТИЧЕСКАЯ ОКСИРЕДУКЦИЯ
Процесс циклического восстановления и окисления НАД в реакциях анаэробного гликолиза полу-
чил название гликолитическая оксиредукция.
В анаэробных условиях образуемый в шестой, ГАФдегидрогеназной реакции, НАДН используется
в последней реакции для восстановления пирувата до лактата. Образуемый таким образом НАД опять возвращается в шестую реакцию.

В аэробных условиях НАДН отдает свои атомы водорода на челночные системы (см ниже) для их передачи в дыхательную цепь митохондрий.
ЭНЕРГЕТИЧЕСКИЙ ЭФФЕКТ ОКИСЛЕНИЯ ГЛЮКОЗЫ
На подготовительном этапе на активацию глюкозы затрачивается 2 молекулы АТФ, каждая из которых оказывается на триозе – глицеральдегид-
фосфате и диоксиацетонфос-фате. В следующий второй этап входит только глицераль-
дегидфосфат, но его уже две молекулы, каждая из которых окисляется до пирувата с об-
разованием 2-х молекул АТФ
вреакциях субстратного фосфорилирования. Таким образом, суммируя, получаем, что на пути от глюкозы до пирувата
вчистом виде образуется 2 молекулы АТФ.
Однако надо иметь в виду и глицеральдегидфосфат-
дегидрогеназную реакцию, из которой выходит НАДН. Если условия анаэробные, то он используется в лактатдегидрогеназной реакции – окисляется для образования лактата и в получении АТФ не участвует.
Если же имеется кислород
– НАДН направляется в митохондрию, на процессы окислительного фосфорилирования, и там его окисление приносит
дивиденды в форме АТФ.
ЭФФЕКТ ПАСТЕРА
Эффект Пастера – это снижение потребления глюкозы и прекращение продукции молочной кислоты клеткой в присутствии кислорода.
Луи Пастер, занимавшийся вопросами виноделия, наблюдал подобный фено-мен при производстве вина. Забегая вперед, отметим, что спиртовое броже-ние весьма похоже на гликолиз, только вместо молочной кислоты из пирува-та образуется спирт.