

3. Транскрипция и амплификация генов в оогенезе
Дифференцировка цитоплазмы яйца обусловлена функционированием хромосом,которое особенно выражено у видов с хромосомами типа ламповых щеток. Они имеют отчетливо выраженное хромомерное строение. Из хромомеров в виде петель вытянуты ДНК-вые оси хромосом. Поскольку хромосомы типа ламповых щеток существуют в диплотене и состоят из четырех хроматид, каждый участок таких хромосом представлен четырьмя хромомерами и четырьмя петлями. Петли — это участки хромомера с интенсивной транскрипцией. Обычно в них легко различают тонкий конец, где начинает свое движение РНК-полимераза, и толстый конец, где транскрипция заканчивается. Так называемый матрикс, которым покрыты петли, представляет собой гранулы или фибриллы, состоящие из вновь синтезированной РНК и белка, т. е. рибонуклеопротеиновых частиц. Некоторые петли имеют специфическую морфологию и легко идентифицируются. Существенно отметить, что число петель у тритонов — объектов, у которых хромосомы типа ламповых щеток изучены наиболее подробно,— близко к числу типов иРНК, присутствующих в цитоплазме. Эта иРНК используется не только для формирования цитоплазмы яйца.Большая часть молекул иРНК, синтезированных хромосомами типа ламповых щеток, не связана с рибосомами. Онаиспользуется позже во время раннего эмбриогенеза.
К числу других важных событий, осуществляющихся под генетическим контролем во время оогенеза, относится селективная амплификация (умножение числа копий) рибосомного гена, наблюдающаяся у некоторых животных, например у амфибий.Причина амплификации – резкое увеличение объема яйцеклетки по сравнению со средней соматической клеткой, достигающее иногда трех порядков. Чтобы заполнить такой огромный объем клетки рибосомами, гены рДНК сами увеличиваются в числе настолько, что, например, у Xenopus laevis по окончании амплификации содержание рДНК почти равно количеству ДНК, заключенному в диплоидном наборе хромосом. Число ядрышек — органелл, где происходит сборка рибосом, — соответственно возрастает с 2 до 1,5 тыс.Замечательная особенность молекулярногомеханизма амплификации заключается в том, что он осуществляется по принципу катящегося кольца. Одна из копий гена рДНК покидает хромосому, затем замыкается в кольцо, из которого как бы вытягивается хвост длиной в несколько десятков микрометров. Затем эта структура вновь циклизуется, образуя большое кольцо, на основе которого формируется ядрышко.
В последнее десятилетие явление амплификации отдельных генов было открыто не только в оогенезе. Так, устойчивость клеток к противораковым препаратам может быть обусловлена гиперпродукцией тех ферментов, которые они инактивируют.
4. Дифференциальная активность генов в онтогенезе
Развитие характеризуется сменой активности различных генов.Для того чтобы наглядно представить себе этот процесс, достаточно вспомнить основной биогенетическийзакон Э. Геккеля, согласно которому в эмбриогенезе каждого вида повторяются основные черты эмбрионального развития его эволюционных предшественников. Ясно, что гены, контролирующие образование зачаточных жаберных щелей и других подобных органов, функционируют в онтогенезе плацентарных млекопитающих лишь ограниченное время.
Четкая упорядоченность в связи со стадиями индивидуального развития установленадля смены функционирования генов гемоглобина у млекопитающих. Подобных примеров можно привести немало, однако с точки зрения генетики развития наиболее интересны те случаи, где дифференциальная активность генов может быть прослежена непосредственно по изменению некоторых особенностей хромосом, иногда называемых особенностями хромосомного фенотипа. Наиболее ярким примером такого рода служат пуфы в гигантских политенных хромосомах (рисунок 2). Морфологически они представляют собой вздутия определенных районов хромосом, обусловленные декомпактизацией отдельных дисков и интенсивным синтезом в них РНК- Пуфы, таким образом, можно рассматривать как высокоактивные в функциональном отношении тканеспецифичные и стадиеспецифичные гены. Установлена роль гормонов (в частности, экдизона — гормона окукливания в индукции пуфов), а также роль белков, синтезированных ранними пуфами, в индукции поздних пуфов.
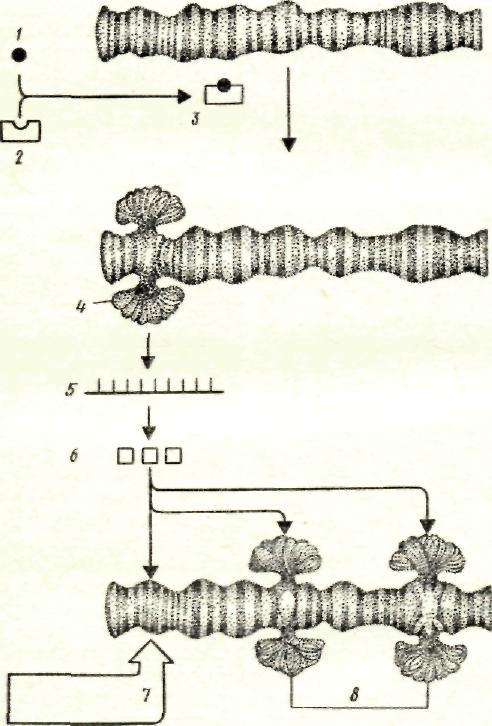
Рисунок 2 – Пуфирование гигантских хромосом в клетках слюнных желез дрозофилы иод действием экдизона: 1 — экдизон, 2— рецепгорный белок, 3— комплекс экдизона и рецепторного белка. 4 - ранний пуф. 5 — иРНК, 6 — белки — продукты раннего пуфа, 7 — регрессия раннего пуфа, 8 — поздние пуфы, возникшие под действием продуктов раннего пуфа
Таким образом, стероидные гормоны и белки, вероятно, не единственные факторы,ответственные за переключение генов в онтогенезе, а следовательно, и за смену фаз индивидуального развития организма. Особенно велика роль стероидных гормонов в регуляции генной активности у животных. Известно, что гормоны синтезируются в специализированных клетках желез внутренней секреции и циркулируют по всему организму. Однако отдельные гормоны активируют гены не во всех клетках, а только в клетках-мишенях, которые содержат специальные рецепторные белки, с которыми специфически связываются молекулы гормона. Это связывание происходит в цитоплазме, а затем образовавшийся комплекс проникает в ядро, где он взаимодействует с определенными негистоновыми белками хромосом. В отсутствие гормонов эти белки блокируют либо промоторные, либо иные, пока неизвестные регуляторные участки определенных генов. Комплекс «гормон — рецепторный белок» снимает блокирующее действие негистонового белка-репрессора, следствием чего являются транскрипция данного гена, созревание иРНК, транспорт ее в цитоплазму и синтез белка.