

2.2. Генетические карты.
Есть два вида хромосомных карт: морфологические (цитологические) и генетические (функциональные). Морфологические карты показывают какие гены и в какой последовательности находятся в хромосоме. Генетические карты показывают и расстояние между генами, лежащими в одной хромосоме.
Впервые Морган и его сотрудники использовали явление кроссинговера для составления генетических карт хромосом с использованием многочисленных мутантных генов. Генетическая карта хромосом – это схема взаимного расположения генов, находящихся в одной группе сцепления. Расстояние между генами на генетической карте хромосом определяют по частоте кроссинговера между ними (рис.).
Единицей расстояния принято считать одну морганиду (1% кроссинговера). Поскольку у человека способ составления генетических карт с помощью гибридологического анализа затруднен, разработаны другие способы определения локализации гена. Среди них:
F– анализ расщепления и сцепление признаков в семьях (по родословным); так были картированы, например, локусы группы крови АВО;
AAS– сравнение аминокислотных последовательностей белков;
CH– изменение морфологии хромосомного участка в сочетании с характерным фенотипом; к этой группе относят анализ “ломких” (фрагильных) сайтов в хромосомах. Фрагильные сайты полезны как маркеры в семейных исследованиях сцепления;
RE– рестрикционный анализ и реконструированные карты района;
ОТ – центромерное картирование и др.



 Образование
Возвратное
Образование
Возвратное
гетерозигот скрещивание
с рецессивным
А






 Ва вродителем
Ва вродителем




 х
х
А
 В а
в 70%
родительского типа 30% рекомбинантов
В а
в 70%
родительского типа 30% рекомбинантов

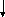














А Ва в 35% 35% 15% 15%
 х
х








 а ва в
а ва в















 А
Ва вА ва
В
А
Ва вА ва
В














 а
в а в а в а в
а
в а в а в а в






 А в
а в 15% 15% 35%
35%
А в
а в 15% 15% 35%
35%
 х
х
















 а Ва в
а Ва в
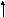


А




 ва В
ва В

 х30%
рекомбинантов 70% родительского типа
х30%
рекомбинантов 70% родительского типа
А




 ва В
ва В



Рис. Определение расстояния между генами при помощи анализирующего скрещивания.
Достоверно идентифицировано около полутора тысяч генов человека, что составляет 1-5% от общего числа.
У большинства из них обнаружены альтернативные формы. Около тысячи известных генов имеют хотя бы один из альтернативных аллелей, соответствующих какой-либо аномалии или заболеванию. Остальные гены кодируют белки группы крови, антигены, иммуноглобулины, ферменты и т. д. В настоящее время не существует принципиальных технических препятствий для получения полной карты генома человека, т.к. разработаны генно-инженерные подходы. В настоящее время геном человека полностью расшифрован.
2.3. Хромосомная теория наследственности.
В 1902 –1903 гг. для исследователя – Саттон (из США) и Бовери (из Германии) – независимо друг от друга предположили, что задатки (гены) расположены в хромосомах. Эта идея положила начало созданию хромосомной теории наследственности. Термин “ген” был предложен позже (в 1909 г. Иоганнсеном). Параллелизм в поведении генов и хромосом в процессе образования гамет убедительно говорил о том, что гены расположены в хромосомах. В 1910 г. хромосомная теория наследственности получила свое дальнейшее развитие в опытах Нобелевского лауреата Т. Моргана и его сотрудников, которые привели новые доказательства справедливости этой теории, показав связь между конкретными генами и хромосомами. Окончательные доказательства были получены Бриджесом в 1913 г., открывшем изменения в наследовании признаков, сцепленных с полом, при не расхождении половых хромосом.
Основные положения хромосомной теории наследственности следующие:
гены локализованы в хромосомах;
гены расположены в хромосомах в определенной линейной последовательности;
гены наследственно дискретны;
каждый ген имеет определенное место (локус) в хромосоме;
гены относительно стабильны;
гены могут изменяться (мутировать);
7) гены, локализованные в одной хромосоме, наследуются совместно, образуя группу сцепления;
число групп сцепления равно гаплоидному набору хромосом и постоянно для каждого вида организмов;
признаки, зависящие от сцепления генов, наследуются совместно;
сцепление генов может нарушаться процессом кроссинговера, в результате образуются рекомбинантные хромосомы;
частота кроссинговера является функцией расстояния между генами: чем больше расстояние, тем больше величина кроссинговера (прямая зависимость);
частота кроссинговера зависит от силы сцепления между генами: чем сильнее сцеплены гены, тем меньше величина кроссинговера (обратная зависимость);
сцепление генов и кроссинговер позволяют производить картирование хромосом.
На современном этапе развития хромосомная теория наследственности получила дальнейшее развитие благодаря изучению тонкой структуры и функции генов про- и эукариот. Эволюционировали наши представления о гене как о неделимой структуре. Открыты дробные структуры эукариотических генов, механизм их регуляции и т.д. Созданы искусственные гены. Все эти знания позволили сформировать один из фундаментальных принципов генетики о единстве дискретности и непрерывности генетического материала.