

4 Лекция по БТ КИНЕТИКА
.docxКИНЕТИКА ПРОЦЕССОВ ФЕРМЕНТАЦИИ
ОСНОВНЫЕ МОДЕЛИ И КИНЕТИЧЕСКИЕ УРАВНЕНИЯ
ДИНАМИКА ИЗМЕНЕНИЯ ПАРАМЕТРОВ ПРОЦЕССА ФЕРМЕНТАЦИИ

КАК ОПИСЫВАТЬ СКОРОСТИ ПРОТЕКАЮЩИХ ПРОЦЕССОВ?
-
Можно найти уравнения, описывающие изменения во времени X(t), S(t), P(t) или Qx(t), Qs(t), Qp(t) или удельных скоростей μ(t), qs (t) и qp(t)
-
Эти уравнения, однако, ничего не дают для описания закономерностей процесса. Нужны «автономные» уравнения, описывающие зависимости между кинетическими характеристиками (см. выше) и текущими концентрациями S, X, P и другими параметрами процесса
КИНЕТИЧЕСКИЕ ЗАВИСИМОСТИ
-
Мы уже говорили о том, что кинетические характеристики процесса ферментации –μ, qp ,qs – не являются постоянными, а изменяются в ходе процесса
-
Эти изменения связаны с тем, что существуют зависимости этих параметров от текущих концентраций субстратов (S), биомассы (X) и продуктов метаболизма (P)
ЗАВИСИМОСТЬ СКОРОСТИ РОСТА ОТ КОНЦЕНТРАЦИИ СУБСТРАТА
-
Мы уже рассматривали одно уравнение для скорости роста биомассы
-
QX = dX/dt = μX
-
В этом уравнении μ постоянна
-
На самом деле величина μ зависит по меньшей мере от концентрации субстрата S, которая в ходе процесса непрерывно уменьшается
-
Ниже рассмотрены варианты этой зависимости
МОДЕЛЬ КОБОЗЕВА
-
По аналогии с химической кинетикой можно считать, что в реакции участвуют два «реагента» - биомасса Х и субстрат S
-
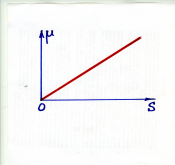
dX/dt = KXS (K – константа скорости)
-
Или μ = KS (прямая на графике)
-
Модель предсказывает неограниченное увеличение скорости роста при увеличении концентрации субстрата
МОДЕЛЬ БЛЭКМАНА
-
Практически известно, что скорость роста микроорганизмов не может быть сколь угодно большой, и повышение концентрации субстрата выше определённого предела не приводит к увеличению скорости роста биомассы
-
Блэкман предположил, что при достижении некоторого критического значения концентрации субстрата скорость роста становится постоянной по величине
ЗАВИСИМОСТЬ ПО БЛЭКМАНУ

УРАВНЕНИЯ БЛЭКМАНА
-
dX/dt = KSX при S < S*
-
dX/dt = KS*X при S > S*
-
или, что то же самое:
-
μ = KS при S < S*
-
μ = KS* при S > S*
-
В этих уравнениях K – константа скорости Кобозева, S* - критическая концентрация субстрата
МОДЕЛЬ МОНО
-
В природе не бывает резких изломов зависимостей, как в модели Блэкмана
-
Обычно удельная скорость роста с повышением концентрации субстрата постепенно возрастает, а затем плавно переходит в постоянное значение
-
Моно предложил описать эту зависимость специальным уравнением, основанным на ферментативной кинетике
МОДЕЛЬ МОНО





0 0
S
УРАВНЕНИЕ МОНО
-
dX / dt = μmXS / (Ks + S), или
-
μ = μmS / (Ks + S),
-
В этих уравнениях
-
S – концентрация субстрата
-
μm – константа максимальной удельной скорости роста
-
Ks - субстратная константа Моно
РЕПЕРНЫЕ ТОЧКИ
-
Характерные точки для графика (S) легко найти простой подстановкой в уравнение Моно :
-
= 0 при S = 0
-
= ½m при S = Ks
-
= m при S
ОПРЕДЕЛЕНИЕ КИНЕТИЧЕСКИХ КОНСТАНТ
-
Кинетические константы, входящие в уравнения модели, определяются из экспериментальных данных путём их математической обработки с помощью специальных методов
-
Некоторые из этих методов будут рассмотрены ниже на примере уравнения Моно
МЕТОД ЛАЙНУИВЕРА И БЭРКА
-
Уравнение Моно можно преобразовать так, чтобы его переменные ( μ и S ) выразить в обратных величинах ( 1/μ ) и ( 1/S ):
-
( 1/μ ) = ( 1/μm ) + ( 1/S )*(Ks / μm )
-
Это – уравнение прямой с «новыми» константами ( 1/μm ) и (Ks / μm ), которые можно найти графическим построением
ГРАФИЧЕСКОЕ ПОСТРОЕНИЕ
-
В результате эксперимента имеется некоторое количество парных данных ( 1/μ ) и ( 1/S ) для каждой точки
-
Если нанести эти данные на график в обратных координатах, получается прямая, пересекающая ось ординат в точке 1/μ = 1/μm, а ось абсцисс – в точке 1/S = - 1/KS






МЕТОД
КОРНИШ-БОУДЕНА
-
Можно преобразовать уравнение Моно к такому виду:
-
μ = μm – KS(μ/S), откуда графически получается прямая в координатах μ - (μ/S). Прямая пересекает ось ординат в точке μ = μm ,а ось абсцисс – в точке (μ/S) = (μm/KS)
-
Таким образом, по экспериментальным данным можно найти кинетические константы процесса
ГРАФИЧЕСКОЕ ПОСТРОЕНИЕ КОРНИШ-БОУДЕНА

МОДЕЛЬ МОЗЕРА
-
Для некоторых процессов зависимость скорости роста от концентрации субстрата имеет сигмоидальный характер (см. рисунок)
-
Мозером предложено описывать данную ситуацию уравнением:
-
μ = μmSk /(KS + Sk )
СИГМОИДНАЯ ЗАВИСИМОСТЬ

МОДЕЛЬ ПЕРТА
-
Эта модель описывает влияние не лимитирующего, а «стимулирующего» субстрата
-
μ = μо + μ1S/(KS + S)
-
Интересно, что при S = 0 уже есть рост, то есть μ = μо
СТИМУЛИРУЮЩИЙ СУБСТРАТ

МОДЕЛЬ АНДРЮСА
-
В этой модели учитывается ингибирование повышенными концентрациями субстрата (см. рис.)
-
Зависимость описывается уравнением
-
μ = μmS / (Ks + S + S2/KI), где KI – константа ингибирования
-
Оптимальная концентрация субстрата
-
Sopt = (KS K i)0,5
ЗАВИСИМОСТЬ С ЭКСТРЕМУМОМ

ПРОДУКТ-ЗАВИСИМЫЕ МОДЕЛИ
-
Во всех рассмотренных до сих пор уравнениях в качестве единственного параметра, влияющего на удельную скорость роста микроорганизмов, использовалась концентрация субстрата (т.е. это «субстрат-зависимые» модели)
-
Возможны случаи, когда не субстрат, а накапливающийся в среде продукт метаболизма влияет на скорость роста, и тогда это «продукт-зависимые модели»
МОДЕЛЬ ХИНШЕЛЬВУДА

УРАВНЕНИЕ ХИНШЕЛЬВУДА:
-
μ = μm– KP
-
P – концентрация продукта метаболизма
-
μm и K - кинетические константы
-
μ = μm при P = 0
-
μ = 0 при P = μm / K
МОДЕЛЬ ИЕРУСАЛИМСКОГО

УРАВНЕНИЕ ИЕРУСАЛИМСКОГО
-
μ = μm / (1 + P / KP ) или
-
μ = μm KP /(P + KP) , где KP – константа ингибирования, а μm – максимальная удельная скорость роста
-
При P = 0 μ = μm
-
При P >> μ = 0
-
При P = KP μ = 0,5μm
ЛИНЕАРИЗАЦИЯ УРАВНЕНИЯ ИЕРУСАЛИМСКОГО
-
(1/ μm ) + (1/ μmKP)P
-
При (1/ μ) P = 0 P = - KP
-
При P = 0 (1/ μ) = (1/ μm )
-
Графическое выражение линеаризованного уравнения представлено на следующем слайде
ГРАФИЧЕСКОЕ ВЫРАЖЕНИЕ ЛИНЕАРИЗОВАННОГО УРАВНЕНИЯ

МОДЕЛЬ БЕРГТЕРА
-
Встречаются процессы, в которых зависимость μ (P) имеет сигмоидный характер
-
Уравнение Бергтера:
-
μ = μm / (1 + P k / KP ), где
-
KP – константа ингибирования
-
k - показатель степени, как новая константа
-
Обычно k > 1

СТИМУЛИРУЮЩИЙ ПРОДУКТ МЕТАБОЛИЗМА
-
Иногда встречаются процессы, в которых продукт метаболизма не ингибирует, а стимулирует рост
-
Эту зависимость можно описать уравнением:
-
μ = μ0 + μ1P / ( KP + P )

ЧАСТИЧНО ИНГИБИРУЮЩИЙ ПРОДУКТ МЕТАБОЛИЗМА
-
Бывают ситуации, когда продукт метаболизма как бы «частично» ингибирует рост (не до нуля)
-
Эта зависимость может быть описана уравнением:
-
μ = μ0 + μ1 / (1 + P / KP )

БИОМАССА КАК ВЛИЯЮЩИЙ ФАКТОР
-
На ОБЩУЮ скорость роста влияет концентрация биомассы (как сомножитель). Влияет ли концентрация биомассы на УДЕЛЬНУЮ скорость роста ?
-
Да, но это влияние не просто «эффект тесноты», а опосредствованное влияние через концентрацию ингибирующего продукта.
-
Поэтому и зависимости при этом похожи, но вместо P в уравнения подставляют X.
ОТМИРАНИЕ (ДИССИМИЛЯЦИЯ) БИОМАССЫ
-
Все рассмотренные ранее зависимости относятся к собственно росту микроорганизмов
-
Однако в процессе роста, и особенно при его замедлении, одновременно с ростом происходит диссимиляция (отмирание) микроорганизмов. Реально измеряя биомассу, мы наблюдаем объединённый процесс роста-диссимиляции
ВАРИАНТЫ
УЧЁТА КИНЕТИКИ ОТМИРАНИЯ БИОМАССЫ
-
dX / dt = μX – X ( общее уравнение)
-
1) μ=0 нет отмирания
-
2) μ = K Герберт
-
3) μ = KX Ферхюльст
-
4) μ = KP Рамкришна
МНОГОФАКТОРНЫЕ КИНЕТИЧЕСКИЕ УРАВНЕНИЯ
-
До сих пор мы рассматривали кинетические уравнения, в которых на скорость роста влиял только один фактор – субстрат (только один), или продукт, или биомасса.
-
Но в реальности часто на процесс влияет не один, а несколько факторов. Существуют многофакторные зависимости, и прежде всего многосубстратные.
ТИПЫ МНОГОФАКТОРНЫХ КИНЕТИЧЕСКИХ УРАВНЕНИЙ
-
Мультипликативные уравнения
-
Аддитивные уравнения
-
Альтернативные уравнения
-
Уравнения с неразделяющимися эффектами факторов
-
Уравнения с однородными факторами (например, многосубстратные)
-
Уравнения со смешанными факторами
МУЛЬТИПЛИКАТИВНЫЕ КИНЕТИЧЕСКИЕ УРАВНЕНИЯ
-
Функция является произведением однофакторных зависимостей
-
= f1(S1)* f2(S2), где S1 и S2 – концентрации субстратов S1 и S2
-
Каждый фактор может иметь собственную форму зависимости, например, Моно – Моно, Моно – Перт, Моно –Андрюс, Андрюс – Андрюс и т.п.
АДДИТИВНЫЕ КИНЕТИЧЕСКИЕ УРАВНЕНИЯ
-
Многофакторная функция является суммой однофакторных
-
Такие зависимости встречаются довольно редко, чаще для двух-трёх субстратов одного назначения, например, глюкоза и лактоза [(S1) и (S2)]
-
= 1S1/(K1 + S1 ) + 2S2/(K2 + S2 ), где K1 ,K2 , 1, 2 – кинетические константы
АЛЬТЕРНАТИВНЫЕ КИНЕТИЧЕСКИЕ УРАВНЕНИЯ
-
Многофакторная зависимость подчиняется принципу кинетического минимума
-
Для каждого субстрата существует своя зависимость i(Si). Реализуется та из них при заданных < Si > , для которой i(Si) минимален:

КИНЕТИЧЕСКИЕ УРАВНЕНИЯ С НЕРАЗДЕЛЯЮЩИМИСЯ ПЕРЕМЕННЫМИ
-
Все три рассмотренных варианта многофакторных уравнений формируются из однофакторных зависимостей
-
В сложных случаях многофакторную зависимость трудно разбить на однофакторные. Например, уравнение «конкурентного торможения» вторым субстратом S2:
-
μ = μmS1 /(KS + S1 + S2 / K i )
ВЛИЯНИЕ ТЕМПЕРАТУРЫ
-
Температура оказывает влияние на константы скорости кинетических уравнений роста микроорганизмов
-
В принципе любая из констант в той или иной мере может быть подвержена влиянию температуры
-
Однако в кинетических уравнениях для роста чаще всего полагают, что температура влияет на максимальную удельную скорость роста m
ЗАКОН АРРЕНИУСА
-
По аналогии с химической кинетикой температурное влияние часто пытаются описывать законом Аррениуса:
-
m = m0 e- E/RT
-
m0 – предэкспоненциальный множитель
-
E – энергия активации
-
R – универсальная газовая постоянная
-
T – абсолютная температура
НЕДОСТАТКИ УРАВНЕНИЯ АРРЕНИУСА
-
Уравнение описывает только возрастание констант с повышением температуры
-
Реальная зависимость имеет вид кривой с экстремумом
-
При этом ниспадающая часть этой кривой круче, чем восходящая

«ДВОЙНОЙ АРРЕНИУС»
-
В клетках микроорганизмов одновременно протекают процессы синтеза и распада клеточного материала
-
При этом зависимость «объединённой» константы m от температуры отображает разность двух сопряжённых процессов, каждый из которых подчиняется закону Аррениуса со «своими» константами:
-
m = 1 e- E1/RT - 2 e- E2/RT
ЭМПИРИЧЕСКИЕ УРАВНЕНИЯ ЗАВИСИМОСТИ ОТ ТЕМПЕРАТУРЫ
-
Часто для описания m (Т) используют уравнение m = 0 + 1T + 2 T2
-
Здесь 0 , 1 и 2 - коэффициенты, определяемые из экспериментальных данных
-
Это уравнение, однако, описывает только СИММЕТРИЧНЫЕ зависимости
-
Более точно – уравнение более высоких порядков: m = 0+ 1T + 2T2 + 3T3 + 4T4
ВИД ЗАВИСИМОСТИ M(РН)

ЗАВИСИМОСТЬ M(РН)
-
Величина рН = - lg [H+]
-
[H+] = 10-pH [г-ион/литр]
-
В водных растворах: [H+] * [OH-] = 10-14
-
[OH-] = 10-(14 – pH) [г-ион/литр]
-
Влияние рН осуществляется «через» влияние либо водородных, либо гидроксильных ионов
-
Поэтому можно использовать уравнения (P), подставляя вместо продукта P либо [H+] для левой ветви кривой, либо [OH-] – для правой . Или использовать эмпирическое уравнение: m = 0+ 1pH + 2(pH)2 + 3(pH)3 + 4(pH)4
МОДЕЛИ КИНЕТИКИ БИОСИНТЕЗА ПРОДУКТА
-
QP = qP X
-
qP – удельная скорость биосинтеза продукта метаболизма
- Функция от удельной скорости роста
- Функция от концентраций субстратов и продуктов метаболизма
- Функция от возраста культуры
ОБОСНОВАНИЕ РОСТ-ЗАВИСИМЫХ МОДЕЛЕЙ БИОСИНТЕЗА
-
Предполагается, что комплексный показатель – удельная скорость роста микроорганизмов , зависящий от многих параметров, «вбирает в себя» всё влияние этих параметров, и его можно использовать как параметр-аргумент в уравнениях, описывающих кинетику биосинтеза продукта
-
На этом предположении основаны уравнения, предложенные и использованные для различных процессов биосинтеза продуктов – связанных с ростом и «вторичных» метаболитов
РОСТ-ЗАВИСИМЫЕ МОДЕЛИ
-
1 – прямая связь с ростом:
-
qP =K
-
2 – Людекинг и Пайри : qP =K + qP0
-
3 – qP =a /(b + )
-
4 – qP =a /(b - )
-
5 – [3] + qP0
-
6 – [4] + qP0

РОСТ-ЗАВИСИМЫЕ МОДЕЛИ
-
Зависимости с экстремумом
-
(7 ) qP = b -c 2
-
(8 ) qP = a +b + c2

НЕДОСТАТКИ РОСТЗАВИСИМЫХ МОДЕЛЕЙ БИОСИНТЕЗА ПРОДУКТОВ
-
«Рост-зависимые» модели неявно предполагают, что совсем неважно, каким образом формируется то или иное значение удельной скорости роста μ. Например, уменьшить μ можно, снизив концентрацию лимитирующего субстрата, или увеличив или понизив величину рН или температуры.
-
Такое предположение справедливо для процессов биосинтеза продуктов, связанных с ростом.
НЕДОСТАТКИ РОСТЗАВИСИМЫХ МОДЕЛЕЙ БИОСИНТЕЗА ПРОДУКТОВ
-
Для процессов, НЕ связанных с ростом, важно, каким именно образом достигается снижение величины μ.
-
Лимитирование концентрациями углеродного, или азотного субстрата, или кислорода, или повышение рН, или снижение температуры, давая ОДНО И ТО ЖЕ значение μ, могут давать совершенно разные скорости биосинтеза продукта метаболизма qP
МОДЕЛИ КИНЕТИКИ БИОСИНТЕЗА С ВНЕШНИМИ ПАРАМЕТРАМИ-АРГУМЕНТАМИ
-
Для таких процессов необходимо использовать уравнения, описывающие зависимости qP непосредственно от внешних факторов – концентраций субстрата S, или продукта P, и/или температуры и/или pH
ВАРИАНТЫ СУБСТРАТЗАВИСИМЫХ УРАВНЕНИЙ КИНЕТИКИ БИОСИНТЕЗА
-
qP = qmS / (KS + S) (Моно)
-
qP = qmS / (KS + S + S2 / Ki) (Андрюс)
-
qP = qm[ S1 / (K1+S1)]*[S2 /(K2+S2+S22 / Ki)] (Моно-Андрюс)
-
qP = qmS / (KS+S) (1+P/KP) (Моно-Иерусалимский)
-
qP = qmS / (KX + S) (Контуа)
КИНЕТИКА ДЕГРАДАЦИИ ПРОДУКТОВ МЕТАБОЛИЗМА
-
Часто продукты биосинтеза бывают лабильными. Они разрушаются в процессе самой ферментации.

-
В материальный баланс продукта в таких случаях следует
включать скорость деградации (инактивации) продукта.
ВАРИАНТЫ
ОПИСАНИЯ КИНЕТИКИ ДЕГРАДАЦИИ ПРОДУКТА
-
= 0 (стабильный продукт)

-
= K (не имеет смысла при P = 0)

-
= KP (кинетика 1-го порядка)
-
= KPn (кинетика n-го порядка)


-
= KPX (деградация с участием не только самого продукта, но и биомассы)
КИНЕТИКА ПОТРЕБЛЕНИЯ СУБСТРАТА
-
Чтобы «замкнуть» материальный баланс, в котором почти везде участвует концентрация субстрата, необходимо дополнить его уравнением кинетики потребления субстрата.
-
Это уравнение в общем виде может быть представлено как аддитивное, в правой части которого фигурируют затраты на собственно рост микроорганизмов (Q1), на образование продукта метаболизма (Q2) и на поддержание жизнедеятельности (Q3):
-
- dS/dt = Q1+ Q2+ Q3
-
Q1=QX /YXS ( рост) и QP /YPS (образование продукта)
ПОДДЕРЖАНИЕ ЖИЗНЕДЕЯТЕЛЬНОСТИ - 1
-
В клетках микроорганизмов всё время идут процессы деградации (разрушения) некоторых веществ (белков, нуклеиновых кислот и др.) и одновременно процессы их синтеза (репарации). На этот процесс необходимо расходовать часть субстрата .
-
Кроме того, субстрат расходуется на синтез определённых, но неизмеряемых продуктов, образующихся в небольшом количестве
ПОДДЕРЖАНИЕ ЖИЗНЕДЕЯТЕЛЬНОСТИ - 2
-
В процессе ферментации можно создать режим, при котором концентрация биомассы и продукта не изменяются (то есть ΔX = ΔP = 0), а субстрат всё время добавляется, предотвращая отмирание биомассы
-
Это дополнительное расходование биомассы происходит не только в такой искусственной ситуации, но и в течение всего процесса. Просто эти расходы обычно «приписывают» к расходам на рост и образование продукта.
ПОДДЕРЖАНИЕ ЖИЗНЕДЕЯТЕЛЬНОСТИ - 3
-
Правильнее выделить эту статью расхода в отдельный член уравнения – на поддержание жизнедеятельности микроорганизмов Q3
-
Очевидно, что эти затраты пропорциональны концентрации микроорганизмов X: Q3 = X*mS
-
Здесь mS – коэффициент поддержания жизнедеятельности или УДЕЛЬНАЯ скорость расходования субстрата на поддержание жизнедеятельности микроорганизмов [г субстрата / г биомассы в час]
ИТОГОВОЕ УРАВНЕНИЕ КИНЕТИКИ ПОТРЕБЛЕНИЯ СУБСТРАТА
-
Подставляя найденные зависимости, получаем уравнение
-
QS = (-dS / dt) = (1 /YXS )QX + (1 /YPS ) QP + mSX
-
В этом уравнении коэффициенты YXS и YPS соответствуют стехиометрическим и не «приписывают» себе затраты субстрата на другие компоненты процесса
СПОСОБ ОПРЕДЕЛЕНИЯ MS
-
Если пренебречь затратами на биосинтез продукта QP:
-
QS= QX(1/YXS) + mSX
-
или qS= (1/YXS) +mS
-
Графическое выражение зависимости представлено на графике qS()
-
В точке пересечения прямой с осью ординат ( = 0) имеем qS= mS
-
А с осью абсцисс (qS= 0)
-
= YXS*mS

СПОСОБ ОПРЕДЕЛЕНИЯ КОЭФФИЦИЕНТОВ УРАВНЕНИЯ
-
Находят по экспериментальным данным за определённый промежуток времени Δt четыре сопряжённых величины – приращения концентраций субстрата ΔS, биомассы ΔX, продукта ΔP и среднюю концентрацию биомассы за этот промежуток времени X
-
Для этой совокупности данных строят уравнение:
-
(ΔS/Δt) = (1/YXS)(ΔX/Δt) + (1/YPS)(ΔP/Δt) +X*mS
-
В этом уравнении 3 неизвестных коэффициента
-
По другим экспериментальным данным строят дополнительные уравнения, и искомые коэффициенты находят методом наименьших квадратов
ВАРИАНТЫ УРАВНЕНИЙ ДЛЯ MS
-
mS = 0 - (нет затрат на поддержание –характерно для конструктивных субстратов)
-
mS = m - (величина постоянна)
-
mS = KS - (затраты возрастают с повышением концентрации субстрата)
-
mS = m /(1 +S / Ki ) – (затраты возрастают при малых S)
СИСТЕМНАЯ МОДЕЛЬ ПРОЦЕССА ФЕРМЕНТАЦИИ
-
Модель процесса включает описание динамики изменения в ходе процесса трёх основных параметров: концентрации биомассы X, субстрата S, и продукта метаболизма P.
-
Система уравнений модели в общем виде:



КОНКРЕТИЗАЦИЯ МОДЕЛИ ПРОЦЕССА
-
Системная модель включает 5 кинетических переменных , каждая из которых может быть описана одним из вариантов уравнений (элементарных) для этой переменной

-
Выбор вида уравнений и определение коэффициентов в них проводится с помощью специальных экспериментов на реальном объекте