

2. 1. 4. Механізм реплікації днк
Реплікація – унікальна властивість молекули ДНК до самоподвоєння. Клітина передає генетичний матеріал своїм нащадкам, реплікуючи (подвоюючи) весь свій геном перед поділом, під час S-періоду інтерфази. Цей процес повинен відбутися протягом 7–12 год без помилок, без втрати частини геному і без надмірних копіювань окремих частин ДНК.
Під час конваріантної реплікації два ланцюги спіралі ДНК розділяються, і кожна нитка стає матрицею для синтезу нового дочірнього ланцюга. Отже, у кожній із двох нових молекул ДНК один ланцюг старий, а інший – заново синтезований. Такий тип реплікації має назву напівконсервативний. Завдяки здатності азотистих основ нуклеотидів утворювати за допомогою водневих зв’язків комплементарні пари нуклеотидна послідовність молекул ДНК однакова у двох дочірніх клітинах і ідентична вихідній материнській молекулі.
Процес реплікації починається у декількох ділянках ДНК – точках (сайтах) ініціації реплікації (спеціальні послідовності нуклеотидів – локуси ori). У кожній із них на ДНК діють декілька білків. Спочатку фермент ДНК-геліказа „розплітає” спіраль ДНК, роз’єднуючи водневі зв’язки між двома ланцюгами. Потім до кожного із ланцюгів приєднуються дестабілізувальні білки, щоб не дати можливості ланцюгам знову утворити водневі зв’язки і відновити спіраль. Для того щоб спіраль розпліталася легше і з меншими затратами енергії, фермент ДНК-топоізомераза „розрізає” один із ланцюгів ДНК у певному місці, і він починає розкручуватися навколо іншого ланцюга (потім ДНК-топоізомераза заново зшиває розрізані фрагменти ДНК). Так утворюється реплікаційна вилка.
Наявні в каріоплазмі нуклеотиди приєднуються до материнського ланцюга своїми азотистими основами згідно з принципом комплементарності (А–Т; Г–Ц), утворюючи водневі зв’язки з нуклеотидами материнського ланцюга. Потім фермент ДНК-полімераза з’єднує сусідні нуклеотиди між собою. Під час приєднання нуклеотидів між ОН–групою одного нуклеотиду та залишком фосфорної кислоти іншого утворюється фосфодиефірний зв’язок. Оскільки ДНК-полімераза не може починати синтез дочірнього ланцюга з „нуля”, а може лише приєднувати нуклеотиди до вільного ОН-кінця (він знаходиться на 3′–кінці дочірнього ланцюга, що росте), то необхідна певна послідовність нуклеотидів із вільним ОН–кінцем, яка слугує початком ланцюга – праймером. Ним є послідовність РНК із 10–60 нуклеотидів, що має назву РНК-затравка, або праймер. Праймер синтезується за участю ферменту РНК-праймази. Таким чином, синтез нового дочірнього ланцюга відбувається у напрямку 5′→3′ (на материнській нитці ДНК 3′→5′). Цей дочірній ланцюг ДНК синтезується швидко, тому має назву провідного, або лідируючого ланцюга (рис. 2).
Оскільки дві нитки ДНК у складі спіралі антипаралельні (одна нитка 3′→5′, а інша 5′→3′), а ДНК-полімераза може „працювати” лише в одному напрямку (5′→3′), то процес синтезу дочірньої нитки ДНК на кожному із материнських ланцюгів відбувається по-різному. На матриці 3′→5′ новий ланцюг синтезується безперервно і швидко (лідируючий ланцюг). На матриці ж 5′→3′ ланцюг повинен синтезуватися у відповідному напрямку (3′→5′), але це неможливо, оскільки ДНК-полімеразі потрібний вільний 3′–кінець для приєднання нуклеотидів. Тому цей дочірній ланцюг синтезується короткими фрагментами (фрагментами Оказакі), які також синтезуються у напрямку 5′→3′. Їх синтез починається із праймерів (РНК-затравок), які потім добудовуються ферментом ДНК-полімеразою. Після того як сусідні фрагменти повністю добудовуються нуклеотидами і примикають один до одного, РНК-затравки видаляються за допомогою ферменту РНК-нуклеази. Утворені порожнини заповнюються нуклеотидами за допомогою ДНК-полімерази, а потім сусідні фрагменти зшиваються за допомогою ферменту ДНК-лігази. (Таким же чином видаляється РНК-затравка на початку лідируючого дочірнього ланцюга.)
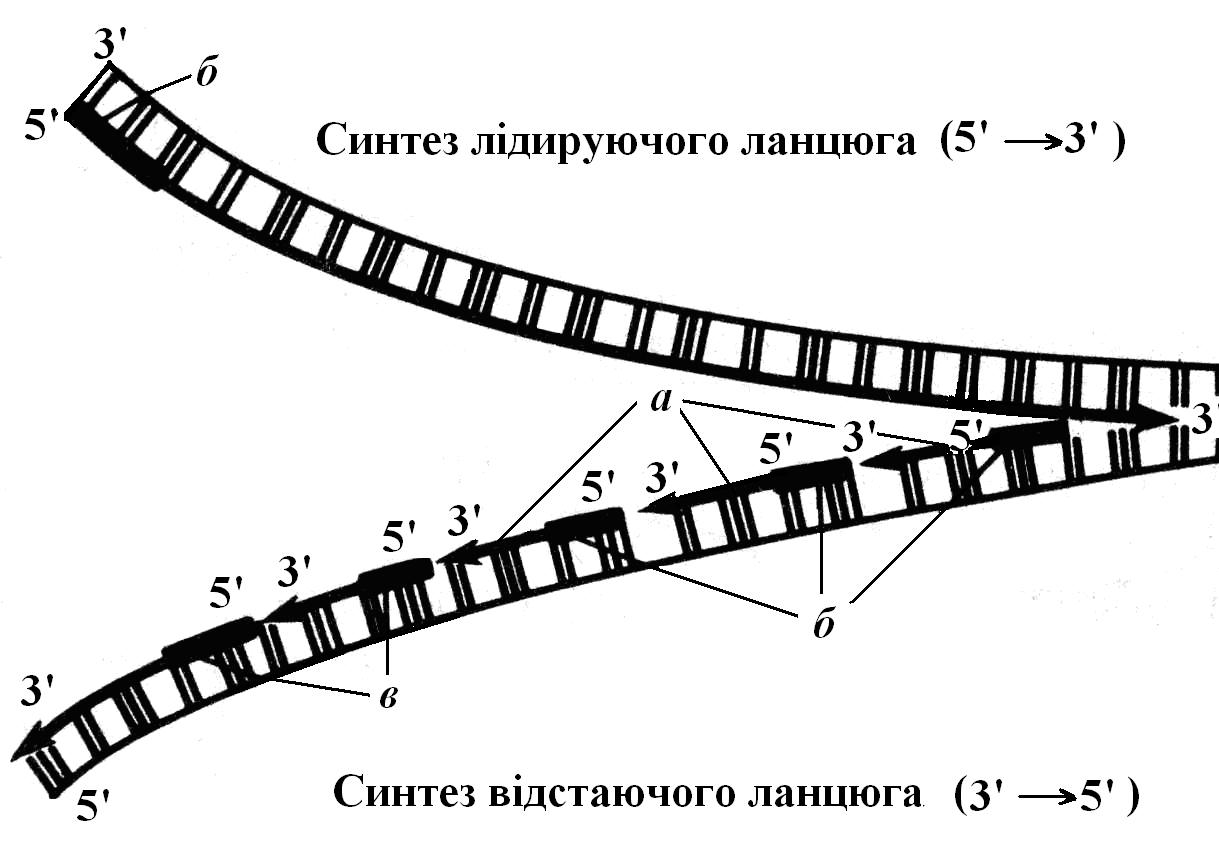
Рис. 2. Реплікація ДНК
а – фрагменти Оказакі; б – РНК-затравки (синтезуються РНК-праймазою); в – ДНК-лігаза зшиває фрагменти Оказакі в єдиний ланцюг.
Хоча окремі фрагменти Оказакі синтезуються у напрямку 5′→3′, дочірній ланцюг в цілому подовжується у напрямку 3′→5′. Цей ланцюг синтезується довше, ніж провідний, тому має назву відстаючий ланцюг.
Фрагменти Оказакі в еукаріотів, у тому числі і в людини, містять 1000–2000 нуклеотидів. Швидкість синтезу ДНК в еукаріотів – 100 нуклеотидів за секунду. Швидкість реплікації ДНК у людини відносно низька: швидкість синтезу 1 мкм ДНК складає 0,5 мкм/хв. Якби синтез ДНК починався із однієї точки і безперервно продовжувався б до кінця хромосоми, реплікація однієї хромосоми людини довжиною 7 см відбувалася б протягом 3 міс. Тривалість же S-періоду інтерфази складає 7–12 год. Тому в молекулі ДНК будь-якої хромосоми є багато точок ініціації реплікації, кожна з яких є сигналом для синтезу свого реплікона.
Реплікон – фрагмент ДНК від однієї точки ініціації реплікації до іншої (від одного локусу ori до іншого). У людини їх ≈ 50 тис., довжина кожного ≈ 30 мкм. Синтез різних репліконів може відбуватися асинхронно і протягом різного часу. Від кожного реплікона реплікація відбувається в обох напрямках доти, доки сусідні реплікони не зіллються. Тому реплікація ДНК у кожній хромосомі проходить відносно швидко.
Кінцевим результатом процесу реплікації є утворення двох молекул ДНК, нуклеотидна послідовність яких повністю ідентична материнській спіралі ДНК.
Якщо який-небудь із ферментів реплікації має знижену активність, це викликає захворювання. Наприклад, порушення активності ДНК-гелікази призводить до розвитку синдрому Вернера – надшвидкого старіння (20-річна людина виглядає, як 70–80-річна) або синдрому Блума (раннього виникнення новоутворень).