

8.2.6. Эмбриональная индукция
Эмбриональная индукция — это взаимодействие частей развивающегося зародыша, при котором один участок зародыша влияет на судьбу другого участка. Явление эмбриональной индукции с начала XX в. изучает экспериментальная эмбриология.
Классическими считают опыты немецкого ученого Г. Шпемана и его сотрудников (1924) на зародышах амфибий. Для того чтобы иметь возможность проследить за судьбой клеток определенного участка зародыша, Шпеман использовал два вида тритонов: тритона гребенчатого, яйца которого лишены пигмента и потому имеют белый цвет, и тритона полосатого, яйца которого благодаря пигменту имеют желто-серый цвет.
Один из опытов заключается в следующем: кусочек зародыша из области дорсальной губы бластопора на стадии гаструлы тритона гребенчатого пересаживают на боковую или вентральную сторону гаструлы тритона полосатого (рис. 8.8). В месте пересадки происходит развитие нервной трубки, хорды и других органов. Развитие может достичь довольно продвинутых стадий с образованием дополнительного зародыша на боковой или вентральной стороне зародыша реципиента. Дополнительный зародыш содержит в основном клетки зародыша реципиента, но светлые клетки зародыша-донора тоже обнаруживаются в составе различных органов.
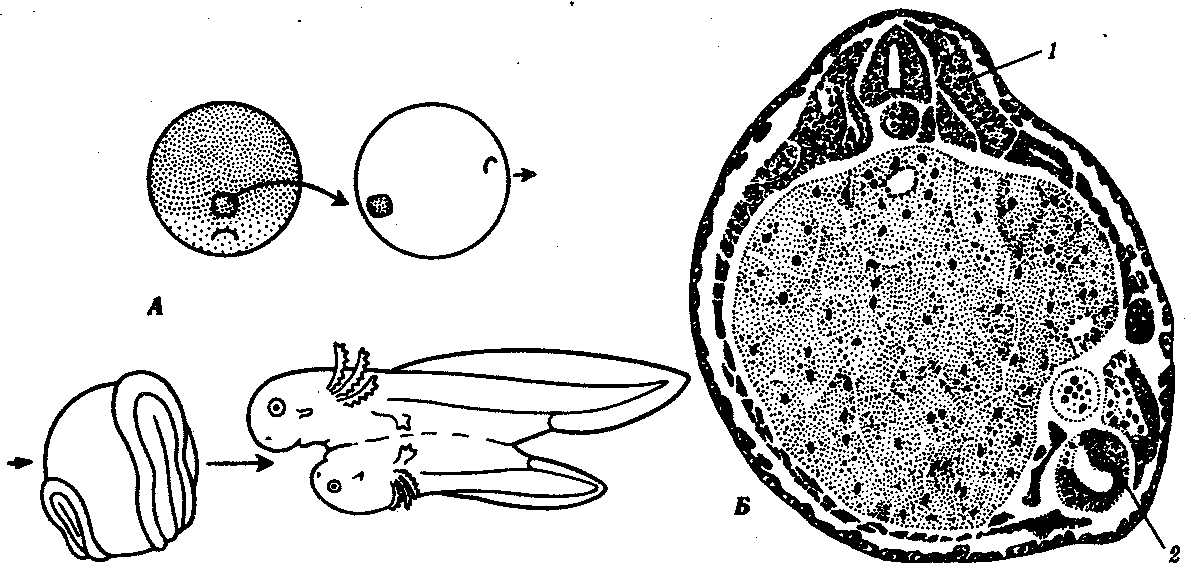
Рис. 8.8. Пересадка спинной губы от зародыша-донора на брюшную сторону зародыша-реципиента. А—схема опыта; Б—поперечный срез на стадии закладки двух комплексов осевых органов:
1—первичный зародыш, 2—вторичный, индуцированный зародыш
Из этого и подобных опытов следует несколько выводов. Во-первых, участок, взятый из спинной губы бластопора, способен направлять или даже переключать развитие того материала, который находится вокруг него, на определенный путь развития. Он как бы организует, или индуцирует, развитие зародыша как в обычном, так и в нетипичном месте. Во-вторых, боковая и брюшная стороны гаструлы обладают более широкими потенциями к развитию, нежели их презумптивное (предполагаемое) проспективное направление, так как вместо обычной поверхности тела в условиях эксперимента там образуется целый зародыш. В-третьих, достаточно точное строение новообразованных органов в месте пересадки указывает на эмбриональную регуляцию. Это означает, что фактор целостности организма приводит к достижению хорошего конечного результата из нетипичных клеток в нетипичном месте, как бы управляя процессом, регулируя его в целях достижения этого результата.
Г. Шпеман назвал спинную губу бластопора первичным эмбриональным организатором. Первичным потому, что на более ранних стадиях развития подобных влияний обнаружить не удавалось, а организатором потому, что влияние происходило именно на морфогенез. В настоящее время установлено, что главная роль в спинной губе бластопора принадлежит хордомезодермальному зачатку, который назвали первичным эмбриональным индуктором, а само явление, при котором один участок зародыша влияет на судьбы другого,— эмбриональной индукцией.
В 30-е гг. исследователи пытались установить природу индуцирующего действия. Вскоре выяснилось, что разнообразные убитые ткани, вытяжки из самых различных тканей беспозвоночных и позвоночных животных, а также растений, несколько классов химических соединений (белки, нуклеопротеины, стероиды и даже неорганические вещества) могут вызывать индукцию. Таким образом была установлена химическая природа организаторов. Одновременно стало ясно, что специфичность ответа прямо не связана с. химическими свойствами индуктора.
Внимание эмбриологов переключилось на индуцируемые ткани. Оказалось, что специфичность действия индуктора-раздражителя может быть весьма различной, а сам эффект индуцирующего воздействия ограничивается способностью того или иного участка развивающегося зародыша воспринимать это воздействие и отвечать на него.
Некоторые индукторы, по-видимому, более или менее специфичны в определении судьбы индуцируемой ткани. Об этом свидетельствуют следующие опыты. Если пересадить спинную губу ранней гаструлы, то индуцируется развитие структур переднего мозга (головной индуктор), если же пересадить спинную губу поздней гаструлы, то развиваются спинной мозг и мезодермальные ткани (туловищный индуктор, рис. 8.9). Было показано также, что наиболее сильное нейрализующее влияние оказывает фракция нуклеопротеинов, а мезодермализующим индуктором оказался белок. Если имплантировать оба эти индуктора в виде смеси клеток или смеси веществ, то получаются хорошо развитые зародыши.

Рис. 8.9. Результаты пересадки головного (А) и туловищного (Б) индукторов Объяснение см. в тексте
Другие индукторы действуют как неспецифические пусковые механизмы, как бы высвобождая ответ, уже детерминированный в клетках индуцируемой ткани. Было показано, что, например, слуховой пузырек выступает не только в роли индуктора слухового аппарата, но и является активатором различных морфогенетических процессов. Будучи пересажен в область боковой линии эмбриона тритона, он влечет за собой индукцию конечности. Конечность можно индуцировать также пересадкой носовой плакоды или гипофиза. Легче всего добавочные конечности индуцируются в области боковой линии, но они могут быть получены и на брюшной стороне. Эти примеры указывают на то, что специфический ответ зависит не столько от индуктора, сколько отреагирующей области.
Способность эмбрионального материала реагировать на различного рода влияния изменением своей презумптивной судьбы получила название компетенции. Установлено, например, что компетенция к образованию нервной системы у амфибий затрагивает всю эмбриональную эктодерму и возникает с момента начала гаструляции. К концу гаструляции эта компетенция прекращается. Таким образом, изменение хода развития возможно лишь в том случае, если область компетенции к образованию некоторой закладки шире, чем область, из которой она в норме развивается, а также если индукционное действие происходит в определенный интервал онтогенетического развития.
Изучение индукционных взаимодействий у разных представителей типа хордовых показало, что области и сроки компетенции неодинаковы. Так, у асцидий на стадии 8 бластомеров, когда уже все основные зачатки предопределены, проводили некоторые перемещения бластомеров. Материал хордомезодермы и основная часть нейрального материала у них локализованы в заднем вегетативном бластомере. Небольшая часть нейрального материала, формирующего головной ганглий, находится в заднем анимальном бластомере, расположенном над задним вегетативным (рис. 8.10).
Для проверки индукционных взаимодействий между ними анимальный ярус бластомеров поворачивали на 180° так, чтобы задний анимальный бластомер терял контакт с задним вегетативным. Головной ганглий не развился нигде. Это означает, что для развития головного ганглия необходимо индукционное влияние на задний анимальный бластомер со стороны заднего вегетативного. Кроме того, очевидно, что задний анимальный бластомер не обладает автономностью развития, но только он компетентен к восприятию воздействия со стороны заднего вегетативного бластомера, содержащего хордомезодермальный зачаток.
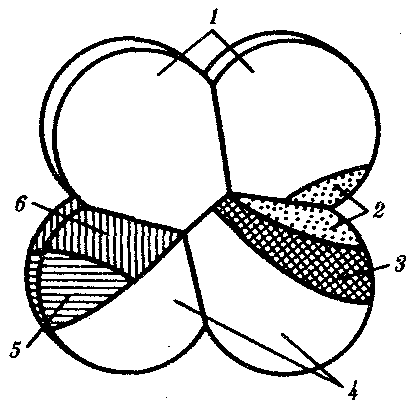
Рис. 8.10. Карта презумптивных зачатков у зародыша асцидий
на стадии восьми бластомеров:
1—эпидермис, 2—нервная пластинка, 3—хорда, 4—энтодерма, 5—сомиты, 6—мезенхима
Во всех других классах хордовых индукционные взаимодействия между хордомезодермальным и нейральным зачатками подобны таковым у амфибий. Полагают, что в ходе эволюции хордовых произошли расширение областей и удлинение срока компетенции. Это расценивают как признак существенного эволюционного прогресса.
Явления индукции многочисленны и разнообразны. Помимо первичной индукции со стороны спинной губы бластопора описаны индукционные влияния на более поздних, нежели гаструляция, этапах развития. Все они являются вторичными и третичными, представляя собой каскадные взаимодействия, типичные для дифференцировки, потому что индукция многих структур зависит от предшествующих индукционных событий. Примером вторичной индукции может служить действие глазного бокала (выпячивание переднего мозга) на прилежащий покровный эпителий, под влиянием чего эпителий впячивается, а затем отшнуровывается хрусталиковый пузырек—зачаток глазного хрусталика (рис. 8.11). Расположенный над хрусталиком покровный эпителий тоже испытывает сложные изменения, теряет пигмент и становится роговичным эпителием. Это пример третичной индукции. Таким образом получается, что глазной бокал возникает только после развития передней части головного мозга, хрусталик — после формирования бокала, а роговица — после образования хрусталика.
Вместе с тем индукция носит не только каскадный, но и переплетающийся характер, т.е. в индукции той или иной структуры может участвовать не одна, а несколько тканей. В свою очередь, такая структура может служить индуктором для нескольких других тканей. Например, глазной бокал служит главным, но не единственным индуктором хрусталика. Морфогенез всегда сопровождается значительными перемещениями тканей друг относительно друга. Так, презумптивный хрусталик, т.е. эпидермис, из которого в последующем должен развиться хрусталик, во время гаструляции лежит над энтодермой будущей глотки, которая служит первым индуктором хрусталика. Затем под этим эпидермисом оказывается сердечная мезодерма, которая тоже действует как индуктор. И только позднее, во время нейруляции на переднем конце нервной трубки выпячиваются глазные пузыри, образующие глазной бокал и сетчатку, являющуюся главным индуктором хрусталика (рис. 8.12).
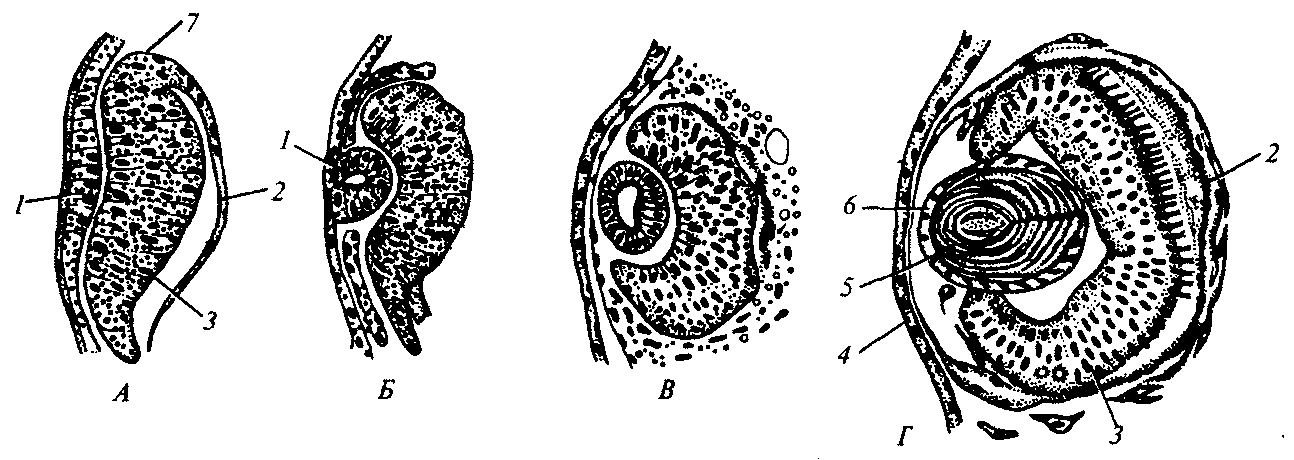
Рис. 8.11. Развитие (А —Г) глаза у хвостатой амфибии:
1—хрусталиковая плакода, 2—пигментный эпителий, 3—сетчатка, 4—роговица, 5—хрусталиковые волокна, 6—хрусталиковый эпителий, 7—глазной бокал
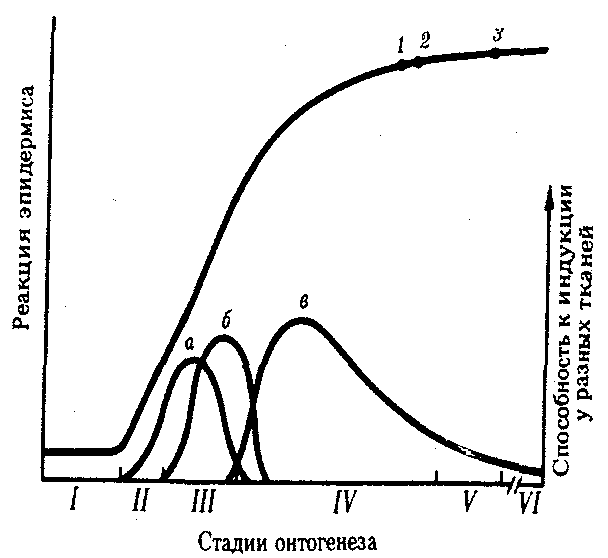
Рис. 8.12. Последовательные индукционные взаимодействия,
необходимые для образования хрусталика у зародыша амфибии:
I—ранний зародыш, II—гаструла, III—нейрула, IV—стадия хвостовой почки, V—личинка, VI—взрослая особь; 1—плакода, 2—пузырек, 3—волокна, а—энтодерма, б—сердечная мезодерма, в—сетчатка
Удаляя ту или иную из индуцирующих тканей, определили степень участия каждой из них в индукции хрусталика. Оказалось, что при удалении сетчатки глазного бокала у 42% зародышей амфибий все же формировались хрусталики и, следовательно, энтодерма и мезодерма в сумме обладают почти таким же индуцирующим действием, как и сетчатка глазного бокала. Полагают, что многочисленность индуцирующих тканей может иметь решающее значение для точного установления места формирования органа. Кроме того, сети индукции могут играть важную роль в канализации развития, обеспечивая нормальное течение органогенеза, даже если один из компонентов ин-
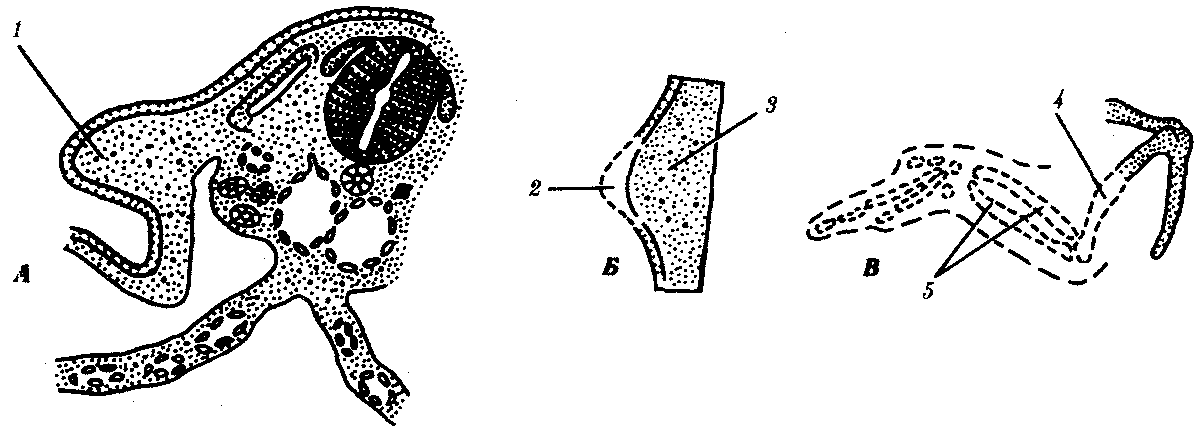
Рис. 8.13. Влияние удаления апикального эктодермального гребня
на развитие почки крыла.
А—схема расположения почки крыла; Б—почка крыла; В—недоразвитие скелета (пунктир) после удаления апикального гребня:
1—почка крыла, 2 —апикальный эктодермальный гребень, 3—мезенхима, 4 —плечевая кость, 5—локтевая и лучевая кости
Чаще всего близлежащие участки зародыша оказывают взаимное влияние друг на друга. Демонстративным примером являются взаимодействия в зачатке конечности. Конечность развивается из скопления клеток, происходящих из боковой мезодермы, и покрывающих их клеток эктодермы (рис. 8.13). Развитие конечности начинается с активации клеток боковой мезодермы в непосредственной близости от сомитов, которые, возможно, и оказывают индуцирующие импульсы на мезодерму в области будущей конечности. Активированные мезодермальные клетки зачатка конечности влияют на покрывающую их эктодерму, в результате чего она утолщается. Образовавшееся утолщение эпидермиса на его верхушке называют апикальным эктодермальным гребнем. Последний стимулирует рост почки конечности (при удалении его рост почки конечности прекращается). Мезодерма же поддерживает гребень в активном состоянии и определяет форму конечности. Например, мезодерма из почки крыла при соединении с эктодермой почки ноги образует крыло, покрытое перьями, или мезодерма из почки конечности утиного зародыша с эктодермой куриного приводит к развитию перепончатой конечности.
Различают гетерономную и гомономную виды индукции. К гетерономной относят случаи, подобные описанному, при которых один кусочек зародыша индуцирует иной орган (хордомезодерма индуцирует появление нервной трубки и всего зародыша в целом). Гомономная индукция заключается в том, что индуктор побуждает окружающий материал к развитию в том же направлении, что и он сам. Например, область нефротома, пересаженная другому зародышу, способствует развитию окружающего материала в сторону формирования головной почки, а прибавление в культуру фибробластов сердца маленького кусочка хряща влечет за собой процесс образования хряща.
Чтобы воспринять действие индуктора, компетентная ткань должна обладать хотя бы минимальной организацией. Одиночные клетки не воспринимают действие индуктора, а чем больше клеток в реагирующей ткани, тем активнее ее реакция. Для оказания индуцирующего действия иногда достаточно лишь одной клетки индуктора.
Индукционные взаимодействия могут проявляться в культуре ткани in vitro, но по-настоящему полноценными они бывают только в структуре целостного организма.
Весьма интересны результаты опытов, помогающие оценить взаимосвязь индукционных взаимодействий с цитодифференцировкой и морфогенезом. Ранее уже было описано определяющее влияние мезенхимы на морфогенез конечностей позвоночных.
Многочисленными опытами показано также большое влияние мезенхимы на морфогенез желез эпителиального происхождения. Легочная энтодерма, например, при выращивании с печеночной мезенхимой приобретает строение печеночных балок, а эпителий молочной железы под влиянием мезенхимного зачатка слюнной железы приобретает морфологию слюнной железы. Это происходит как при выращивании in vitro, так и при трансплантации в организм животного-реципиента. Подобные результаты с несомненностью указывают на необходимость индуцирующего влияния мезенхимы на морфогенез.
Однако не менее интересен факт, что морфогенез не всегда сопряжен с определенным направлением дифференцирован эпителия. Так, рекомбинантная слюнная железа, полученная из зачатка молочной железы и мезенхимы слюнной, при подсадке лактирующей самке-реципиенту начинает вырабатывать молоко несмотря на то, что имеет морфологию по типу слюнной железы. Это свидетельствует о возможности разобщения, об автономности процессов морфогенеза и цитодифференцировки и может быть объяснено более ранней детерминацией цитодифференцировки другими, более ранними актами индукции. Подобные наблюдения позволяют по-другому взглянуть на возможности преобразований морфогенезов в процессе эволюции.
Таким образом, явления индукции обнаружены на самых разных этапах развития многих позвоночных. В акте индукции следует различать два компонента: индуктор и реагирующую область. Изложенные выше положения кратко обобщены на схеме 8.4.
В настоящее время интенсивно ведутся работы по изучению молекулярных и клеточных механизмов индукции. В теоретическом смысле явление эмбриональной индукции помогает по-новому оценить взаимоотношение таких процессов, как зависимая дифференцировка и детерминация, а также цитодифференцировка и морфогенез. Понятия детерминации и морфогенеза более подробно рассматриваются в § 8.3.
