

Функции мозжечка
Мозжечок контролирует движения тела. К мозжечку приходят импульсы от всех рецепторов, которые раздражаются во время движений тела. Функция мозжечка может нарушаться при принятии алкоголя или других веществ, вызывающих головокружение. Поэтому под действием опьянения люди не способны нормально координировать свои движения. В последние годы появляется все больше доказательств, что мозжечок имеет значение и в познавательной деятельности человека.
Черепно-мозговые нервы
Помимо спинного мозга очень важны и двенадцать черепно-мозговых нервов: I и II пары -обонятельный и зрительный нервы; III, IV VI пары - глазодвигательные нервы; V пара -тройничный нерв - иннервирует жевательные мышцы; VII - лицевой нерв - иннервирует мимические мышцы, содержит также секреторные волокна к слезной и слюнным железам; VIII пара - преддверно-улитковый нерв - связывает органы слуха, равновесия и гравитации; IX пара - языкоглоточный нерв — иннервирует глотку, ее мышцы, околоушную железу, вкусовые почки языка; X пара - блуждающий нерв -разделяется на ряд ветвей, которые иннервируют легкие, сердце, кишечник, регулируют их функции; XI пара - добавочный нерв - иннервирует мышцы плечевого пояса. В результате слияния спинномозговых нервов образуется XII пара - подъязычный нерв - иннервирует мышцы языка и подъязычный аппарат.
Развитие и дифференциация структур Нервная система у высокоорганизованных животных обусловили её разделение на соматическую и вегетативную нервную систему. Особенность строения вегетативной Нервная система та, что её волокна, отходящие от ЦНС, не доходят непосредственно до эффектора, а сначала вступают в периферические ганглии, где оканчиваются на клетках, отдающих аксоны уже непосредственно на иннервируемый орган. В зависимости от того, где расположены ганглии вегетативной Нервная система, и некоторых её функциональных особенностей вегетативную Нервная система делят на 2 части: парасимпатическую и симпатическую.
Структурная и функциональная единица Нервная система - нейрон, состоящий из тела нервной клетки и отростков - аксона и дендритов. Кроме нервных клеток, в структуру Нервная система входят глиальные клетки. Нейроны являются в известной мере самостоятельными единицами - их протоплазма не переходит из одного нейрона в другой. Взаимодействие между нейронами осуществляется благодаря контактам между ними - синапсам. В области контакта между окончанием одного нейрона и поверхностью другого в большинстве случаев сохраняется особое пространство - синаптическая щель - шириной в несколько сот ангстрем. Основные функции нейронов: восприятие раздражений, их переработка, передача этой информации и формирование ответной реакции. В зависимости от типа и хода нервных отростков (волокон), а также их функций нейроны подразделяют на: а) рецепторные (афферентные), волокна которых проводят нервные импульсы от рецепторов в ЦНС; тела их находятся в спинальных ганглиях или ганглиях черепномозговых нервов; б) двигательные (эфферентные), связывающие ЦНС с эффекторами; тела и дендриты их находятся в ЦНС, а аксоны выходят за её пределы (за исключением эфферентных нейронов вегетативной Нервная система, тела которых расположены в периферических ганглиях); в) вставочные (ассоциативные) нейроны, служащие связующими звеньями между афферентными и эфферентными нейронами; тела и отростки их расположены в ЦНС.
Деятельность Нервная система основывается на двух процессах: возбуждении и торможении. Возбуждение может быть распространяющимся (см. Импульс нервный) или местным - нераспространяющимся, стационарным (последнее открыто Н. Е. Введенским в 1901) Торможение - процесс, тесно связанный с возбуждением и внешне выражающийся в снижении возбудимости клеток. Одна из характерных черт тормозного процесса - отсутствие способности к активному распространению по нервным структурам (явление торможения в нервных центрах впервые было установлено И. М. Сеченовым в 1863).
Клеточные механизмы возбуждения и торможения подробно изучены. Тело и отростки нервной клетки покрыты мембраной, постоянно несущей на себе разность потенциалов (так называемый мембранный потенциал). Раздражение расположенных на периферии чувствительных окончаний афферентного нейрона преобразуется в изменение этой разности потенциалов (см. Биоэлектрические потенциалы). Возникающий вследствие этого нервный импульс распространяется по нервному волокну и достигает его пресинаптического окончания, где вызывает выделение в синаптическую щель высокоактивного химического вещества - медиатора. Под влиянием последнего в постсинаптической мембране, чувствительной к действию медиатора, происходит молекулярная реорганизация поверхности. В результате постсинаптическая мембрана начинает пропускать ионы и деполяризуется, вследствие чего на ней возникает электрическая реакция в виде местного возбуждающего постсинаптического потенциала (ВПСП), вновь генерирующего распространяющийся импульс. Нервные импульсы, возникающие при возбуждении особых тормозящих нейронов, вызывают гиперполяризацию постсинаптической мембраны и, соответственно, тормозящий постсинаптический потенциал (ТПСП). Помимо этого, установлен и др. вид торможения, формирующийся в пресинаптической структуре, - пресинаптическое торможение, обусловливающее длительное снижение эффективности синаптической передачи . В основе деятельности Нервная система лежит рефлекс, т. е. реакция организма на раздражения рецепторов, осуществляемая при посредстве Нервная система Термин «рефлекс» был впервые введён в зарождавшуюся физиологию Р. Декартом в 1649, хотя конкретных представлений о том, как осуществляется рефлекторная деятельность, в то время ещё не было. Такие сведения были получены лишь значительно позже, когда морфологи приступили к исследованию строения и функций нервных клеток. Все рефлекторные процессы связаны с распространением возбуждения по определённым нервным структурам - рефлекторным дугам. Основные элементы рефлекторной дуги: рецепторы, центростремительный (афферентный) нервный путь, внутрицентральные структуры различной сложности, центробежный (эфферентный) нервный путь и исполнительный орган (эффектор). Различные группы рецепторов возбуждаются раздражителями разной модальности (т. е. качественной специфичности) и воспринимают раздражения, исходящие как из внешней среды (экстерорецепторы - органы зрения, слуха, обоняния и др.), так и из внутренней среды организма (интерорецепторы, возбуждающиеся при механических, химических, температурных и др. раздражениях внутренних органов, мышц и др.). Нервные сигналы, несущие в ЦНС информацию от рецепторов по нервным волокнам, лишены модальности и обычно передаются в виде серии однородных импульсов. Информация о различных характеристиках раздражений кодируется изменениями частоты импульсов, а также приуроченностью нервной импульсации к определённым волокнам (так называемое пространственно-временное кодирование). Совокупность рецепторов данной области тела животного или человека, раздражение которых вызывает определённый тип рефлекторной реакции, называется рецептивным полем рефлекса. Такие поля могут накладываться друг на друга. Совокупность нервных образований, сосредоточенных в ЦНС и ответственных за осуществление данного рефлекторного акта, обозначают термином нервный центр. На отдельном нейроне в Нервная система может сходиться огромное число окончаний волокон, несущих импульсы от др. нервных клеток. В каждый данный момент в результате сложной синаптической переработки этого потока импульсов обеспечивается дальнейшее проведение лишь одного, определённого сигнала - принцип конвергенции, лежащий в основе деятельности всех уровней Нервная система («принцип конечного общего пути» Шеррингтона, получивший развитие в трудах Ухтомского и др.). Пространственно-временная суммация синаптических процессов служит основой для различных форм избирательного функционального объединения нервных клеток, лежащего в основе анализа поступающей в Нервная система информации и выработки затем команд для выполнения различных ответных реакций организма. Такие команды, как и афферентные сигналы, передаются от одной клетки к другой и от ЦНС к исполнительным органам в виде последовательностей нервных импульсов, возникающих в клетке в том случае, когда суммирующиеся возбуждающие и тормозящие синаптические процессы достигают определённого (критического для данной клетки) уровня - порога возбуждения. Несмотря на наследственно закрепленный характер связей в основных рефлекторных дугах, характер рефлекторной реакции может в значительной степени изменяться в зависимости от состояния центральных образований, через которые они осуществляются. Так, резкое повышение или понижение возбудимости центральных структур рефлекторной дуги может не только количественно изменить реакцию, но и привести к определённым качественным изменениям в характере рефлекса. Примером такого изменения может служить явление доминанты. Важное значение для нормального протекания рефлекторной деятельности имеет механизм так называемой обратной афферентации - информации о результате выполнения данной рефлекторной реакции, поступающей по афферентным путям от исполнительных органов. На основании этих сведений в случае, если результат неудовлетворителен, в сформировавшейся функциональной системе могут происходить перестройки деятельности отдельных элементов до тех пор, пока результат не станет соответствовать уровню, необходимому для организма (П. К. Анохин, 1935). Всю совокупность рефлекторных реакций организма делят на две основные группы: безусловные рефлексы - врождённые, осуществляемые по наследственно закрепленным нервным путям, и условные рефлексы, приобретённые в течение индивидуальной жизни организма путём образования в ЦНС временных связей. Способность образования таких связей присуща лишь высшему для данного вида животных отделу Нервная система (для млекопитающих и человека - это кора головного мозга). Образование условнорефлекторных связей позволяет организму наиболее совершенно и тонко приспосабливаться к постоянно изменяющимся условиям существования. Условные рефлексы были открыты и изучены И. П. Павловым в конце 19 - начале 20 вв. Исследование условнорефлекторной деятельности животных и человека привело его к созданию учения о высшей нервной деятельности (ВНД) и анализаторах. Каждый анализатор состоит из воспринимающей части - рецептора, проводящих путей и анализирующих структур ЦНС, обязательно включающих её высший отдел. Кора головного мозга у высших животных - совокупность корковых концов анализаторов; она осуществляет высшие формы анализаторной и интегративной деятельности, обеспечивая совершеннейшие и тончайшие формы взаимодействия организма с внешней средой. Нервная система обладает способностью не только немедленно перерабатывать поступающую в неё информацию при помощи механизма взаимодействующих синаптических процессов, но и хранить следы прошлой активности (механизмы памяти). Клеточные механизмы сохранения в высших отделах Нервная система длительных следов нервных процессов, лежащие в основе памяти, интенсивно изучаются. Наряду с перечисленными выше функциями Нервная система осуществляет также регулирующие влияния на обменные процессы в тканях - адаптационно-трофическую функцию (И. П. Павлов, Л. А. Орбели, А. В. Тонких и др.). При перерезке или повреждении нервных волокон свойства иннервируемых ими клеток изменяются (это касается как физико-химических свойств поверхностной мембраны, так и биохимических процессов в протоплазме), что, в свою очередь, сопровождается глубокими нарушениями в состоянии органов и тканей (например, трофическими язвами). Если иннервация восстанавливается (в связи с регенерацией нервных волокон), то указанные нарушения могут исчезнуть.
Эволюция кровеносной системы У многих беспозвоночных животных сосудистой системы как таковой нет. В частности, у губок, кишечнополостных и плоских червей транспорт питательных веществ и кислорода в разные части тела осуществляется путем диффузии их тканевых жидкостей. Однако у организмов других групп появляются сосуды, обеспечивающие циркуляцию тканевой жидкости в определенных направлениях. В начале развиваются примитивные сосуды, но в результате развития в сосудах мышечной ткани они в дальнейшем оказываются способными к сокращению. Параллельно развивается в кровь жидкость, заполняющая сосуды.
Кровеносная система впервые встречается (развивается) у кольчатых червей, у которых она является замкнутой, представляя собой систему, состоящую из верхнего спинного и нижнего брюшного сосудов, объединенных кольцевыми сосудами, опоясывающими кишечник. Кроме того, от спинного и брюшного сосудов отходят более мелкие сосуды к стенкам тела. К головному отделу кровь идет по спинной стороне, а к заднему -- по брюшной.
У членистоногих, кровеносная система не замкнута, т. к. сосуды открываются в полость тела. Спинной сосуд разделяется перегородками (клапанами) на отдельные камеры -- сердца, сокращения которых заставляют проходить кровь в артерии, а из последних -- в пространства между органами. Из этих пространств кровь затем поступает в околосердечную полость.
У моллюсков кровеносная система тоже не замкнута и представлена сердцем, состоящим из нескольких предсердий и желудочка, а также артериальными и венозными сосудами. Вены впадают в предсердие, тогда как артерии отходят от желудочка.
Кровь у беспозвоночных несет кислород к тканям, удаляет двуокись углерода и транспортирует питательные вещества, гормоны, а также шлаковые продукты азотистого обмена. Кровь беспозвоночных по сравнению с кровью позвоночных содержит меньшее количество белков и форменных элементов. Единственным форменным элементом крови большинства беспозвоночных являются лейкоциты. Поэтому очень простой состав крови у беспозвоночных компенсируется разными способами транспорта кислорода, при которых используются гемоглобин, гемоцианин или другие дыхательные пигменты.
Предполагают, что развитие гемоглобина беспозвоночных шло независимо от развития гемоглобина позвоночных, поскольку этот пигмент у них обладает несколько отличной функцией. В частности, у беспозвоночных он выполняет запасательную функцию, обеспечивая защиту против недостатка кислорода в неблагоприятной среде.
По мере усложнения организации живых существ отмечается повышение количества кислорода в крови, т. е. повышается кислородная способность дыхательных пигментов.
Считают, что гемоглобин имеет древний возраст. Поскольку некоторые бактерии синтезируют гемопротеин, содержащий участки с аминокислотным составом, близким к последовательностям глобинов высших эукариотов, то предполагают, что гемоглобин впервые возник еще у прокариот. Однако не исключено, что у эукариот и прокариот имела место конвергентная эволюция.
Гемоглобин встречается у плоских, круглых и кольчатых червей, членистоногих, моллюсков, иглокожих, рыб, земноводных, рептилий, птиц, млекопитающих. Молекулярная масса гемоглобина беспозвоночных колеблется в пределах 17 000-2 750 000. У многих членистоногих и моллюсков встречается гемопианин, представляющий собой медьсодержащий белок молекулярной массой 400 000--5 000 000. Дыхательный пигмент обеспечивает у беспозвоночных буферную способность крови. Кроме того он ответственен и за осмотический баланс кровяной жидкости. Большинство насекомых не содержит совсем дыхательного пигмента, имея в крови растворенный кислород.
У полухордовых кровеносные сосуды еще не выстланы эндотелием. Сердце урохордовых представляет собой мускульное выпячивание около желудка. У хордовых кровеносная система характеризуется дальнейшим совершенствованием. У них развиваются сердце, сосуды, кровь.
У ланцетника кровеносная система является замкнутой, но сердца еще нет; его функцию выполняет передний отдел крупного сосуда в виде брюшной аорты. Впервые сердце появляется у водных позвоночных. В частности, у рыб сердце двухкамерное с предсердием и желудочком.
Круг кровообращения один, в котором артериальная и венозная кровь не смешиваются. Из сердца венозная кровь идет к жабрам, где, окисляясь, она становится артериальной, после чего расходится по артериям ко всем частям тела. К сердцу кровь вновь доставляется венами. У пресмыкающихся желудочек разделен неполной перегородкой на правую (венозную) и левую (артериальную) половины. У крокодилов желудочек разделен полностью. Однако у пресмыкающихся разделения артериального и венозного тока крови еще не происходит.
У наземных позвоночных развивается трех-, а затем четырехкамерное сердце, что явилось результатом очень крупного ароморфоза. У птиц и млекопитающих сердце построено из мышц и разделено на четыре камеры в виде двух предсердий и двух желудочков Имеются два круга кровообращения, один из которых большой, второй малый. Благодаря этим кругам артериальная и венозная кровь не смешиваются. Артериальная кровь выходит из сердца, венозная вступает в сердце.
Сердце позвоночных относят к миогенному типу, поскольку его сокращения начинаются внутри за счет сократительных элементов, называемых миофибриллами. У рыб, земноводных и пресмыкающихся сокращения сердца начинаются в тонкой мышечной стенке венозного синуса и продолжаются посредством внутренней проводящей системы через предсердие, потом к желудочкам. У птиц и млекопитающих венозного синуса нет.
У позвоночных усложняются строение и функции крови. Кровь состоит из форменных элементов и плазмы, увеличивается их масса. Например, в крови млекопитающих клеточная масса крови составляет 45% , плазменная -- 55% . Гемоглобин содержится в эритроцитах, которые у всех позвоночных, кроме млекопитающих, содержат ядра. У земноводных клетки крови очень крупные по сравнению с пресмыкающимися, птицами и млекопитающими. В 1 мл крови человека содержится 5 млн эритроцитов и 8--10 тыс лейкоцитов. Группы крови современных людей также являются результатом их эволюции В плазме крови содержатся ферменты, гормоны и другие соединения, необходимые для метаболизма клеток.
В ходе эволюции у высших животных взаимосвязи обмена веществ разных органов, обеспечиваемые кровью, получили дальнейшее усложнение. Одна из важнейших функций крови заключается в транспорте кислорода от легких к тканям и в транспорте СО2, являющимся конечным продуктом дыхания, от тканей к легким, причем эта функция крови развивалась в направлении способности транспортировать большие объемы кислорода. Основная часть кислорода, потребляемого организмом человека, транспортируется гемоглобином эритроцитов, масса которого составляет примерно треть массы эритроцитов. Гемоглобин представляет собой очень сложный белок, состоящий из двух а-полипептидных цепей и двух -полипептидных цепей, каждый из которых присоединяет по одной гемовой группе. Его происхождение является очень древним
Эритроциты участвуют также в транспорте СО2 из тканей в легкие, откуда он выводится в процессе выдыхания. Эволюционным приобретением млекопитающих, в том числе человека, является то, что их гемоглобин приспособлен как для транспорта кислорода, так и для транспорта СОд, причем оба эти транспорта взаимоусиливаемы.
Кровь ответственна и за перенос питательных веществ из тонкого кишечника в печень и другие органы, а также за выведение шлаков из тканей в почки. Эти функции тоже развивались в направлении увеличения количественной способности в связи с увеличивающейся интенсификацией метаболизма по мере усложнения организации живых существ.
Диффузия кислорода, двуокиси углерода, питательных веществ и метаболитов происходит лишь в капиллярах.
Параллельно с развитием кровеносной системы шло развитие лимфатической системы, которая впервые появилась у хордовых. Эта система в ходе эволюции обособилась из венозной системы.
Лимфатическая система состоит из лимфатических сосудов, лимфатических узлов и лимфы.
Лимфатические сосуды построены из эластичной и мышечной тканей и выстланы внутри эндотелием. Для них характерен такой же внутренний объем, как и у кровеносных капилляров. Лимфатические сосуды очень разветвлены. Они в виде очень тонких капилляров начинаются с межклеточных пространств, формируя затем более крупные сосуды. С помощью лимфатических сосудов лимфа проходит от тканей в венозное русло. Все лимфатические сосуды сливаются в один общий сборный сосуд (млечный путь). Одиночные лимфатические узлы обнаруживаются в толще слизистой оболочки и подслизистой основы кишечника у рыб, земноводных, пресмыкающихся, птиц и млекопитающих, но групповые лимфатические узлы развиваются лишь у млекопитающих.
У позвоночных по мере усложнения их организации наступает приближение лимфатических капилляров к клеткам и лимфоид-ным узелкам. Например, расстояние между лимфатическими капиллярами, кишечными эпителиоцитами и лимфоидными узелками у рыб составляет 150-180 мкм, но затем прогрессивно уменьшается у земноводных, пресмыкающихся и птиц, составляя у млекопитающих всего лишь 50-30 мкм.
В ходе эволюции многоклеточных животных кровеносная система сформировалась на месте рудиментов первичной полости тела, вытесненной у высших животных вторичной полостью тела, или целомом. Кровеносная система приняла на себя в организме высших животных универсальную транспортную роль передвижение питательных веществ, усвоенных органами пищеварения, и кислорода от органов дыхания ко всем органам тела, перенос продуктов метаболизма к органам выделения, углекислоты — к органам дыхания, доставка гормонов от эндокринных желез к органам, где данные гормоны будут действовать. В силу этого кровеносная система стала и одной из важнейших интегрирующих систем организма, обеспечивающих его целостность. Кровеносная система имеется у немертин кольчатых червей, моллюсков, членистоногих. иглокожих, полухордовых и хордовых. Ее строение у животных разных типов сильно варьирует. Два основных продольных сосуда — спинной и брюшной, тянущиеся соответственно над и под кишечником, связаны друг с другом поперечными сосудами; последние кольцеобразно охватывают кишечник и дают ответвления к покровам и различным внутренним органам. У животных всех типов, условно объединяемых названием «беспозвоночные» кровь по брюшному сосуду течет назад, а по спинному вперед. У хордовых наоборот: по брюшному сосуду кровь течет вперед, а по спинному назад. Пульсирующей функцией, обеспечивающей движение крови, у хордовых обладает брюшной сосуд или его производное — сердце, у беспозвоночных эту функцию несет спинной сосуд. У общих предков вторичноротых животных (к ним принадлежат полухордовые, иглокожие и хордовые) кровеносная система имела очень примитивное строение, представляя собой систему лакун, т. е. полостей без эпителизированных стенок, где направление кровотока еще не установилось. В дальнейшем совершенствование кровеносной системы происходило независимо и по-разному в различных ветвях ствола вторичноротых. Поэтому состояние кровеносной системы хордовых сформировалось особым путем и не может быть выведено из развитой кровеносной системы беспозвоночных — как вторичноротых, так и (тем более) первичноротых, к которым принадлежат кольчатые черви, членистоногие, моллюски и др.
|
Основные сокращения |
|
|
На рисунках красным цветом указана артериальная кровь, синим цветом — венозная; ак — артериальный конус; ба — брюшная аорта; бар — бедренная артерия; бв — бедренная вена; вжа — выносящая жаберная артерия; вп — воротная система почек; вс — венозный синус; всп — воротная система печени; вяв — верхняя яремная вена; ж — желудочек сердца; жа — жаберная артерия; зкв — задняя кардинальная вена; зпв — задняя полая вена; ка — кожная артерия; кв — кожная вена; кла — кожно-легочная артерия; Кп — Кювьеров проток; кр — корень аорты; ла — легочная артерия; лв — легочная вена; лда — левая дуга аорты; лж — левый желудочек; лк — луковица аорты; |
лп — левое предсердие; няв — нижняя яремная вена; оса — общий ствол дуг аорты; ола — общий ствол легочных артерий; ояв — общая яремная вена; пв — подкишечная вена; пда — правая дуга аорты; пж — правый желудочек; пжа — приносящая жаберная артерия; пка — подключичная артерия; пкв — передняя кардинальная вена; пкл — подключичная вена; пп — правое предсердие; ппв — передняя полая вена; пр — предсердие; пчв — печеночная вена; поа — почечная артерия; пов — почечная вена; са — сонная артерия; сп — сонный проток; спа — спинная аорта; ха — хвостовая артерия; хв — хвостовая вена; ча — чревная артерия |
Среди хордовых
наиболее примитивной кровеносной
системой обладают животные подтипа
бесчерепных.
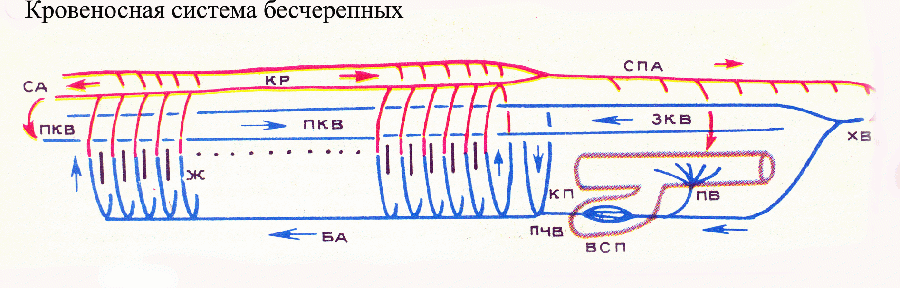
У типичного
представителя бесчерепных — ланцетника
— под глоткой тянется продольный сосуд
— брюшная аорта, играющая роль основного
пульсирующего органа. По брюшной аорте
кровь течет вперед. От этого магистрального
сосуда начинаются многочисленные (около
100 пар) жаберные артерии, которые
расположены в перегородках между
жаберными щелями, пронизывающими боковые
стенки глотки. Основания жаберных
артерий пульсируют, давая добавочный
импульс току крови. Через жаберные щели
непрерывно процеживается вода, и здесь
происходит газообмен между кровью и
водой: кровь в жаберных артериях
обогащается кислородом и отдает
углекислый газ, т. е. становится
артериальной. Обогащенную кислородом
кровь жаберные артерии доставляют на
спинную сторону глотки, где они впадают
в пару продольных сосудов — корни
спинной аорты. Впереди корни аорты
переходят в сонные артерии, несущие
арте кровь в головной конец тела, корни
аорты впадают в продольный спинной
сосуд — спинную аорту, по которой
артериальная кровь течет назад. От
спинной аорты отходят артерии к различным
внутренним органам и к кожным покровам,
где артерии распадаются на капилляры.
Из хвостовой области тела кровь собирается
в хвостовую вену, впереди впадающую в
подкишечную вену. Последняя собирает
кровь от кишечника, а затем в печеночном
выросте кишки вновь распадается на
капилляры, образующие воротную систему
печени. Из нее печеночная вена несет
кровь в брюшную аорту. У ланцетника
имеются еще две пары магистральных вен:
передние и задние кардинальные, тянущиеся
от переднего и заднего концов тела к
средней его части, где они впадают в
пару коротких Кювьеровых протоков.
Последние несут кровь в задний конец
брюшной аорты. Таким образом, брюшная
аорта получает обедненную кислородом
(венозную) кровь. У второго подтипа
хордовых — оболочников — кровеносная
система подверглась редукции. Как и
целый ряд других органов, в связи с
переходом к неподвижному (прикрепленному)
образу жизни на морском дне. Освоение
такого образа жизни в эволюции обычно
ведет к морфо-физиологическому регрессу.
У типичных представителей оболочников
асцидий — под желудком располагается
трубкообразное сердце, от которого
отходят два магистральных сосуда:
спереди — жаберная артерия, тянущаяся
по брюшной стороне глотки и отдающая
ветки к перегородкам между жаберными
отверстиями, позади — кишечная артерия,
которая разветвляется на сосуды, идущие
к внутренним органам. В конечном итоге
артерии впадают в лакуны, т. е. кровеносная
система оболочников не замкнута. Одна
из таких лакун — жаберно-кишечный синус
— лежит над глоткой и собирает кровь
из жаберной области. Интересно, что
направление пульсации сердца у оболочников
периодически меняется: сердце попеременно
гонит кровь то вперед, в жаберную артерию,
то назад, в кишечную. Некоторые ученые
видят в этом примитивную черту — еще
не установившееся направление кровотока.
Уникальной особенностью оболочников
является их кровяной пигмент —
гемованадин, в котором вместо ионов
железа (как в гемоглобине остальных
хордовых) или меди (как в гемоцианине
беспозвоночных ряда групп) используются
ионы ванадия. Эволюция третьего подтипа
хордовых — позвоночных — в целом шла
в направлении морфо-физиологического
прогресса, что, в частности, ярко
проявилось в последовательном
совершенствовании кровеносной системы:
ее дифференциации, интеграции,
интенсификации функций в ходе филогенеза
позвоночных. У всех позвоночных
центральным пропульсаторным органом
является сердце. У
миног,
представителей наиболее примитивного
класса современных позвоночных —
круглоротых, сердце имеет три отдела
(камеры):
 венозный синус (венозная пазуха),
предсердие и желудочек . От желудочка
отходит брюшная аорта, начало которой
расширено (луковица аорты). Впереди
брюшная аорта разделяется на пару
поджаберных артерий. От брюшной аорты
и поджаберных артерий начинаются 8 пар
приносящих жаберных артерий, распадающихся
в жабрах на капилляры (этого не было у
бесчерепных). От жабер обогащенная
кислородом кровь поступает в выносящие
жаберные артерии, которые впадают в
спинную аорту. В целом схема кровеносной
системы круглоротых сходна с таковой
бесчерепных, но отличается наличием
еще одной магистральной вены (нижней
яремной), выносящей кровь из вентральной
части головы, и отсутствием Кювьеровых
протоков (специфическая черта круглоротых).
Обе пары кардинальных вен, печеночная
и нижняя яремная вены впадают
непосредственно в венозную пазуху. Еще
одной специфической особенностью
кровеносной системы круглоротых,
является ее неполная замкнутость:
жаберные мешки окружены околожаберными
синусами, заполненными кровью и
представляющими собой кровеносные
лакуны. В связи с этим активное участие
в циркуляции крови принимают мышцы
жаберного аппарата, сокращение которых
выталкивает кровь из лакун и сосудов
жаберной области. У другой группы
круглоротых — миксин — работе сердца
помогают также пульсирующие участки
ряда вен (кардинальных, хвостовой и
воротной вены печени). У всех остальных
позвоночных — челюсторотых — по
сравнению с круглоротыми был достигнут
существенный прогресс в организации
кровеносной системы. Она стала полностью
замкнутой, сердце полностью приняло на
себя роль центрального пульсирующего
органа. Это сделало возможным повышение
кровяного давления и интенсификацию
кровотока, что способствовало общему
повышению уровня жизнедеятельности. У
хрящевых рыб (акулы, скаты, химеры)
сердце состоит из 4 камер:
венозный синус (венозная пазуха),
предсердие и желудочек . От желудочка
отходит брюшная аорта, начало которой
расширено (луковица аорты). Впереди
брюшная аорта разделяется на пару
поджаберных артерий. От брюшной аорты
и поджаберных артерий начинаются 8 пар
приносящих жаберных артерий, распадающихся
в жабрах на капилляры (этого не было у
бесчерепных). От жабер обогащенная
кислородом кровь поступает в выносящие
жаберные артерии, которые впадают в
спинную аорту. В целом схема кровеносной
системы круглоротых сходна с таковой
бесчерепных, но отличается наличием
еще одной магистральной вены (нижней
яремной), выносящей кровь из вентральной
части головы, и отсутствием Кювьеровых
протоков (специфическая черта круглоротых).
Обе пары кардинальных вен, печеночная
и нижняя яремная вены впадают
непосредственно в венозную пазуху. Еще
одной специфической особенностью
кровеносной системы круглоротых,
является ее неполная замкнутость:
жаберные мешки окружены околожаберными
синусами, заполненными кровью и
представляющими собой кровеносные
лакуны. В связи с этим активное участие
в циркуляции крови принимают мышцы
жаберного аппарата, сокращение которых
выталкивает кровь из лакун и сосудов
жаберной области. У другой группы
круглоротых — миксин — работе сердца
помогают также пульсирующие участки
ряда вен (кардинальных, хвостовой и
воротной вены печени). У всех остальных
позвоночных — челюсторотых — по
сравнению с круглоротыми был достигнут
существенный прогресс в организации
кровеносной системы. Она стала полностью
замкнутой, сердце полностью приняло на
себя роль центрального пульсирующего
органа. Это сделало возможным повышение
кровяного давления и интенсификацию
кровотока, что способствовало общему
повышению уровня жизнедеятельности. У
хрящевых рыб (акулы, скаты, химеры)
сердце состоит из 4 камер:
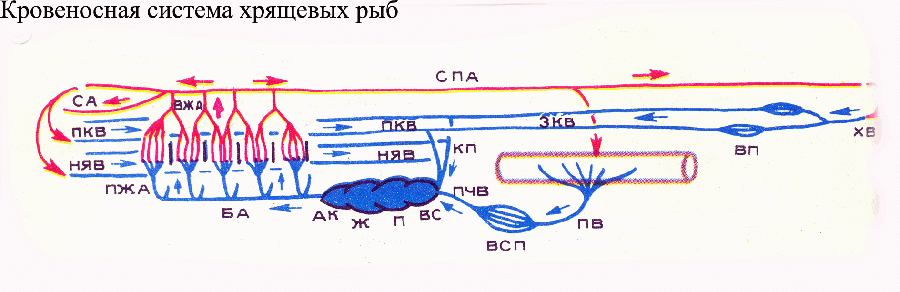 к З камерам, известным нам по круглоротым,
добавился еще артериальный конус.
Функция его — распределение крови между
передними и задними приносящими жаберными
артериями. Парные поджаберные сосуды,
имевшиеся у круглоротых, редуцированы.
У большинства хрящевых рыб жабры
сохраняются лишь на 4 жаберных дугах и
на подъязычной дуге висцерального
аппарата; соответственно, имеется 5 пар
приносящих жаберных артерий. Как и у
бесчерепных, магистральные вены впадают
не прямо в сердце, а в пару Кювьеровых
протоков. В отличие от круглоротых, у
хрящевых рыб нижняя яремная вена парная,
а на пути крови из хвостовой области
имеется еще одна капиллярная сеть — в
почках (воротная система почек). Класс
костных рыб объединяет ряд филогенетически
рано обособившихся друг от друга групп,
у которых кровеносные системы существенно
различаются по строению. У наиболее
распространенных и много численных
костных рыб (к ним относится большинство
наших обычных пресноводных рыб подвергся
редукции артериальный конус сердца, и
оно стало трехкамерным, как у круглоротых.
В начале брюшной аорты имеется расширение
(луковица аорты). Развиты лишь 4 пары
приносящих жаберных артерий. Воротная
система левой почки редуцирована.
Наземные
позвоночные возникли от древних
кистеперых рыб,
обитавших в пресных мелководных, богатых
растительностью тропических водоемах,
отчасти заболоченных. В таких водоемах
часто возникал дефицит растворенного
в воде кислорода (в связи с усиленным
гниением отмиравших растений). Важным
приспособлением к жизни в этих условиях
было развитие легких, позволявших при
дефиците кислорода в воде дышать
атмосферным воздухом. Среди современных
костных рыб близкий образ жизни ведут
родственные кистеперым рыбам двоякодышащие,
тогда как единственный доживший до
наших дней вид кистеперых — латимерия
— обитает в морских глубинах и легкие
для дыхания не использует.
Морфо-физиологической особенностью
рыб, дышащих и жабрами, и легкими (древние
кистеперые и современные двоякодышащие),
является решение проблемы рационального
использования обоих органов газообмена.
Дело в том, что при нормальном для рыб
жаберном дыхании обогащенная кислородом
кровь из передних жабер поступает в
голову, тогда как из задних — в спинную
аорту, несущую кровь назад по всему
телу. Легкие сформировались позади
глотки, т. е. позади самых задних жабер.
И поэтому если кровь из легких, как из
жабер, направить прямо в корни аорты,
то эта артериальная кровь поступит в
спинную аорту, а органы головы (и, в
частности, головной мозг, особенно
нуждающийся в постоянном поступлении
кислорода) не смогут ее использовать.
Поэтому возникает необходимость нового
перераспределения крови. Эта проблема
была решена возвращением артериальной
крови из легких в сердце (т. е. образованием
дополнительного легочного, или малого,
круга кровообращения) и формированием
в сердце особых приспособлений для
перераспределения крови. У
двоякодышащих
к З камерам, известным нам по круглоротым,
добавился еще артериальный конус.
Функция его — распределение крови между
передними и задними приносящими жаберными
артериями. Парные поджаберные сосуды,
имевшиеся у круглоротых, редуцированы.
У большинства хрящевых рыб жабры
сохраняются лишь на 4 жаберных дугах и
на подъязычной дуге висцерального
аппарата; соответственно, имеется 5 пар
приносящих жаберных артерий. Как и у
бесчерепных, магистральные вены впадают
не прямо в сердце, а в пару Кювьеровых
протоков. В отличие от круглоротых, у
хрящевых рыб нижняя яремная вена парная,
а на пути крови из хвостовой области
имеется еще одна капиллярная сеть — в
почках (воротная система почек). Класс
костных рыб объединяет ряд филогенетически
рано обособившихся друг от друга групп,
у которых кровеносные системы существенно
различаются по строению. У наиболее
распространенных и много численных
костных рыб (к ним относится большинство
наших обычных пресноводных рыб подвергся
редукции артериальный конус сердца, и
оно стало трехкамерным, как у круглоротых.
В начале брюшной аорты имеется расширение
(луковица аорты). Развиты лишь 4 пары
приносящих жаберных артерий. Воротная
система левой почки редуцирована.
Наземные
позвоночные возникли от древних
кистеперых рыб,
обитавших в пресных мелководных, богатых
растительностью тропических водоемах,
отчасти заболоченных. В таких водоемах
часто возникал дефицит растворенного
в воде кислорода (в связи с усиленным
гниением отмиравших растений). Важным
приспособлением к жизни в этих условиях
было развитие легких, позволявших при
дефиците кислорода в воде дышать
атмосферным воздухом. Среди современных
костных рыб близкий образ жизни ведут
родственные кистеперым рыбам двоякодышащие,
тогда как единственный доживший до
наших дней вид кистеперых — латимерия
— обитает в морских глубинах и легкие
для дыхания не использует.
Морфо-физиологической особенностью
рыб, дышащих и жабрами, и легкими (древние
кистеперые и современные двоякодышащие),
является решение проблемы рационального
использования обоих органов газообмена.
Дело в том, что при нормальном для рыб
жаберном дыхании обогащенная кислородом
кровь из передних жабер поступает в
голову, тогда как из задних — в спинную
аорту, несущую кровь назад по всему
телу. Легкие сформировались позади
глотки, т. е. позади самых задних жабер.
И поэтому если кровь из легких, как из
жабер, направить прямо в корни аорты,
то эта артериальная кровь поступит в
спинную аорту, а органы головы (и, в
частности, головной мозг, особенно
нуждающийся в постоянном поступлении
кислорода) не смогут ее использовать.
Поэтому возникает необходимость нового
перераспределения крови. Эта проблема
была решена возвращением артериальной
крови из легких в сердце (т. е. образованием
дополнительного легочного, или малого,
круга кровообращения) и формированием
в сердце особых приспособлений для
перераспределения крови. У
двоякодышащих
 сердце состоит из венозного синуса
(получающего, как обычно у рыб, венозную
кровь), предсердия, желудочка и
артериального конуса. Венозный синус
впадает в правую половину предсердия,
тогда как в левую его половину впадают
легочные вены, приносящие артериальную
кровь из легких. В предсердии, желудочке
и артериальном конусе имеются неполные
внутренние перегородки (в частности,
спиральный клапан артериального конуса),
позволяющие разделить кровь, поступившую
из левой и правой половин предсердия.
Из левой половины предсердия артериальная
кровь поступает в нижнюю часть сильно
укороченной брюшной аорты и по ней — к
двум передним парам приносящих жаберных
артерий; затем эта кровь поступает по
сонным артериям в голову. Из правой
половины предсердия венозная кровь
направляется в верхнюю часть брюшной
аорты и по ней — к двум задним (ближайшим
к сердцу) парам жаберных артерий.
Подчеркнем, что легочные артерии
начинаются от четвертой (самой задней)
пары выносящих жаберных артерий. Механизм
распределения крови в сердце двоякодышащих
рыб лабилен и позволяет направлять
больше или меньше крови в передние или
задние жаберные артерии в зависимости
от физиологического состояния рыбы и
от того, какой орган (легкие или жабры)
в данный момент работают интенсивнее.
В венозной системе у двоякодышащих рыб
редуцирована правая задняя полая вена,
зато появился новый магистральный
венозный ствол — задняя полая вена,
несущая кровь из воротной системы почек
прямо в венозный синус, минуя Кювьеровы
протоки. У наземных позвоночных (тетрапод)
произошли значительные изменения в
строении кровеносной системы по сравнению
с рыбами (хотя общее направление этих
перестроек намечено уже у двоякодышащих).
У всех тетрапод брюшная аорта отчасти
редуцирована, отчасти разделена на
несколько обособленных артериальных
стволов, начинающихся от сердца, причем
это разделение произошло по-разному у
предков современных земноводных,
пресмыкающихся и млекопитающих. У
современных земноводных
сердце состоит из венозного синуса
(получающего, как обычно у рыб, венозную
кровь), предсердия, желудочка и
артериального конуса. Венозный синус
впадает в правую половину предсердия,
тогда как в левую его половину впадают
легочные вены, приносящие артериальную
кровь из легких. В предсердии, желудочке
и артериальном конусе имеются неполные
внутренние перегородки (в частности,
спиральный клапан артериального конуса),
позволяющие разделить кровь, поступившую
из левой и правой половин предсердия.
Из левой половины предсердия артериальная
кровь поступает в нижнюю часть сильно
укороченной брюшной аорты и по ней — к
двум передним парам приносящих жаберных
артерий; затем эта кровь поступает по
сонным артериям в голову. Из правой
половины предсердия венозная кровь
направляется в верхнюю часть брюшной
аорты и по ней — к двум задним (ближайшим
к сердцу) парам жаберных артерий.
Подчеркнем, что легочные артерии
начинаются от четвертой (самой задней)
пары выносящих жаберных артерий. Механизм
распределения крови в сердце двоякодышащих
рыб лабилен и позволяет направлять
больше или меньше крови в передние или
задние жаберные артерии в зависимости
от физиологического состояния рыбы и
от того, какой орган (легкие или жабры)
в данный момент работают интенсивнее.
В венозной системе у двоякодышащих рыб
редуцирована правая задняя полая вена,
зато появился новый магистральный
венозный ствол — задняя полая вена,
несущая кровь из воротной системы почек
прямо в венозный синус, минуя Кювьеровы
протоки. У наземных позвоночных (тетрапод)
произошли значительные изменения в
строении кровеносной системы по сравнению
с рыбами (хотя общее направление этих
перестроек намечено уже у двоякодышащих).
У всех тетрапод брюшная аорта отчасти
редуцирована, отчасти разделена на
несколько обособленных артериальных
стволов, начинающихся от сердца, причем
это разделение произошло по-разному у
предков современных земноводных,
пресмыкающихся и млекопитающих. У
современных земноводных
 от артериального
конуса сердца начинаются три пары
сосудов: сонные артерии, дуги аорты
(которые на спинной стороне сливаются,
образуя спинную аорту) и легочные (а у
бесхвостых амфибий — кожно-легочные)
артерии. Первые из них соответствуют 1
паре жаберных артерий с собственно
сонными артериями рыб; вторые — II паре
жаберных артерий с корнями аорты рыб;
третьи — IV паре жаберных артерий с
легочными артериями рыб (у хвостатых
амфибий иногда сохраняются две пары
дуг аорты, соответствующие II и III парам
жаберных артерий). У рыб все эти сосуды
связаны друг с другом над глоткой корнями
аорты; такая связь сохраняется и у
некоторых тетрапод в виде сонных протоков
между сонными артериями и дугами аорты,
а также Боталловых протоков между дугами
аорты и легочными артериями. По сонным
артериям кровь направляется к голове;
по дугам аорты — в спинную аорту и по
ней к различным органам тела; легочные
(или кожно-легочные) артерии обеспечивают
кровоснабжение органов дыхания. У
земноводных несовершенство механизма
вентиляции легких не позволяет этим
животным ограничиться легочным дыханием,
и кожа у них является важным дополнительным
органом дыхания. У хвостатых и безногих
амфибий, легочные артерии не объединены
с кожными. Кровоснабжение кожных
покровов, как и у других позвоночных,
осуществляется ветвями спинной аорты.
Венозная система земноводных отличается
от таковой рыб редукцией задних
кардинальных вен, которые замещаются
задней полой веной, получающей кровь
из воротных систем почек и печени и
доставляющей ее в венозную пазуху
сердца. Передние кардинальные вены у
наземных позвоночных сохраняются, но
получают новое название: их традиционно
именуют верхними яремными венами.
Кювьеровы протоки у тетрапод называют
передними полыми венами. Сердце
земноводных состоит из пяти отделов:
венозного синуса (пазухи), левого и
правого предсердий, желудочка и
артериального конуса. В венозный синус
впадают передние и задняя полые вены.
В левое предсердие впадают легочные
вены, приносящие артериальную кровь из
легких. В желудочке кровь из левого и
правого предсердий в определенной
степени смешивается. Нередко это
упрощенно объясняют примитивностью и
несовершенством сердца земноводных,
но в действительности дело обстоит
сложнее и интереснее. Объединение в
желудочке сердца земноводных крови,
поступающей из левого и правого
предсердий, имеет важное функциональное
значение: если бы такого смешивания
крови не было, земноводным не удалось
бы эффективно использовать кожное
дыхание! Артериальная кровь от легких
через легочные вены попадает в левое
предсердие, тогда как от кожи — через
полые вены в венозный синус и затем в
правое предсердие. В жизни земноводных
нередки ситуации, когда этим животным
приходится использовать в основном
(или даже исключительно) кожное дыхание:
так происходит при нырянии и длительном
пребывании под водой. В этом случае
легкие уже не снабжают организм
кислородом, а сами его потребляют. Если
бы желудочек сердца земноводных был
разделен, артериальная кровь от кожи
поступала бы по легочным артериям в
легкие, а наиболее нуждающиеся в кислороде
органы головы оказались бы его лишенными.
Поэтому земноводным необходимо именно
такое сердце, имеющее механизм для
оптимального перераспределения потоков
крови в соответствии с потребностями
и физиологическим состоянием животных.
При преимущественно легочном дыхании,
напротив, целесообразно разделение
крови, поступающей в желудочек сердца
из разных предсердий. В полость желудочка
от его стенок выступают мышечные гребни,
которые частично разделяют ее на
небольшие отсеки. Это препятствует
смешиванию крови. В результате при
сокращении желудочка в артериальный
конус сначала поступает относительно
наименее обогащенная кислородом кровь
из правой части желудочка, затем смешанная
и, наконец, наиболее богатая кислородом
артериальная кровь из левой стороны
желудочка. При легочном дыхании первая
(относительно наиболее венозная) порция
крови направляется в ближайшие к сердцу
легочные (или кожно-легочные) артерии,
вторая — в дуги аорты и третья (наиболее
артериальная) — в сонные артерии. Если
же животное находится под водой и дышит
только кожей, несколько изменяется
соотношение фаз пульсации желудочка и
артериального конуса, в результате чего
в легочные артерии подается относительно
меньше крови, а в желудочке кровь из
левого и правого предсердий сильнее
смешивается, что позволяет, как было
указано выше, оптимально использовать
артериальную кровь, поступающую от
кожи. У
пресмыкающихся
в связи с усовершенствованием механизма
вентиляции легких (развитие мощного
легочного насоса — грудной клетки)
необходимость в кожном дыхании отпадает
и появляется принципиальная возможность
разделения артериального и венозного
потоков крови. Однако у большинства
рептилий перегородка в желудочке сердца
остается неполной, и кровь там может
смешиваться. Благодаря этой особенности
у пресмыкающихся сохраняется возможность
перераспределения крови в сердце и
регуляции поступления больших или
меньших ее количеств в разные сосуды.
А это очень важно, и вот почему. Большое
значение для пресмыкающихся имеет
особая форма терморегуляции, называемая
гелиотермией,
— повышение температуры тела до
оптимального уровня путем обогревания
в лучах солнца (инсоляции).
При инсоляции физиологически выгодно
направлять больше крови в периферические
сосуды (в частности, к кожным покровам)
и меньше — в легкие! где испарение влаги
приводит к некоторому понижению
температуры тела. Напротив, когда
животные, достигнув оптимальной
температуры тела, переходят к активной
деятельности (поиски пищи, перемещения
по территории и т. п.), необходимо больше
крови направлять к легким. В соответствии
с этими потребностями и регулируется
поток крови в малом и большом кругах
кровообращения. Брюшная аорта у предков
рептилий разделилась на три артериальных
ствола, начинающихся от сердца: левую
и правую дуги аорты (сливающиеся в
спинную аорту) и общий ствол легочных
артерий.
от артериального
конуса сердца начинаются три пары
сосудов: сонные артерии, дуги аорты
(которые на спинной стороне сливаются,
образуя спинную аорту) и легочные (а у
бесхвостых амфибий — кожно-легочные)
артерии. Первые из них соответствуют 1
паре жаберных артерий с собственно
сонными артериями рыб; вторые — II паре
жаберных артерий с корнями аорты рыб;
третьи — IV паре жаберных артерий с
легочными артериями рыб (у хвостатых
амфибий иногда сохраняются две пары
дуг аорты, соответствующие II и III парам
жаберных артерий). У рыб все эти сосуды
связаны друг с другом над глоткой корнями
аорты; такая связь сохраняется и у
некоторых тетрапод в виде сонных протоков
между сонными артериями и дугами аорты,
а также Боталловых протоков между дугами
аорты и легочными артериями. По сонным
артериям кровь направляется к голове;
по дугам аорты — в спинную аорту и по
ней к различным органам тела; легочные
(или кожно-легочные) артерии обеспечивают
кровоснабжение органов дыхания. У
земноводных несовершенство механизма
вентиляции легких не позволяет этим
животным ограничиться легочным дыханием,
и кожа у них является важным дополнительным
органом дыхания. У хвостатых и безногих
амфибий, легочные артерии не объединены
с кожными. Кровоснабжение кожных
покровов, как и у других позвоночных,
осуществляется ветвями спинной аорты.
Венозная система земноводных отличается
от таковой рыб редукцией задних
кардинальных вен, которые замещаются
задней полой веной, получающей кровь
из воротных систем почек и печени и
доставляющей ее в венозную пазуху
сердца. Передние кардинальные вены у
наземных позвоночных сохраняются, но
получают новое название: их традиционно
именуют верхними яремными венами.
Кювьеровы протоки у тетрапод называют
передними полыми венами. Сердце
земноводных состоит из пяти отделов:
венозного синуса (пазухи), левого и
правого предсердий, желудочка и
артериального конуса. В венозный синус
впадают передние и задняя полые вены.
В левое предсердие впадают легочные
вены, приносящие артериальную кровь из
легких. В желудочке кровь из левого и
правого предсердий в определенной
степени смешивается. Нередко это
упрощенно объясняют примитивностью и
несовершенством сердца земноводных,
но в действительности дело обстоит
сложнее и интереснее. Объединение в
желудочке сердца земноводных крови,
поступающей из левого и правого
предсердий, имеет важное функциональное
значение: если бы такого смешивания
крови не было, земноводным не удалось
бы эффективно использовать кожное
дыхание! Артериальная кровь от легких
через легочные вены попадает в левое
предсердие, тогда как от кожи — через
полые вены в венозный синус и затем в
правое предсердие. В жизни земноводных
нередки ситуации, когда этим животным
приходится использовать в основном
(или даже исключительно) кожное дыхание:
так происходит при нырянии и длительном
пребывании под водой. В этом случае
легкие уже не снабжают организм
кислородом, а сами его потребляют. Если
бы желудочек сердца земноводных был
разделен, артериальная кровь от кожи
поступала бы по легочным артериям в
легкие, а наиболее нуждающиеся в кислороде
органы головы оказались бы его лишенными.
Поэтому земноводным необходимо именно
такое сердце, имеющее механизм для
оптимального перераспределения потоков
крови в соответствии с потребностями
и физиологическим состоянием животных.
При преимущественно легочном дыхании,
напротив, целесообразно разделение
крови, поступающей в желудочек сердца
из разных предсердий. В полость желудочка
от его стенок выступают мышечные гребни,
которые частично разделяют ее на
небольшие отсеки. Это препятствует
смешиванию крови. В результате при
сокращении желудочка в артериальный
конус сначала поступает относительно
наименее обогащенная кислородом кровь
из правой части желудочка, затем смешанная
и, наконец, наиболее богатая кислородом
артериальная кровь из левой стороны
желудочка. При легочном дыхании первая
(относительно наиболее венозная) порция
крови направляется в ближайшие к сердцу
легочные (или кожно-легочные) артерии,
вторая — в дуги аорты и третья (наиболее
артериальная) — в сонные артерии. Если
же животное находится под водой и дышит
только кожей, несколько изменяется
соотношение фаз пульсации желудочка и
артериального конуса, в результате чего
в легочные артерии подается относительно
меньше крови, а в желудочке кровь из
левого и правого предсердий сильнее
смешивается, что позволяет, как было
указано выше, оптимально использовать
артериальную кровь, поступающую от
кожи. У
пресмыкающихся
в связи с усовершенствованием механизма
вентиляции легких (развитие мощного
легочного насоса — грудной клетки)
необходимость в кожном дыхании отпадает
и появляется принципиальная возможность
разделения артериального и венозного
потоков крови. Однако у большинства
рептилий перегородка в желудочке сердца
остается неполной, и кровь там может
смешиваться. Благодаря этой особенности
у пресмыкающихся сохраняется возможность
перераспределения крови в сердце и
регуляции поступления больших или
меньших ее количеств в разные сосуды.
А это очень важно, и вот почему. Большое
значение для пресмыкающихся имеет
особая форма терморегуляции, называемая
гелиотермией,
— повышение температуры тела до
оптимального уровня путем обогревания
в лучах солнца (инсоляции).
При инсоляции физиологически выгодно
направлять больше крови в периферические
сосуды (в частности, к кожным покровам)
и меньше — в легкие! где испарение влаги
приводит к некоторому понижению
температуры тела. Напротив, когда
животные, достигнув оптимальной
температуры тела, переходят к активной
деятельности (поиски пищи, перемещения
по территории и т. п.), необходимо больше
крови направлять к легким. В соответствии
с этими потребностями и регулируется
поток крови в малом и большом кругах
кровообращения. Брюшная аорта у предков
рептилий разделилась на три артериальных
ствола, начинающихся от сердца: левую
и правую дуги аорты (сливающиеся в
спинную аорту) и общий ствол легочных
артерий.
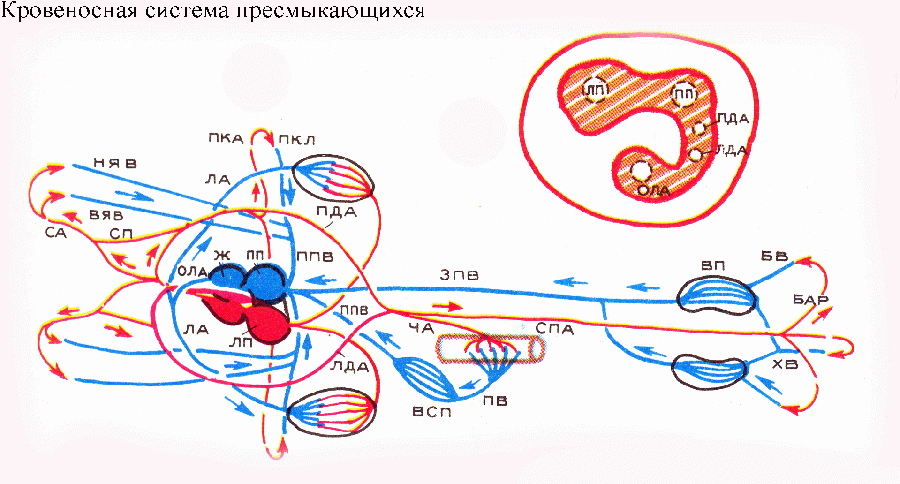 Правая дуга аорты
получает из желудочка сердца наиболее
чистую артериальную кровь; в связи с
этим сонные и подключичные артерии
(несущие кровь к голове и к передним
конечностям) у рептилий начинаются от
правой дуги аорты. Основные вены в
принципе соответствуют таковым амфибий.
Сердце у большинства пресмыкающихся
трехкамерное: два предсердия и желудочек.
Характерные для земноводных венозная
пазуха и артериальный конус редуцировались.
В левое предсердие впадают легочные
вены, в правое — три полые вены. В
желудочке имеется неполная перегородка
довольно сложной формы, расположенная
почти в горизонтальной плоскости и
разделяющая желудочек на спинной и
брюшной отделы. Оба предсердия впадают
в спинной отдел желудочка. Распределение
крови в желудочке происходит следующим
образом. Первым сокращается правое
предсердие, и венозная кровь из него
стекает в брюшной отдел желудочка, от
которого начинается общий ствол легочных
артерий. Затем сокращается левое
предсердие, артериальная кровь из
которого заполняет спинной отдел
желудочка. От этого отдела начинаются
обе дуги аорты, причем начало левой дуги
расположено правее и ниже начала правой
(затем, по ходу, дуги аорты перекрещиваются).
При систоле желудочка сердца его
перегородка полностью отделяет брюшной
отдел от спинного, так что в обе дуги
аорты поступает артериальная кровь, а
в легочные артерии — венозная. Если же
при инсоляции требуется направить
меньше крови в легочные артерии и больше
— в дуги аорты, просвет общего ствола
легочных артерий сужается (сокращением
гладкой мускулатуры в его стенках) и
часть венозной крови из брюшного отдела
желудочка возвращается в спинной. В
левую дугу аорты в этом случае попадает
смешанная кровь. Как было уже указано,
голова и передние конечности всегда
получают артериальную кровь через
правую дугу аорты. Нетрудно заметить,
что полное анатомическое разделение
желудочка сердца такой горизонтальной
перегородкой невозможно, так как при
этом легочные артерии оказались бы
изолированы от потока крови. Эту проблему
решили высшие
рептилии — крокодилы:
у их предков первоначальная горизонтальная
перегородка в желудочке сердца отчасти
редуцировалась, отчасти преобразовалась
в новую, вертикальную перегородку,
полностью разделившую желудочек на
левую и правую камеры. Причем от левого
желудочка начинается правая дуга аорты,
а от правого — левая дуга аорты и общий
ствол легочных артерий. Артериальные
сосуды, начинающиеся от правого желудочка,
получают из сердца венозную кровь.
Однако когда крокодилы находятся на
поверхности воды или на суше и нормально
дышат, по левой дуге аорты венозная
кровь проходит только до перекреста
двух дуг аорты, где между последними
имеется связь — так называемое Паниццево
отверстие. Благодаря более высокому
давлению в правой дуге аорты артериальная
кровь из нее через это отверстие поступает
и в левую дугу аорты. А при нырянии и при
инсоляции у крокодилов происходит
перераспределение крови в магистральных
артериях, как и у других пресмыкающихся:
сужается просвет легочных артерий,
повышается давление крови в левом
желудочке и левой дуге аорты и венозная
кровь начинает поступать по левой дуге
дальше Паниццева отверстия. Отметим,
что левая дуга у крокодилов не сливается
с правой, а продолжается в чревную
артерию, снабжающую кровью кишечник.
Спинную же аорту образует лишь правая
дуга. Предки птиц в филогенезе древних
рептилий обособились от группы
псевдозухий, от которых возникли и
крокодилы. Не случайно поэтому кровеносная
система птиц обнаруживает определенное
сходство с таковой крокодилов, как бы
продолжая намеченное у последних
направление эволюционных перестроек.
У птиц, как и у крокодилов, сердце
четырехкамерное, и желудочек разделен
полной вертикальной перегородкой на
левую и правую камеры. Птицы
приобрели гомойотермию
— способность поддерживать температуру
своего тела на постоянном уровне
«теплокровность». Поэтому для них отпала
необходимость в инсоляции и перераспределении
крови в сердце и магистральных артериях,
столь важном для пресмыкающихся. Левая
дуга аорты в этих условиях стала ненужной
и полностью редуцировалась. От сердца
птицы отходят два артериальных ствола:
Правая дуга аорты
получает из желудочка сердца наиболее
чистую артериальную кровь; в связи с
этим сонные и подключичные артерии
(несущие кровь к голове и к передним
конечностям) у рептилий начинаются от
правой дуги аорты. Основные вены в
принципе соответствуют таковым амфибий.
Сердце у большинства пресмыкающихся
трехкамерное: два предсердия и желудочек.
Характерные для земноводных венозная
пазуха и артериальный конус редуцировались.
В левое предсердие впадают легочные
вены, в правое — три полые вены. В
желудочке имеется неполная перегородка
довольно сложной формы, расположенная
почти в горизонтальной плоскости и
разделяющая желудочек на спинной и
брюшной отделы. Оба предсердия впадают
в спинной отдел желудочка. Распределение
крови в желудочке происходит следующим
образом. Первым сокращается правое
предсердие, и венозная кровь из него
стекает в брюшной отдел желудочка, от
которого начинается общий ствол легочных
артерий. Затем сокращается левое
предсердие, артериальная кровь из
которого заполняет спинной отдел
желудочка. От этого отдела начинаются
обе дуги аорты, причем начало левой дуги
расположено правее и ниже начала правой
(затем, по ходу, дуги аорты перекрещиваются).
При систоле желудочка сердца его
перегородка полностью отделяет брюшной
отдел от спинного, так что в обе дуги
аорты поступает артериальная кровь, а
в легочные артерии — венозная. Если же
при инсоляции требуется направить
меньше крови в легочные артерии и больше
— в дуги аорты, просвет общего ствола
легочных артерий сужается (сокращением
гладкой мускулатуры в его стенках) и
часть венозной крови из брюшного отдела
желудочка возвращается в спинной. В
левую дугу аорты в этом случае попадает
смешанная кровь. Как было уже указано,
голова и передние конечности всегда
получают артериальную кровь через
правую дугу аорты. Нетрудно заметить,
что полное анатомическое разделение
желудочка сердца такой горизонтальной
перегородкой невозможно, так как при
этом легочные артерии оказались бы
изолированы от потока крови. Эту проблему
решили высшие
рептилии — крокодилы:
у их предков первоначальная горизонтальная
перегородка в желудочке сердца отчасти
редуцировалась, отчасти преобразовалась
в новую, вертикальную перегородку,
полностью разделившую желудочек на
левую и правую камеры. Причем от левого
желудочка начинается правая дуга аорты,
а от правого — левая дуга аорты и общий
ствол легочных артерий. Артериальные
сосуды, начинающиеся от правого желудочка,
получают из сердца венозную кровь.
Однако когда крокодилы находятся на
поверхности воды или на суше и нормально
дышат, по левой дуге аорты венозная
кровь проходит только до перекреста
двух дуг аорты, где между последними
имеется связь — так называемое Паниццево
отверстие. Благодаря более высокому
давлению в правой дуге аорты артериальная
кровь из нее через это отверстие поступает
и в левую дугу аорты. А при нырянии и при
инсоляции у крокодилов происходит
перераспределение крови в магистральных
артериях, как и у других пресмыкающихся:
сужается просвет легочных артерий,
повышается давление крови в левом
желудочке и левой дуге аорты и венозная
кровь начинает поступать по левой дуге
дальше Паниццева отверстия. Отметим,
что левая дуга у крокодилов не сливается
с правой, а продолжается в чревную
артерию, снабжающую кровью кишечник.
Спинную же аорту образует лишь правая
дуга. Предки птиц в филогенезе древних
рептилий обособились от группы
псевдозухий, от которых возникли и
крокодилы. Не случайно поэтому кровеносная
система птиц обнаруживает определенное
сходство с таковой крокодилов, как бы
продолжая намеченное у последних
направление эволюционных перестроек.
У птиц, как и у крокодилов, сердце
четырехкамерное, и желудочек разделен
полной вертикальной перегородкой на
левую и правую камеры. Птицы
приобрели гомойотермию
— способность поддерживать температуру
своего тела на постоянном уровне
«теплокровность». Поэтому для них отпала
необходимость в инсоляции и перераспределении
крови в сердце и магистральных артериях,
столь важном для пресмыкающихся. Левая
дуга аорты в этих условиях стала ненужной
и полностью редуцировалась. От сердца
птицы отходят два артериальных ствола:
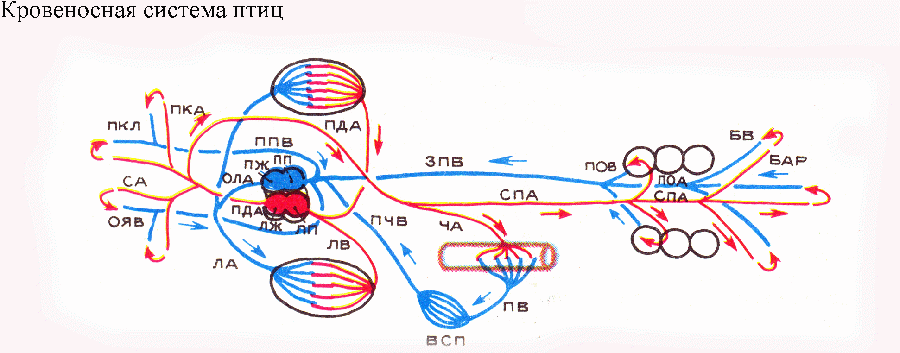 правая дуга аорты — от левого желудочка
(получает артериальную кровь) и общий
ствол легочных артерий — от правого
желудочка (получает венозную кровь).
Общая схема кровеносной системы птиц,
таким образом, упрощена по сравнению с
таковой рептилий. Морфо-физиологический
прогресс здесь достигнут не усложнением,
а упрощением и рационализацией системы.
Воротная система почек у птиц подверглась
редукции, и кровоснабжение почек
осуществляется почечными артериями,
отходящими от спинной аорты. У
предков млекопитающих
брюшная аорта разделилась не на три,
как у типичных пресмыкающихся, а на два
артериальных ствола: общий ствол дуг
аорты и общий ствол легочных артерий.
правая дуга аорты — от левого желудочка
(получает артериальную кровь) и общий
ствол легочных артерий — от правого
желудочка (получает венозную кровь).
Общая схема кровеносной системы птиц,
таким образом, упрощена по сравнению с
таковой рептилий. Морфо-физиологический
прогресс здесь достигнут не усложнением,
а упрощением и рационализацией системы.
Воротная система почек у птиц подверглась
редукции, и кровоснабжение почек
осуществляется почечными артериями,
отходящими от спинной аорты. У
предков млекопитающих
брюшная аорта разделилась не на три,
как у типичных пресмыкающихся, а на два
артериальных ствола: общий ствол дуг
аорты и общий ствол легочных артерий.
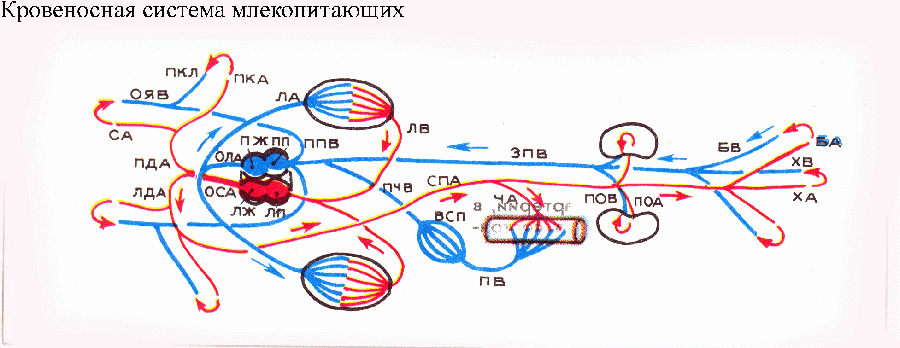 Это упростило задачу разделения крови
в сердце млекопитающих, которое стало
четырехкамерным, как у птиц. Желудочек
сердца разделился вертикальной
перегородкой на левый (от которого
начинается общий ствол дуг аорты) и
правый (откуда берет начало легочный
ствол). Впереди общий ствол дуг аорты
разделяется на левую дугу аорты
(продолжающуюся в спинную аорту) и
безымянную артерию, которая гомологична
редуцированной правой дуге аорты — она
не достигает спинной аорты и переходит
в правую подключичную артерию. Сонные
артерии начинаются симметрично: левая
— от левой дуги аорты, правая — от
безымянной артерии (у некоторых видов
млекопитающих обе сонные артерии
начинаются от общего ствола с правой
подключичной артерией, который называется
в этом случае плечеголовным). В венозной
системе млекопитающих, как и у птиц,
редуцировалась воротная система почек.
Возможно, в обоих случаях это связано
с повышением кровяного давления (по
сравнению с низшими позвоночными). При
видимом сходстве кровеносных систем
млекопитающих и птиц (четырехкамерное
сердце, единственная дуга аорты образует
спинную аорту, редукция воротной системы
почек) между ними имеются глубокие
различия. У птиц от левого желудочка
сердца начинается правая дуга аорты, а
левая утрачена, тогда как у млекопитающих
от левого желудочка начинается общий
ствол дуг аорты, левая дуга развита
полностью, а правая редуцирована. Эти
различия связаны с длительной независимой
эволюцией предков птиц и млекопитающих,
в ходе которой сходные прогрессивные
черты кровеносной системы были приобретены
разными путями. Вероятно, у общих предков
птиц и млекопитающих еще сохранялся
единый желудочек сердца и артериальный
конус, а также короткий общий артериальный
ствол — брюшная аорта. Разделение
желудочка сердца на камеры и брюшной
аорты на магистральные артерии происходило
самостоятельно и по-разному в
филогенетических стволах предковых
форм рептилий, ведущих к птицам и
млекопитающим. Развитие горизонтальной
внутрижелудочковой перегородки в сердце
типичных пресмыкающихся интересно как
пример инадаптивного пути эволюционных
преобразований. По В. О. Ковалевскому,
при инадаптивном развитии формирующееся
приспособление несет в себе предпосылки
для возникновения каких-либо внутренних
конструктивных дефектов, обнаруживающихся
не сразу, но постепенно нарастающих по
мере развития. Инадаптивная эволюция
является результатом своего рода
«неудачного» выбора направления
эволюционных изменений. А такой «неудачный
выбор» вполне вероятен: естественный
отбор благоприятствует любым изменениям,
которые дают какие-либо преимущества
организмам в борьбе за существование
в данный исторический момент, но не
может «предвидеть» ни результатов
дальнейшей эволюции в том или другом
направлении, ни новых форм взаимодействия
организма с внешней средой, которые
сложатся в будущем. На уровне организации
пресмыкающихся их неполная внутрижелудочковая
перегородка в целом отвечает физиологическим
потребностям этих животных, разделяя
артериальную и венозную кровь и в то же
время позволяя перераспределять потоки
крови в соответствии с характером
активности. Однако из-за своего положения
относительно устий предсердий и мест
отхождения артерий эта перегородка не
может полностью завершить разделение
желудочка сердца на артериальную и
венозную камеры. На базе такого устройства
сердца его дальнейшая прогрессивная
эволюция затруднена и требует довольно
сложных перестроек (произошедших у
крокодилов и птиц). В то же время на
первых этапах возникновения
внутрижелудочковой перегородки ее
вертикальное или горизонтальное
положение, так же как и разделение
брюшной аорты на три или на два
магистральных ствола, вероятно, не имели
существенных преимуществ друг перед
другом. Однако путь эволюционных
преобразований сердца у предков
млекопитающих был более перспективным
для полного разделения желудочка, что
обнаружилось на более поздних этапах
эволюции. В эволюции позвоночных, как
отмечалось выше, в целом происходило
общее усложнение и совершенствование
кровеносной системы. Однако не все
эволюционные перестройки кровеносной
системы представляют собой ароморфозы
(т. е. изменения на пути морфо-физиологического
прогресса, по А. Н. Северцову). Усложнение
организации само по себе еще не является
прогрессивным изменением. Иногда оно
служит своего рода компенсацией за
несовершенство какой-либо функции или
представляет собой результат развития
частного приспособления (идиоадаптации,
по А. Н. Северцову) к специфическому
образу жизни. Именно так следует оценивать
появление второго (малого) круга
кровообращения и усложнение строения
сердца у двоякодышащих рыб. Лишь в тех
случаях, когда усложнение строения
связано с интенсификацией функций, с
повышением уровня жизнедеятельности
(выражающимся в интенсификации процессов
обмена веществ) и возрастанием степени
гомеостаза (независимости внутренней
среды организма от внешних условий),
можно говорить об ароморфозах. В
эволюции кровеносной системы хордовых
несомненными ароморфозами были развитие
сердца у древнейших позвоночных,
интенсификация его работы и формирование
замкнутости кровеносной системы у
челюстноротых, преобразования сердца
и магистральных сосудов при разделении
потоков артериальной и венозной крови
у пресмыкающихся, птиц и млекопитающих.
Это упростило задачу разделения крови
в сердце млекопитающих, которое стало
четырехкамерным, как у птиц. Желудочек
сердца разделился вертикальной
перегородкой на левый (от которого
начинается общий ствол дуг аорты) и
правый (откуда берет начало легочный
ствол). Впереди общий ствол дуг аорты
разделяется на левую дугу аорты
(продолжающуюся в спинную аорту) и
безымянную артерию, которая гомологична
редуцированной правой дуге аорты — она
не достигает спинной аорты и переходит
в правую подключичную артерию. Сонные
артерии начинаются симметрично: левая
— от левой дуги аорты, правая — от
безымянной артерии (у некоторых видов
млекопитающих обе сонные артерии
начинаются от общего ствола с правой
подключичной артерией, который называется
в этом случае плечеголовным). В венозной
системе млекопитающих, как и у птиц,
редуцировалась воротная система почек.
Возможно, в обоих случаях это связано
с повышением кровяного давления (по
сравнению с низшими позвоночными). При
видимом сходстве кровеносных систем
млекопитающих и птиц (четырехкамерное
сердце, единственная дуга аорты образует
спинную аорту, редукция воротной системы
почек) между ними имеются глубокие
различия. У птиц от левого желудочка
сердца начинается правая дуга аорты, а
левая утрачена, тогда как у млекопитающих
от левого желудочка начинается общий
ствол дуг аорты, левая дуга развита
полностью, а правая редуцирована. Эти
различия связаны с длительной независимой
эволюцией предков птиц и млекопитающих,
в ходе которой сходные прогрессивные
черты кровеносной системы были приобретены
разными путями. Вероятно, у общих предков
птиц и млекопитающих еще сохранялся
единый желудочек сердца и артериальный
конус, а также короткий общий артериальный
ствол — брюшная аорта. Разделение
желудочка сердца на камеры и брюшной
аорты на магистральные артерии происходило
самостоятельно и по-разному в
филогенетических стволах предковых
форм рептилий, ведущих к птицам и
млекопитающим. Развитие горизонтальной
внутрижелудочковой перегородки в сердце
типичных пресмыкающихся интересно как
пример инадаптивного пути эволюционных
преобразований. По В. О. Ковалевскому,
при инадаптивном развитии формирующееся
приспособление несет в себе предпосылки
для возникновения каких-либо внутренних
конструктивных дефектов, обнаруживающихся
не сразу, но постепенно нарастающих по
мере развития. Инадаптивная эволюция
является результатом своего рода
«неудачного» выбора направления
эволюционных изменений. А такой «неудачный
выбор» вполне вероятен: естественный
отбор благоприятствует любым изменениям,
которые дают какие-либо преимущества
организмам в борьбе за существование
в данный исторический момент, но не
может «предвидеть» ни результатов
дальнейшей эволюции в том или другом
направлении, ни новых форм взаимодействия
организма с внешней средой, которые
сложатся в будущем. На уровне организации
пресмыкающихся их неполная внутрижелудочковая
перегородка в целом отвечает физиологическим
потребностям этих животных, разделяя
артериальную и венозную кровь и в то же
время позволяя перераспределять потоки
крови в соответствии с характером
активности. Однако из-за своего положения
относительно устий предсердий и мест
отхождения артерий эта перегородка не
может полностью завершить разделение
желудочка сердца на артериальную и
венозную камеры. На базе такого устройства
сердца его дальнейшая прогрессивная
эволюция затруднена и требует довольно
сложных перестроек (произошедших у
крокодилов и птиц). В то же время на
первых этапах возникновения
внутрижелудочковой перегородки ее
вертикальное или горизонтальное
положение, так же как и разделение
брюшной аорты на три или на два
магистральных ствола, вероятно, не имели
существенных преимуществ друг перед
другом. Однако путь эволюционных
преобразований сердца у предков
млекопитающих был более перспективным
для полного разделения желудочка, что
обнаружилось на более поздних этапах
эволюции. В эволюции позвоночных, как
отмечалось выше, в целом происходило
общее усложнение и совершенствование
кровеносной системы. Однако не все
эволюционные перестройки кровеносной
системы представляют собой ароморфозы
(т. е. изменения на пути морфо-физиологического
прогресса, по А. Н. Северцову). Усложнение
организации само по себе еще не является
прогрессивным изменением. Иногда оно
служит своего рода компенсацией за
несовершенство какой-либо функции или
представляет собой результат развития
частного приспособления (идиоадаптации,
по А. Н. Северцову) к специфическому
образу жизни. Именно так следует оценивать
появление второго (малого) круга
кровообращения и усложнение строения
сердца у двоякодышащих рыб. Лишь в тех
случаях, когда усложнение строения
связано с интенсификацией функций, с
повышением уровня жизнедеятельности
(выражающимся в интенсификации процессов
обмена веществ) и возрастанием степени
гомеостаза (независимости внутренней
среды организма от внешних условий),
можно говорить об ароморфозах. В
эволюции кровеносной системы хордовых
несомненными ароморфозами были развитие
сердца у древнейших позвоночных,
интенсификация его работы и формирование
замкнутости кровеносной системы у
челюстноротых, преобразования сердца
и магистральных сосудов при разделении
потоков артериальной и венозной крови
у пресмыкающихся, птиц и млекопитающих.
.В филогенезе у одноклеточных ,и простейших многоклеточных организмов питательные вещества и кислород, необходимые для их роста и развития, воспринимались, непосредственно из внешней среды, в нее же они выделяли и продукты своей жизнедеятельности.
У многоклеточных, как более сложно устроенных организмов, по мере специализации и дифференциации их клеток непосредственное восприятие из внешней среды всего необходимого для обмена веществ становилось все более невозможным, поэтому у них развивались специальные органы питания и выделения. Соответственно повышалась способность ткани основного вещества — мезенхимы передавать питательные вещества и кислород от места, их усвоения к месту потребления сначала путем диффузии, а потом путем образования кровеносной системы.
Примитивная кровеносная система представлена жидкими участками (золь), заполняющими лакуны и каналы в студне (гель) основного вещества. Эти жидкие участки можно рассматривать как примитивную кровь, а лакуны и. каналы — как примитивные сосуды, но не имеющие еще собственных стенок. Последние обособляются от массы основного вещества позднее в виде эластической оболочки – интимы.
Первичная функция сосудистой системы — трофически-транспортная, что определяет ее связь с питающей трубкой, вокруг которой в первую очередь и развиваются сосуды. Перемещение крови в примитивной кровеносной системе происходит пассивно — только при движении тела и при перистальтике питающей трубки, т. е. при сокращении их мускулатуры.
С увеличением подвижности животных обеспечение более интенсивного обмена веществ пассивным перемещением крови в сосудах становится недостаточным. Поэтому у беспозвоночных, начиная с немертин, вокруг интимы некоторых сосудов появляются мышечные элементы, т. е. формируются сосуды с мышечной стенкой. Сокращение сосудистых стенок способствует самостоятельному, току крови, и сосуды из простого проводящего русла становятся двигательным аппаратом крови. Эта функция сосудистой системы усиливается с образованием замкнутой системы сосудов (у кольчецов) и особенно с появлением сердца (у моллюсков и членистоногих) в результате разрастания мышц в определенных участках сосудов. Максимального развития она достигает у наземных теплокровных животных, имеющих четырехкамерное сердце, которое становится центральным органом движения крови.
Самостоятельное передвижение крови, особенно при наличии сердца и замкнутой системы сосудов, обусловило возникновение «давления крови», а отсюда и формирование основных сосудистых стволов. Усиленный газообмен повлек за собой появление в крови дыхательных пигментов (гемоглобина) и возникновение специальных сосудов в области органов дыхания (в жабрах, легких). Повышенная выделительная функция крови стимулировала развитие специальных сосудов в области соответствующих выделительных органов (мезонефрос и метанефрос). Большая подвижность животных сопровождается разрастанием сосудистых сетей ив органах движения. Все это указывает на теснейшую связь сосудистой системы с работой самых различных органов.
Одновременно с усложнением строения сосудистой системы совершался процесс ее обособления от тканей основного вещества (мезенхимы, соединительной ткани) путем эндотелизации стенок сосудов, т. е.. нарастания эндотелия на внутренней поверхности интимы сосудов; Эндотелизация сосудов начинается уже у некоторых беспозвоночных, но только у водных позвоночных (рыб) она достигает высокой степени и закрепляется в онтогенезе образованием в мезенхиме ангиобластов (кровяных островков): Из клеток последних происходят не только эндотелий сосудов, но и клетки самой крови — эритроциты, лейкоциты, а у птиц и млекопитающих еще и тромбоциты. В связи с насыщением крови клетками закладываются и специальные органы кровообразования, в которых развиваются клетки крови.
Обособление сосудистой системы, особенно с образованием сердца, усиливается ее связями со специальной сосудисто-нервной системой (симпатическая) по мере развития в сосудах клеток, способных сокращаться, т. е. мышечных и эндотелиальных. В стенках сосудов появляется также большое число и интерорецепторов.
У членистоногих и моллюсков кровеносная система незамкнутая, но у последних благодаря наличию на жаберных сосудах примитивных сердец (по количеству жаберных щелей) выделяются венозные сосуды. Подобное примитивное строение кровеносной системы характерно для немертин, у которых имеются один спинной и два брюшных сосуда. В дорсальном сосуде кровь течет по направлению к голове, а в вентральных — к хвосту.
У аннелид замкнутая система кровеносных сосудов представлена дорсальным и вентральным продольными сосудами, метамерно соединенными между собой поперечными париетальными и висцеральными анастомозами. Кровь в этих сосудах перемещается в результате сокращения стенки спинного сосуда, чему иногда способствуют и сокращения, поперечных сосудов.
У примитивных хордовых (оболочники), как и у аннелид, имеются продольные спинной и брюшной сосуды, соединенные между собой поперечными висцеральными и париетальными анастомозами. Кровь перемещается благодаря сокращениям стенки дорсального сосуда. У ланцетника наряду с дорсальным артериальным продольным сосудом выделяются два краниальных венозных сосуда (яремные вены) и кардинальные вены, являющиеся продолжением хвостовых вен. Кардинальная и яремная вены, объединяясь, образуют венозный синус, продолжающийся в вентральную аорту, от которой отходят жаберные артерии, имеющие у своего начала расширения (жаберные сердца), обеспечивающие продвижение крови через капиллярные сосуды. У рыб венозный синус продолжается в предсердие, а затем в желудочек. Начиная с ланцетника, происходит образование воротной системы печени, которая берет начало под кишечной веной и заканчивается в печеночном выступе. Последний у рыб преобразуется в печень.
Хвостовые вены, проходя через почки, участвуют в образовании воротной системы почек, от которой, затем начинаются задние кардинальные вены, на уровне сердца сливающиеся с передними кардинальными венами.
У наземных позвоночных в связи с заменой жаберного типа дыхания на легочный отмечают редукцию и преобразование жаберных дуг, завершающееся развитием малого круга кровообращения. Последнее приводит к разделению предсердия на две камеры — правую (венозную) и левую (артериальную). У рептилий идет дальнейшее усложнение строения сердца, сопровождающееся частичным подразделением и желудочка на правую и левую половины (неполное), которое завершается у птиц и млекопитающих полным обособлением правой половины сердца от левой.
Из кровеносных сосудов наиболее сложным преобразованиям подвергаются жаберные артерий: из них развиваются дуга аорты и сонные артерии. Из шести жаберных дуг, свойственных высшим рыбам, у наземных позвоночных в образовании артериальных сосудов принимают участие только III, IV и VI пары, в то время как другие редуцируются или сохраняются лишь частично, соединившись с другими дугами; VI пара участвует в образовании легочных артерий, из которых левая сохраняет связь с дорсальной аортой в виде ductus arteriosus (у плода) или lig. arteriosum (у взрослых особей) между легочной артерией и аортой. Между правой артериальной дугой и дорсальной аортой связи нет; IV- пара жаберных артериальных дуг является источником образования правой и левой дуг аорты у рептилий, правой — у птиц, левой - у млекопитающих. У последних правая становится подключичной правой артерией; III пара жаберных артериальных дуг преобразуется во внутренние сонные артерии.
В зависимости от длины шеи, степени ее подвижности и от взаимоотношений с окружающими органами при формировании дуги аорты у различных видов домашних млекопитающих наблюдаются характерные, типы отхождения от нее сосудов. Исходной формой следует признать первоначальное отхождение от нее правой подключичной артерии, затем двух общих сонных и, наконец, левой подключичной артерии (у человека). При концентрации сосудов формируется плечеголовная артерия (у собаки), при слиянии обеих сонных артерий образуется общий ствол сонных артерий (у свиньи), и, наконец, плечеголовная артерия, сливаясь с левой подключичной артерией, образует общий плечеголовной ствол.
В онтогенезе сосудистая система начинает свое развитие в экстраэмбриональной области, в мезодермальном слое трофрбласта, а затем - в мезодерме желточного мешка в виде кровяных островков. В этих островках образующиеся небольшие щели постепенно сливаются в будущие кровеносные сосуды, выстланные одним слоем плоских эндотелиальных - клеток. Часть клеток, располагающихся в просвете первичных сосудов, постепенно округляется и преобразуется в ангиобласты. Внезародышевые сосуды участвуют в образовании сосудов желточного мешка и сосудов пуповины, которые соединяются с сосудами, развивающимися в мезенхиме тела зародыша. Развитие сосудов тела зародыша совпадает с закладкой первых сомитов. Эти сосуды первоначально представлены двумя .сердечными трубками, двумя дорсальными и двумя вентральными аортами. От дорсальных аорт отходят две желточные артерии (рис. 191), направляющиеся в желточный мешок, и два сосуда — к плаценте. От желточного мешка и от развивающейся плаценты к телу зародыша отходят парные желточные (8) и парные пупочные .вены (12), вступающие в каудаль'ные концы сердечных трубок, участвуя в образовании желточного круга кровообращения, который вскоре редуцируется, и плацентарного круга кровообращения, функционирующего До конца- плодного развития.
В конце 4-й недели эмбриогенеза (у крупных животных) дорсальные аорты объединяются в единую нисходящую аорту, развиваются парные краниальные и каудальные кардинальные вены, которые на каждой стороне тела сливаются в правую и левую кардинальные вены, впадающие в венозный синус каудального конца развивающегося сердца.
В краниальном отделе туловища дорсальные и вентральные аорты сохраняют свою самостоятельность и на каждой стороне тела соединяются между собой шестью жаберными дугами, из которых последняя развита не полностью.
Сердце закладывается в виде двух эндотелиальных трубок (рис. 192—5). После образования- головной кишки оба зачатка сердца срастаются в непарную сердечную трубку (10), вокруг которой" постепенно развивается миокард, а висцеральный листок перикарда образует эпикард (7).
На сердечной трубке различают венозный синус (1), предсердие (2), желудочек (3) и артериальный конус (4); из них, как и в филогенезе, сначала формируется трехкамерное сердце, а затем и четырехкамерное. Отличие заключается лишь в том, что. в плодный период в перегородке предсердий сохраняется овальное отверстие — for. ovale, временно служащее для плацентарного кровообращения, которое после рождения со стороны левого предсердия закрывается клапаном — valvula foraminis ovalis. Со стороны правого предсердия от бывшего здесь отверстия сохраняется лишь небольшой след в виде овальной ямки — fossa ovalis.
В желудочках сердца плодов разрастается густая сеть мышечных перекладин; у взрослых животных они превращаются в сухожильные струны, сосцевидные мышцы, мышечные перекладины и поперечные мышцы сердца.
Дифференцировка, которая привела к образованию Metazoa, потребовала коренного изменения способов питания, переноса кислорода и соответствующих анатомических особенностей. У животных всех классов наблюдаются и качественные, и количественные изменения общего энергетического метаболизма в зависимости от их видовой принадлежности. Например, у млекопитающих производство энергии на единицу веса обратно пропорционально размерам тела [998]. Более того, для каждой ткани в животном организме характерен свой собственный особый энергетический метаболизм. Так, у крысы интенсивность дыхания на 1 мог веса (величина Q02) для разных тканей различается в 10 и более раз [17]. Кроме того, эти ткани сильно различаются [17] по своей способности переключаться с брожения на дыхание, т. е. для них характерны различные точки Пастера (22, Е).
В конечном счете у всех известных животных поступление кислорода в клетки происходит путем обычной пассивной диффузии без зависящего от энергии (активного) транспорта. Но только очень мелкие животные, диаметром порядка миллиметра, могут удовлетворять свои потребности в кислороде, не имея для этого специальных органов. Многие современные «диффузионные» животные [1509] имеют форму листка толщиной менее 1миллиметра, например плоские черви (планарии), и если принимать во внимание только мета-болизирующие ткани, то к таким животным можно отнести и кишечнополостных (медузы и актинии). В условиях, когда 224 Глава 22 свободного кислорода в атмосфере было меньше, размеры таких животных также должны были быть меньше. Любое дальнейшее увеличение размеров должно было потребовать развития дыхательных органов и кровеносной системы. Первыми животными, которые приобрели их, были иглокожие (морские звезды, морские ежи)1. Но у этих животных в крови не содержалось пигментов. Хотя пигменты, родственные гемоглобину, наблюдаются уже у простейших (20, А), пигменты крови обнаруживаются впервые у аянелид (кольчатые черви, представителем которых, например, является дождевой червь). Высшие животные, как правило, имеют пигменты, причем иногда в довольно большом количестве; эти пигменты могут служить не только для переноса кислорода, но и для накапливания его. Так, у крысы гемоглобина содержится в 250 раз больше, чем ци-тохрома [499]. Но гемоглобины распределены в животном царстве беспорядочно, и, возможно, они имеют полифилетическое происхождение [936], хотя их предшественники должны быть родственными друг другу. Существуют даже рыбы, не имеющие пигментов крови, —¦ это антарктические ледяные рыбы [1588, 1789]. Несомненно, пигменты крови были утрачены ими в процессе эволюции. У моллюсков и рыб кислород поглощается через жабры. Большие количества воды, проходящие через жабры, уносят также большое количество тепла, в связи с чем эти животные пойкилотермны (холоднокровны). Хотя некоторые из более высокоорганизованных рыб (например, тунцы) могут, используя тонкий механизм противоточного теплообмена, поддерживать сравнительно высокую температуру в мышцах [345, 346], назвать их настоящими теплокровными животными нельзя. Переход на сушу потребовал, разумеется, крупных изменений механизмов, с помощью которых происходит поглощение кислорода. Существующие до выхода на сушу органы дыхания могли измениться. Некоторые из развившихся при этом механизмов доказали свою ценность у ограниченных групп животных, некоторые из них сохранились, но завели эти группы в эволюционный тупик. Только введение нового органа — легкого — предоставило возможность для дальнейшей эволюции. Об этом захватывающе рассказывается в книге Стина [1789]. Использование принципа, на котором основано устройство легкого, сделало возможным появление настоящих теплокровных животных, а затем и животных с высоко развитым мозгом. Как писал Стин, следствием этого явилось то, что теперь человек может писать о рыбах, а не наоборот. В норме поток электронов в дыхательной цепи (митохондрий служит для образования АТФ, и потому разобщение этих двух процессов рассматривается как вредное явление. Но некоторые виды тканей млекопитающих, а именно бурая жировая ткань, приспособились для функционирования в качестве отопителей (так называемая термогенная ткань). Такие ткани имеются, например, у новорожденных млекопитающих, а также у взрослых грызунов. Производство тепла на единицу веса за счет бурой жировой ткани может в 20 и более раз превышать производство тепла в обычной жировой ткани. Как показали эксперименты с изолированными митохондриями жировой ткани, в обычных физиологических условиях теплопродукция регулируется посредством «слабосопряженного состояния» и на нее не влияют биохимические стимулы [296, 376, 590, 591, 850, 928, 1149, 1483, 1724]. Заметим, что разобщение — это довольно специализированный механизм для производства тепла без превращения его в работу у теплокровных животных. Другим дополнительным источником для получения тепла служит, разумеется, мышечная дрожь [831]. Еще один очень общий и фундаментальный механизм производства тепла может состоять в стимулировании тиреоидными гормонами насоса, активно выводящего Na, что приводит к добавочному потреблению АТФ. Соответствующие эксперименты проведены на печени и мышцах крысы [913]. Увеличение работы насоса приводит к усиленной обратной диффузии Na, однако никакой видимой работы при этом не производится. Предполагают, что этот процесс произошел в филогенезе от направленной стимуляции мышц у низших позвоночных, а именно стимуляции для движения животного к месту с оптимальной внешней температурой [1798]. Тогда можно сказать, что насос отключили от органа, который он обслуживал, и разорвалась связь между мышечной активностью и терморегуляцией, осуществляемой с помощью поведенческих реакций. Сравнительная физиология теплопродукции в различных группах позвоночных изложена в работах Уиттоу [1987].
У членистоногих, этой необычайно успешной ветви высших беспозвоночных, включающей насекомых, развились резко отличающиеся от присущих позвоночным механизмы, обеспечивающие достаточное поступление кислорода [1994]. Система трахей, характерная для насекомых, развивалась много раз независимо. В этом случае нет локализованного органа дыхания. Воздух поступает .в организм животного по системе, состоящей из множества тонких трубочек — трахей, откуда кислород диффундирует непосредственно в клетки. Этот тип дыхательной системы не позволяет насекомым достигать очень больших размеров, за что мы должны быть очень благодарны Природе [967]. Самыми крупными из когда-либо существовавших насекомых были стрекозы каменноугольного периода. Так как у высших организмов сохранились механизмы брожения, их клетки в условиях недостатка кислорода хотя бы частично могут возвращаться к брожению. Это позволяет многим растениям и животным, ааробным в обычных условиях, удивительно хорошо выдерживать недостаток кислорода [1698]. Разумеется, растения, которые сами способны вырабатывать кислород, должны гораздо меньше страдать от его недостатка при условии, однако, хорошего освещения. Поразительное следствие способности к брожению у позвоночных состоит в том, что они могут входить в так называемую «кислородную задолженность», чтобы производить максимум энергии в тех случаях, когда требуется большое мышечное усилие. У животных, способных к быстрому передвижению, образуются большие количества молочной кислоты, но при этом им не требуется тут же поглощать соответствующее количество кислорода. Молочная кислота удаляется позднее, снова превращаясь в углевод за счет энергии, получаемой при дыхании. Брожение прекращается, как только снова в достаточном количестве появляется кислород. При более тщательном исследовании оказалось, что брожение ингибируется кислородом. Это явление уже давно было известно биохимикам как эффект Пастера (13, Б, 22, Е; [17, 319, 561, 1044, 1502, 1503]), и его часто изучали на простых эукариотах, а именно на дрожжах. Среди прокариотов оно наблюдается уже у факультативных аэробов, например, у Escherichia coli. Многие дифференцированные беспозвоночные, которые обычно живут при небольшом давлении кислорода (обитатели донного ила и эндопаразиты, включая авнелвд, гельминтов и моллюсков) приспособились к частичному анаэробиозу; для разных организмов характерны различные предпочитаемые ими давления кислорода [269, 270, 617]. Таким образом, брожение у них .вновь, как у некоторых простейших (20, А), прочно выходит на первое место [269, 270, 314, 578, 853, 1621]. Однако не раз было показано, что даже у «анаэробных» паразитов сохраняются митохондрии (пусть измененные) и остается способность к дыханию. Это относится к круглым [1751] и ленточным червям [372], <а также к паразитическим стадиям трематод, обитающих в крови [411]. Пока не ясно, может ли кто-нибудь из паразитов долго и на всех стадиях развития жить при полном отсутствии кислорода [269, 270]. Во всяком случае, не все эндопаразиты являются частичными или полными анаэробами. Не только для простейших (20, А) кровь служит аэробным местообитанием. Некоторые паразитические черви прекрасно извлекают кислород из больших объемов крови. Некоторые эндопаразиты «обленились» и выделяют продукты брожения даже в тех случаях, когда они располагают достаточными количествами кислорода. Видимо, у них отсутствует эффект Пастера. Различные труппы позвоночных развили множество способов справляться с продолжительным недостатком кислорода в среде, накапливать кислород и сокращать его потребление. Одними из первых исследователей в этой области были Ирвинг [910] и Шоландер [1650]. Для человека особенно интересны позвоночные, способные нырять и подолгу оставаться под водой, например морские черепахи, тюлени и киты [55, 56, 936, 1126, 1789]. Большинство из этих животных должно время от времени вдыхать воздух. Самые выдающиеся мастера подводного плавания—-это черепахи; некоторые из них могут оставаться под водой часами и даже долее [186, 187]. Черепаха Pseudemys scripta, способная неделями оставаться под водой, заслуживает звания позвоночного —¦ факультативного анаэроба [1526, 1808]. Эти черепахи резко уменьшают свой аэробный энергетический метаболизм и получают энергию в основном за счет брожения, хотя в данном случае нельзя пренебрегать и возможностью поглощения некоторого количества кислорода и через кожу. При длительном пребывании под водой, по-видимому, даже мозг и сердце, которые у других ныряющих животных особенно нуждаются в кислороде, переходят на брожение. Как и у других ныряющих животных, общая интенсивность образования энергии в воде у черепах ниже, чем на воздухе. Добавление в верстке. Прекрасный обзор метаболических последствий ныряния у животных и человека недавно опубликовали Хочачка и др. [854].
Термин "дыхание" используют для обозначения тех процессов, при помощи которых животные и растительные клетки потребляют кислород, отдают углекислоту и переводят энергию в форму, доступную для биологического использования, например, в форму химической энергии, заключенной в фосфатных связях АТФ. В биологии понятие дыхания имеет три разных значения. Первоначально оно означало внешнее дыхание, т. е. вдыхание и выдыхание воздуха. Позже, когда стало ясно, что существенным процессом является обмен газами между клеткой и окружающей ее средой, этим термином стали обозначать газообмен. Наконец, когда стали известны детали клеточного метаболизма, это понятие отнесли к тем ферментативным реакциям в клетке, которые ответственны за использование кислорода (цитохромы называют дыхательными ферментами).
Взаимосвязь типов дыхательных систем и среды обитания
Выделяют два основных типа приспособлений организмов к среде обитания. Первый тип связан с выживанием и последующим нормальным функционированием организма при изменении конкретных условий существования. Данный тип адаптации осуществляется при помощи биохимических, физиологических и поведенческих механизмов. Второй тип приспособлений носит эволюционный характер. Речь идет о закрепленных в ходе эволюции тех или иных особенностях строения тела или физиологических реакции, обеспечивающих оптимальное взаимодействие со средой, в том числе органов дыхания. Способ, при помощи которого кислород извлекается из окружающей среды (внешние дыхание) и доставляется в клетки для участия в выработке энергии, в значительной степени определяет общий уровень биоэнергетики особи. В процессе эволюции выработалось несколько типов дыхания, и эффективность их функционирования крайне важна для нормального энергообеспечения организма. Уровень энергетического обмена определяется в первую очередь типом дыхания (жаберным, легочным или трахейным) и является базовой величиной, на фоне которой происходят частые изменения биоэнергетики под влиянием различных экологических факторов (температуры, солености, освещенности, химического состава воздуха или воды и т.д.). Следовательно, среда обитания (водная или воздушная) имеет определяющее значение для формирования того или иного типа дыхания (или часто встречающаяся их комбинация). Каждый из способов дыхания в результате отбора в ходе эволюции превратился в наиболее эффективный тип извлечения кислорода из среды обитания того или иного организма.
У наиболее просто организованных животных (простейших, губки, кишечнополостные) специализированные органы дыхания отсутствуют, и кислород попадает в клетки диффузно из воды через клеточные оболочки. Диффузия происходит потому, что концентрация кислорода в воде выше, чем в клетках данных организмов. Однако такой способ дыхания может обеспечить кислородом только мелкие организмы. Ведь кислород способен диффундировать внутрь клетки только на 0,5 мм. Уровень энергетического обмена животных, не имеющих специальных органов дыхания и обладающих небольшой подвижностью, очень низкий. У более высокоорганизованных животных появляются специализированные участки тела, воспринимающие кислород из окружающей среды, - жабры, легкие и трахеи. Однако между неспецифическим способом дыхания, когда кислород поступает в организм путем простой диффузии, и специализированными органами дыхания имеется промежуточный тип газообмена - кожное дыхание. В этом случае кислород диффундирует через всегда увлажненную кожу, обильно снабженную кровеносными сосудами, и с кровью доставляется к различным тканям. У многих животных, например у плоских и круглых червей, а также некоторых других беспозвоночных, кожное дыхание - единственный способ получения кислорода из окружающей среды. Кожное дыхание в качестве дополнительного способа газообмена встречается и у позвоночных, например у амфибий. Общая особенность специализированных органов дыхания – очень большая площадь их поверхности, контактирующая с водой или воздухом, что значительно повышает эффективность газообмена. Для органов дыхания характерно обильное кровоснабжение, что необходимо для эффективной доставки диффундирующего кислорода к отдельным тканям. Исключение в этом отношении составляют только насекомые, у которых кислород по трахеям (воздухоносным трубочкам) доставляется непосредственно к отдельным тканям и клеткам. Наконец, непременное свойство органов дыхания – легкость диффузии кислорода через мембраны клеток, которые составляют пограничный с окружающей средой слой жабр или легких. В зависимости от конкретных условий обитания и образа жизни той или иной группы животных имеется большое разнообразие возможных приспособлений для усиления эффективности дыхания. В жабрах эти приспособления направлены на увеличение площади поверхности, контактирующей с водой, а также усиление потока омывающей эти органы воды, что приводит к более эффективной диффузии кислорода внутрь клетки.
Наибольшее разнообразие органов дыхания и приспособлений, увеличивающих эффективность газообмена, наблюдается у водных беспозвоночных животных. Связано это с очень большим разнообразием условий обитания данных организмов. Впервые специализированные органы дыхания – жабры – появляются у водных беспозвоночных: многощетинковых червей – полихет. Роль жабр у них выполняют видоизмененные участки конечностей, имеющие перистую, кустистую или листовидную форму. Внутрь таких жабр заходят ветвящиеся кровеносные сосуды, по которым обогащенная кислородом кровь разносится по всему телу. Сходное строение жабр наблюдается у малощетинковых червей – олигохет и у пиявок. С появлением специализированных органов дыхания у червей уровень энергетического обмена у них становится выше, чем у простейших, губок и кишечнополостных, лишенных этих органов. Большая вариабельность строения органов дыхания среди беспозвоночных наблюдается у беспозвоночных: ракообразных, паукообразных и насекомых. В силу особенностей образа жизни этих животных их органы дыхания приспособлены для извлечения кислорода, как из воды, так и из воздуха. Так, у многих ракообразных органы дыхания представлены жабрами, которые, как и у червей, находятся в тесной связи с конечностями. Это способствует усилению вентиляции и увеличению эффективности газообмена. У десятиногих раков, к которым принадлежит речной рак, жабры находятся внутри особой жаберной полости по бокам тела. Боковые складки панциря этих животных защищают жабры от высыхания у наземных ракообразных. У них образуется система разветвленных дыхательных трубочек – зачатковых трахей насекомых. Степень развития жабр у ракообразных зависит от их образа жизни, в частности от двигательной активности. Так, у крабов, ведущих подвижный образ жизни, площадь поверхности жабр в расчете на единицу веса тела в 2 раза выше, чем у малоподвижных крабов. Дыхательная система ракообразных чутко реагирует на изменение экологических факторов. Например, у раков при снижении концентрации кислорода в водной среде увеличивается объем прокачиваемой через жабры воды и на 40% возрастает извлечение из нее кислорода. Помимо жабр, у многих членистоногих, ведущих наземный образ жизни (паукообразные, насекомые), органами дыхания служат легочные мешки и трахеи. В последнем случае кислород по трахеям (воздухоносным трубочкам) переносится непосредственно к отдельным тканям и клеткам без участия кровеносной системы. Трахеи сообщаются с внешней средой при помощи специальных отверстий – дыхалец. В тканях тела насекомого трахеи многократно ветвятся и заканчиваются мельчайшими трубочками – трахеолами, которые на концах окружены трахеальной клеткой с радиально расходящимися отростками, пронизанными канальцами. Трахеолы образуют сеть в тканях и отдельных клетках. Так, в мышцах цикады расстояние между соседними трахеолами равно 6,5 мкм. У некоторых насекомых имеются расширенные участки трахей (воздушные мешки), которые могут сжиматься и разжиматься при движении тела, что приводит к насасыванию воздуха в трахейную систему. Обычно трахеи у насекомых вентилируются при помощи мышечных сокращений. Частота дыхательных движений, а также закрывание – открывание дыхалец зависят от концентрации СО2 и О2 во вдыхаемом воздухе. Домашняя пчела совершает в покое 40 дыхательных движений в минуту, а в полете – 120. Частота дыхательных движений зависит от температуры. При увеличении температуры воздуха этот показатель у саранчи может возрастать более чем в 5 раз. Столь тщательное рассмотрение строения и функционирования трахей у насекомых не случайно, ибо им присущ очень высокий уровень энергетического обмена, самый высокий среди беспозвоночных. Одна из причин такой особенности – трахейный тип дыхательной системы с непосредственной доставкой кислорода к отдельным клеткам, минуя промежуточные звенья, на которых происходят потери эффективности газообмена. Общей причиной столь значительного уровня биоэнергетики насекомых можно считать их высокую двигательную активность. Ведь полет насекомого требует очень больших энерготрат. Постоянная нагрузка на мышечную систему при полете привела к установлению в процессе эволюции высокого уровня энергетического обмена. На электронно-микроскопических срезах через летательные мышцы насекомых видны необычайно развитые гигантские митохондрии, занимающие практически весь объем цитоплазмы между миофибриллами. У насекомых, как и у других животных с высокой двигательной активностью, в процессе эволюции закрепились структурные и функциональные особенности дыхательной системы, обеспечивающие высокий уровень энергетического обмена. Возможно, одной из причин того, что эти животные в прямом смысле завоевали всю Землю (насекомых в природе больше, чем других животных, - более 1 млн. видов), - высокая дыхательная активность и уровень биоэнергетики. Трахейная дыхательная система насекомых – исключение среди беспозвоночных, дышащих в основном при помощи жабр. Дыхание при помощи жабр характерно для большинства моллюсков, обитающих в воде. Как и у членистоногих, строение жабр у моллюсков варьирует в соответствии с их образом жизни. Жабры у моллюсков расположены или по бокам тела, или в мантийной полости и снабжены ресничками, при помощи которых регулируют ток воды. Двустворчатые моллюски периодически открывают и с силой закрывают створки раковины, прокачивая большие объемы воды и очищая полость мантии: мидии весом около 100 г прокачивают за 1 ч в среднем 205 л воды. При этом, однако, эффективность извлечения кислорода из воды не очень высокая и составляет 3-5 %.У некоторых брюхоногих моллюсков, живущих в море, эффективность извлечения кислорода из воды гораздо выше-40-80%. Следовательно, диапазон приспособляемости у этих животных очень большой. У моллюсков, ведущих наземный образ жизни (легочные моллюски), жабры редуцировались, и органом дыхания стали легкие. Они образовались в результате срастания мантийной полости и стенки тела. Определенную роль в газообмене легочных моллюсков играет кожное дыхание. Своеобразно устроены органы дыхания у иглокожих (морских звезд, морских ежей, голотурий, офиур). Эти животные обитают преимущественно на морском дне и ведут малоподвижный образ жизни, что сказывается на уровне их энергетического обмена. Основным органом дыхания служат примитивно устроенные кожные жабры – выпячивания стенки тела, через которые из воды диффундирует кислород. Циркулирующая внутри этих впячиваний целомическая жидкость переносит кислород к различным тканям и органам. У некоторых иглокожих (офиуры) специальные органы дыхания отсутствуют вообще, как и у самых простых организмов (губки, кишечнополостные). Следствие столь примитивного строения дыхательной системы у иглокожих – чрезвычайно низкий уровень энергетического обмена. А ведь иглокожие относятся к вторичноротым животным – более высокоорганизованным, чем первичноротые: черви, членистоногие, моллюски. Строение и функционирование органов дыхания беспозвоночных отличается большим разнообразием, что связано со спецификой их образа жизни. В соответствии с идеей академика А. Н. Северцова по мере усложнения организации животных энергетический обмен возрастает. Однако условия обитания тех или иных животных способны значительно модифицировать эту тенденцию. Тип дыхательной системы у позвоночных животных, как и у беспозвоночных, определяется средой обитания. Но у позвоночных строение органов дыхания не столь разнообразно. Их дыхательный аппарат связан с одной и той же областью тела (передним отделом кишечника). Это касается жабр, как водных позвоночных, так и легких наземных позвоночных. Жаберные щели как компоненты дыхательной системы закладываются во время зародышевого развития не только водных позвоночных, но и наземных, дышащих легкими. Однако в последнем случае закладки жабр присутствуют только в период эмбриогенеза, а на более поздних этапах развития они редуцируются. Основные органы дыхания у позвоночных – жабры и легкие. Определенную роль в газообмене этих животных играет кожа и некоторые внутренние органы (кишечник, плавательный пузырь,слизистая ротовой и околожаберной полостей). Рассмотрим жаберный тип дыхания на примере круглоротых и рыб. У водных позвоночных дыхание (поглощение кислорода из воды) осуществляется в жаберных щелях – парных отверстиях в стенке глотки. Жаберные щели соединяют глоточную полость с внешней средой. Кислород диффундирует из воды в жаберные лепестки, расположенные на жаберных дугах – краях жаберных щелей. Большая площадь поверхности жаберных лепестков постоянно омывается свежей водой. Они обильно снабжены кровеносными сосудами, и кислород, диффундирующий в кровь, разносится по всему телу. У наиболее примитивных современных позвоночных круглоротых (миноги и миксины) дыхательная система представлена 7 парами жаберных мешков. Они при помощи внутренних жаберных отверстий сообщаются, с одной стороны, с дыхательной трубкой, а с другой – через наружные жаберные отверстия с внешней средой. Жаберные мешки содержат многочисленные жаберные лепестки, в которых и происходит диффузия кислорода. Процесс дыхания осуществляется вследствие сжимания и разжимания (расслабления) жаберного аппарата. При этом вода входит и выходит через наружные жаберные отверстия. Жаберный аппарат у круглоротых занимает очень большой относительный объем по сравнению с другими позвоночными животными – примерно 1/6 часть общего объема тела. Строение и функционирование жабр круглоротых весьма примитивно по сравнению с рыбами. Рыбы дышат в основном при помощи жабр, хотя у некоторых видов в этом процессе участвуют также плавательный пузырь, кишечник, выстилка ротовой и околожаберных полостей. Эти способы газообмена у рыб играют вспомогательную роль по сравнению с жабрами. Некоторые рыбы, живущие в пересыхающих и обедненных кислородом водоемах, дышат при помощи жабр и легких. Жабры у рыб в отличие от круглоротых – хорошо дифференцированный, высокоспециализированный, эффективно функционирующий орган дыхания. Высокой эффективности процесса дыхания у рыб способствует и активное прокачивание воды через жабры благодаря движению жаберных крышек, снабженных мускулаторой. Когда жаберная крышка поднимается, – под ней образуется объем с пониженным давлением. В результате этого ода всасывается из ротовой полости в боковую жаберную полость. Когда жаберная крышка опускается, – вода из жаберной полости выталкивается через жаберные щели наружу. При этом свежая воды омывает жаберные лепестки, где происходит диффузия кислорода в кровь. Жаберные крышки играют роль насоса, прокачивающего воду через жабры, что способствует их активной вентиляции. У многих пелагических рыб – прекрасных пловцов, обитающих в открытом море, вода пассивно прокачивается через жабры. Во время быстрого плавания рот у рыб открыт и жаберные крышки неподвижны, что создает непрерывный поток свежей воды через жаберную систему. Этот тип вентиляции жабр, называемый таранной вентиляцией, наиболее ярко выражен у тунцов, для которых быстрое постоянное плавание – необходимое условие снабжения организма кислородом. Энергетические потребности тунцов в силу особенностей плавания (они совершают скоростные миграции в океане на сверхдальние расстояния) – самые высокие среди рыб. Необычайно высокая площадь поверхности жабр значительно увеличивает возможность диффузии кислорода из воды в организм. На жаберных дугах находятся многочисленные омываемые свежей водой жаберные пластинки, через поверхность которых и происходит диффузия кислорода. Жаберные пластинки устроены весьма сложно. На более крупных первичных пластинках располагаются мелкие многочисленные вторичные пластинки. Они покрыты множеством микрогребней и микроворсинок, что еще более увеличивает площадь дыхательной поверхности жабр. Количество и размеры жаберных пластинок значительно варьируют и определяются условиями обитания рыб: в основном содержанием кислорода в воде, а также типом и скоростью плавания. У рыб, обитающих в воде с низким содержанием кислорода, первичные жаберные пластинки длиннее, а число вторичных пластинок больше, чем у рыб, живущих в водоемах с высокой концентрацией кислорода. Для сравнения площади поверхности жабр различных видов рыб или других животных пользуются величиной, называемой относительной дыхательной поверхностью. В этом случае общую площадь поверхности жабр делят на массу животного. Оказалось, что наиболее высокая относительная площадь поверхности жабр у тунцов, преодолевающих в океане стайерские дистанции со спринтерской скоростью и в связи с этим расходующих для плавания громадное количество энергии. Большую относительную площадь поверхности жабр имеют скумбрии, меч-рыба, парусник – прекрасные пловцы. Для малоподвижных рыб, например скорпен, смарид, плотвы, характерны минимальные величины относительной площади поверхности жабр и как следствие уровня энергетического обмена. Эти показатели у скарпены и у тунца различаются примерно в 100 раз, что свидетельствует о решающей роли экологических факторов в установлении энергообеспечения того или иного вида рыб. Эффективность газообмена у рыб обеспечивает и жаберная кровеносная система. Ток крови в жаберных капиллярах направлен против тока воды, омывающей жабры, что значительно увеличивает эффективность извлечения кислорода из водной среды. Эффективность дыхания зависит и от скорости кровотока, степени развития кровеносных сосудов, концентрации эритроцитов в крови, кислотной емкости крови – способности поглощать то или иное количество кислорода. Хрящевые рыбы (акулы, скаты) извлекают из воды примерно 75% кислорода, а костистые рыбы – 80-85%, тогда как круглоротые, имеющие более совершенный жаберный аппарат, - только 10-25%. Существует тесная количественная взаимосвязь между уровнем интенсивности дыхания и относительной площадью поверхности жабр у рыб. На этом основании предполагается, что наблюдающееся снижение уровня энергетического обмена у рыб по мере увеличения их массы обусловлено уменьшением относительной площади дыхательной поверхности при возрастании веса этих животных. Наиболее поразительная особенность некоторых рыб – дыхание воздухом при помощи легких. Двоякодышащие рыбы и многоперы способны как к жаберному дыханию в воде, так и к легочному – при помощи воздуха. Их легкие представляют собой примитивно устроенные мешки с гладкими внутренними стенками. Легкие начинают функционировать в том случае, когда рыба оказывается вне воды и дыхание жабрами невозможно. Эти рыбы обитают в воде с низким содержанием кислорода и в регулярно пересыхающих водоемах Африки, Южной Америки, Австралии. Например, представитель чешуйтчатниковых рыб южноамериканский лепидосирен получает кислород почти полностью за счет дыхания легкими, а виды этих рыб, обитающие в Австралии, имея легкие, дышат в основном жабрами. Легочное дыхание у рыб, типичных обитателей водной среды, - наглядный пример сложившихся в процессе эволюции приспособлений газообмена к меняющимся факторам окружающей среды. Другие позвоночные дышат при помощи легких. Легочное дыхание значительно отличается от жаберного. Ведь концентрации кислорода в воздухе различаются. В воздухе 20,95% кислорода, а в воде менее 1%. В 1 л пресной воды содержится 6,57 мл кислорода, а в морской – 5,31 мл. Кроме того, в воздухе диффузия кислорода происходит в 3 млн. раз быстрее, чем в воде. Помимо различий, эти два основных типа дыхания имеют и общие черты. Внутренняя поверхность легкого всегда покрыта пленкой влаги, кислород сначала диффундирует через нее и только после этого переносится в кровь через легочный эпителиальный слой. Так что и при легочном и при жаберном дыхании кислород поступает в кровь из водной фазы. Смена в процессе эволюции водной среды обитания на воздушную, связанная с выходом водных животных на сушу, стала важнейшим эволюционным этапом в развитии организмов. В связи с приспособлением к наземному образу жизни многие черты строения животных значительно изменились. Изменения коснулись и дыхательной системы – вместо жабр появились легкие. Строение легких и эффективность их работы естественно различаются в зависимости от систематического положения того или иного организма, а также в огромной степени от экологических условий, в которых они обитают. В ряду наземных позвоночных: амфибии – рептилии – птицы – млекопитающие сложность строения легких возрастает, но в каждой из этих групп строение данных органов значительно меняется в зависимости от факторов среды. У амфибий легкие закладываются во время зародышевого развития в виде парных выпячиваний позади последней пары жаберных мешков – образований, из которых у рыб возникают жабры. На этом основании предполагается, что легкие у наземных позвоночнных животных сформировались в результате преобразования последней пары жаберных мешков, характерных для водных позвоночных. Вопрос о происхождении легких всегда занимали эволюционистов, и на этот счет существует несколько точек зрения. Кроме изложенной выше, существует идея, согласно которой легкие появились в результате специализации парного плавательного пузыря у рыб. У амфибий легкие устроены примитивно – небольшие мешковидные образования с относительно гладкими внутренними стенками. Простота строения легких в данном случае объяснима. Ведь этот орган появляется у амфибий в ходе эволюции фактически впервые. (У двоякодышащих рыб крайне примитивные легкие лишь дополняют другие органы дыхания.) Кроме того, у амфибий большой вклад в газообмен вносят кожа и слизистая оболочка ротовой полости. У некоторых примитивных амфибий (протеус) легкие представлены двумя асимметричными мешками с гладкой внутренней поверхностью. У других амфибий (тритоны, саламандры) внутренняя поверхность легких увеличивается благодаря образованию перегородок и ячеек. При всем при том относительная площадь дыхательной поверхности легких невелика и заметно отличается у разных видов амфибий. Наименьшие величины характерны для саламандр, а наибольшие – для лягушек. Легкие у амфибий активно вентилируются при помощи дыхательных движений, в которых мускулатура ротовой полости и глотки, а также ноздри, способные открываться и закрываться. Воздух через ноздри попадает в ротовую полость, дно которой при этом опущено. Затем ротовая полость сокращается, – и воздух выталкивается в легкие. После этого сокращаются брюшные мышцы, – и воздух попадает обратно в ротовую полость. Важную роль в газообмене у амфибий играет постоянно увлажненная кожа, снабженная многочисленными кровеносными сосудами, что способствует диффузии кислорода из окружающей среды. Соотношение площади поверхности легких и кожи у амфибий в среднем равно 2:3, однако вклад легочного и кожного дыхания у разных видов этих животных значительно отличается. Например, у прудовой лягушки через легкие и кожу поступает примерно одинаковое количество кислорода. А у травяной лягушки легочное дыхание составляет 70%. У хвостатых амфибий (саламандры, тритоны) кожное дыхание играет доминирующую роль в газообмене. Так, у американской безлегочной саламандры (животное без легких – удивительный пример адаптации!) и дальневосточного тритона весь газообмен осуществляется через кожу. Интересно, что у амфибий четко выражены сезонные колебания газообмена и связаны они с легочным дыханием, а кожное дыхание на протяжении года практически не изменяется. У рептилий легкие устроены сложнее, чем у амфибий. Ведь эти животные почти полностью перешли к наземному образу жизни, а кожное дыхание из-за развития рогового слоя кожи у них отсутствует. Только у черепах, связанных с водной средой, помимо легких, в газообмене участвует слизистая оболочка глотки и частично кожа. Легкие у рептилий представлены в виде парных или непарных мешков с пористой, или ячеистой, внутренней структурой. Внутренняя структурированность легких менее выражена, например, у гаттерии и более – у черепах и особенно у крокодилов. Относительная площадь поверхности легких у рептилий невелика: минимальная величина отмечена у варана, а максимальная (в 5 раз выше) – у миссисипского крокодила и питона. У рептилий дыхательные пути устроены сложнее, чем у амфибий. Воздух из ротовой полости через гортанную щель поступает в гортанную полость. От гортани отходит длинная трахея, разделяющаяся в конце на два бронха, через которые воздух попадает в легкие. Дыхательный акт осуществляется при растяжении и сужении грудной клетки и с участием ребер. Сравнивая круглоротых, рыб, амфибий и рептилий, относящихся к холоднокровным организмам, следует отметить одну особенность. Несмотря на различия в их организации, важные морфологические и физиологические преобразования, связанные с переходом к наземному существованию, а также несмотря на разные механизмы газообразования (жабры, легкие, дополнительные органы дыхания), уровень энергетического обмена у этих животных отличается мало. Строение и функционирование легких у теплокровных животных (птиц, млекопитающих) намного сложнее, чем у рассмотренных выше холоднокровных позвоночных. Происходит это в результате значительного усложнения внутренней структуры легких и как следствие увеличения относительной площади дыхательной поверхности, также благодаря более совершенным механизмам активной вентиляции легких. Все эти особенности, а также более высокая постоянная температура тела приводят к тому, что теплокровных животных уровень биоэнергетики намного выше (в среднем более чем в 10 раз) по сравнению с теплокровными животными. Наиболее высокий уровень энергетического обмена среди всех животных – как холоднокровных, так и теплокровных – у птиц. Эволюция легких у теплокровных позвоночных животных привела к увеличению количества и уменьшению размеров ячеек и камер, составляющих внутреннюю структуру этих органов. Легкие у птиц имеют губчатое строение и состоят из системы воздухоносных трубочек, открытых с обоих концов. В этих трубочках и при вдохе, и при выдохе воздух движется в одном направлении, и поэтому газообмен непрерывен. Очень высокая эффективность газообмена. У птиц обуславливает самый высокий среди животных уровень биоэнергетики. У птиц воздух через трахеи попадает в бронхи, которые в легких ветвятся на вторичные и третичные бронхи, а затем на бронхиолы. Последние разделяются на более тонкие воздухоносные капилляры. Они, как и бронхиолы, оплетены густой сетью кровеносных сосудов. В этих частях легких и происходит диффузия кислорода в кровь через тонкую пленку влаги, выстилающую внутренние стенки бронхиол и воздухоносных капилляров. Путь вдыхаемого воздуха заканчивается в воздухоносных мешках – выростах бронхов. 5 пар воздухоносных мешков, во много раз превышеющих по объему легкие, расположены между внутренними органами и активно участвуют в продувании воздуха через легкие, хотя процесс газообмена в них не происходит. Относительная площадь поверхности легких у птиц очень высокая и зависит от двигательной активности этих животных. Например, если у малоактивной курицы эта величина составляет всего 1,1 м2/кг веса тела, то у голубя – 17,5 , а у колибри – 66,6 м2/кг веса тела. Прослеживается прямая зависимость между уровнем подвижности птиц, относительной площадью дыхательной поверхности легких и как следствие уровнем биоэнергетики. Акт дыхания у птиц в покое происходит благодаря расширению и сужению грудной клетки при помощи реберных мышц. Во время полета в дыхательном акте участвуют воздухоносные мешки, которые при подъеме крыльев растягиваются и наполняются воздухом из легких, а при опускании – сжимаются, и воздух выталкивается в легкие. Диффузия кислорода в легких происходит как при вдохе, так и при выдохе, и это создает условия для высоко эффективного газообмена и рекордно высокого уровня биоэнергетики. Для легких млекопитающих характерно альвеолярное строение, обособление сложных дыхательных путей и дальнейшее усложнение механизма дыхания с участием диафрагмы. Воздух попадает в легкие через бронхи, которые затем многократно ветвятся, образуя бронхиолы. Последние заканчиваются альвеолами, имеющими ячеистое строение. Альвеолы опутаны густой сетью кровеносных сосудов, и здесь происходит диффузия вдыхаемого кислорода в кровь. Количество альвеол в легких млекопитающих огромное: от 6 млн. у малоподвижных ленивцев до 500 млн. у хищников. Общая дыхательная поверхность легких благодаря альвеолярному строению громадная. Так, у человека она составляет 10 м2, а у лошади – 500 м2. Относительная дыхательная поверхность у разных видов значительно отличается и связана в основном с уровнем их подвижности. Наибольшая относительная поверхность легких (примерно 5 м2/кг веса тела) у высоко подвижных летучих мышей, землероек, а наименьшая – у ежей, ленивцев. У человека этот показатель не очень высокий – он составляет примерно 1 м2/кг веса тела. Усиленной вентиляции легких млекопитающих способствует дыхательное движение грудной клетки и диафрагмы. Уровень биоэнергетики у млекопитающих ниже, чем у птиц, что связано в основном с большой двигательной активностью последних. Отметим, что сложность строения легких у птиц и млекопитающих – не единственная причина высокого уровня энергетического обмена у этих животных. Высокий уровень биоэнергетики у теплокровных организмов объясняется не только сложным строением и функционированием легких, но и постоянно высокой температурой тела. Температура тела птиц в среднем 410С (колебание температуры у разных видов от 38 до 43, у млекопитающих несколько ниже – в среднем 370 при колебании у разных видов от 35 до 39). Высокая и постоянная температура тела у теплокровных животных поддерживается при помощи химической терморегуляции (рефлекторное увеличение теплопродукции при понижении температуры окружающей среды) и физической терморегуляции (теплоизоляция при помощи перьев у птиц и меха у млекопитающих). Поддержание высокой температуры тела возможно только при очень высоком уровне биоэнергетики, чем и отличаются теплокровные животные от холоднокровных. Влияние экологических факторов на энергетический обмен. Существование любого организма в природе протекает при постоянно меняющихся условиях внешней среды. Меняются температура, влажность, освещенность, радиационный фон, химический состав воды и воздуха. На животных воздействуют различные вещества, в том числе и токсиканты. Существование животных в природных условиях неизбежно сопряжено со стрессом. Все эти факторы влияют на разные стороны жизнедеятельности организмов, на многие физиологические и биохимические процессы, в том числе и на уровень энергетического обмена, весьма чутко реагирующего на воздействие экологических факторов. Наиболее основательно изучено влияние на процессы жизнедеятельности, на метаболизм и, в частности, на энергетический обмен самого универсального фактора окружающей среды – температуры. Она меняется на протяжении сезона, суток и более коротких отрезков времени. Любая особь реагирует на кратковременные или продолжительные изменения температуры, однако, в процессе эволюции сформировались два независимых фундаментальных типа реакции организмов на изменение температуры (Иванов, 1965; Шилов, 1985). Речь идет о холоднокровных (пойкилотермных, или эктотермных) животных и теплокровных (гомойотермных, или эндотермных) животных. Холоднокровные животные. У холоднокровных животных при изменении температуры окружающей среды меняется температура тела, что приводит к изменению всех процессов жизнедеятельности. В частности, температура влияет на энергетический обмен, но по- разному у холоднокровных и теплокровных животных. У холоднокровных температура тела, а следовательно, и скорость процессов жизнедеятельности (двигательная активность, темпы развития и роста, пищеварение, частота дыхательных движений и пр.) зависят от температуры окружающей среды. Подчиняется этому универсальному правилу и энергетический обмен. Это означает, что по мере увеличения температуры окружающей среды интенсивность дыхания (основной показатель энергетического обмена) непрерывно возрастает. И только в зоне критических для данного животного температур интенсивность дыхания перестает увеличиваться и даже снижается, что связано с ее повреждающим воздействием на организм. Необходимо отметить, что процессы усиления интенсивности дыхания при увеличении температуры среды носят достаточно строгий количественный характер: подчиняясь экспоненциальной зависимости (рис.1). Такой вид температурной зависимости дыхания впервые установил известный датский физиолог Август Крог, лауреат Нобелевской премии 1920 года, открывший механизм капиллярного кровообращения. Работы Крога по влиянию температуры на интенсивность дыхания у рыб были выполнены еще в начале века. Впоследствии проводилось много исследований на различных холоднокровных организмах – от простейших до амфибий и рептилий, обитающих в разных температурных условиях (в тропиках, Арктике, Антарктике, умеренном поясе), и в подавляющем большинстве случаев была получена сходная с результатами Крога зависимость интенсивности дыхания от температуры. Эта зависимость получила в научной литературе название "нормальная кривая Крога". Для холоднокровных закономерна прямая зависимость от температуры уровня метаболизма и, в частности, энергетического обмена. При этом ускорение интенсивности энергетического обмена под воздействием температуры подчиняется общему для всех химических реакций правилу Вант-Гоффа. В соответствии с этим правилом при увеличении температуры на 100 C скорость реакции возрастает в 2-3 раза. Эта количественная зависимость справедлива как для химических реакций, так и для различных метаболических превращений в организме, несмотря на необычайную сложность последних. Поскольку у холоднокровных животных интенсивность дыхания непрерывно возрастает по мере увеличения температуры, на основе этой зависимости нельзя выявить зону оптимальных температур, как это делается в случае теплокровных организмов. Несмотря на постоянное увеличение интенсивности дыхания при возрастании температуры окружающей среды, у холоднокровных должна существовать по аналогии с теплокровными зона оптимальных температур. Чтобы энергетический обмен использовать для определения оптимальных температур, был разработан принцип определения суммарной скорости дыхания (Зотин, Озернюк, 1966). Эти работы были выполнены в Инстетуте биологии развития им. Кольцова АН СССР на развивающейся икре рыб. Сущность данного метода состоит в определении суммарной (за определенную стадию развития) скорости дыхания. Выявлены зоны температур, при которых суммарная скорость дыхания минимальная. При низких и более высоких температурах суммарная скорость дыхания возрастает. Минимальная суммарная скорость при определенной температуре свидетельствует о том, что в этом температурном диапазоне на процессы развития расходуется минимальное количество энергии, и, следовательно, с точки зрения энергетического обмена эти температуры оптимальны. Таким образом, развитие холоднокровных животных протекает в таких температурных условиях, при которых энерготраты минимальные, а выживаемость максимальная. Это естественно, поскольку дефицит энергии ограничивает протекание многих реакций в организме, что может привести к нарушению в развитии, а иногда и к гибели. Связь биоэнергетики и оптимальных условий развития предполагалась давно. Идея о том, что у холоднокровных животных в оптимальных температурных условиях энерготраты должны быть минимальными, высказывалась известным советским экологом Г. Г. Винбергом еще в 1936 году. Результаты исследований минимального уровня энергетического обмена холоднокровных животных в определенных значениях внешних условий (температуры, солености и др.) послужили основой для разработки принципа минимума в онтогенезе (Озернюк, 1988). Сущность данного принципа в том, что развитие организма в таких экологических условиях, где энерготраты минимальные, а выживаемость максимальная, является наиболее вероятной траекторией онтогенеза, "каналом", по которому протекает развитие (рис.2). Этот принцип справедлив, по-видимому, и для теплокровных животных. Теплокровные животные. У теплокровных организмов (птиц и млекопитающих) зависимость энергетического обмена от температуры иная, нежели у холоднокровных. Связано это с наличием терморегуляции у теплокровных. Как меняется основной показатель энергетического обмена – теплопродукция или интенсивность дыхания у теплокровных животных при изменении температуры окружающей среды? По мере увеличения температуры среды температура тела у них не меняется, а интенсивность дыхания снижается (рис.3). При достижении оптимальных температур этот показатель биоэнергетики становится минимальным. Область температур, в которой наименьшая интенсивность дыхания (термонейтральная зона), - наиболее комфортная для данных организмов. При дальнейшем увеличении температуры внешней среды интенсивность дыхания снова возрастает. При этом температура тела не меняется. Какие конкретно механизмы поддерживают постоянную температуру тела телокровных животных? Как усиливается энергетический обмен при понижении температуры среды и увеличивается теплоотдача при повышении внешней температуры? В первом случае, когда речь идет о химической терморегуляции, постоянная температура тела поддерживается за счет рефлекторного усиления энергетического обмена, что приводит к увеличению теплообразования. Этот процесс регулируется при помощи центральной нервной системы: когда температура окружающей среды опускается ниже термонейтральной зоны, рефлекторно усиливается обмен веществ до тех пор, пока не произойдет уравновешивание теплообразования в организме и теплоотдачи в окружающую среду. Вследствие этого температура тела у теплокровных при понижении внешней температуры сохраняется на неизменном уровне. Температурный диапазон от термонейтральной области в сторону понижающихся температур называют зоной химической терморегуляции. Терморегуляторное теплообразование у птиц и млекопитающих связано главным образом со скелетной мускулатурой. Повышение теплообразования при охлаждении организма происходит за счет "терморегуляторного тонуса" – микросокращений внешне неподвижных мышц. При этом дыхание животного может увеличиваться на 150%. Дальнейшее охлаждение организма приводит к подключению еще одного механизма терморегуляторного теплообразования с участием сократительной деятельности мышц – холодовой дрожи. В этих случаях дыхание животного может возрастать в 3-4 раза. Вот почему мы дрожим на холоде – мы согреваемся. При продолжительном охлаждении изменяются энергетические процессы, связанные непосредственно с синтезом АТФ. В таких случаях процесс дыхания связан не с синтезом АТФ, а со свободным (нефосфорилирующим) окислением, приводящим к образованию тепла. Механизмы свободного окисления и их роль в терморегуляции на примере птиц (холодовое разобщение окислительного фосфорилированиря) всесторонне изучены В.П.Скулачевым. Один из источников теплообразования у млекопитающих – бурая жировая ткань. Она обильно снабжена кровеносными сосудами и содержит много митохондрий. Каким образом происходит теплообразование в бурой жировой ткани? При охлаждении животного усиливается кроврснабжение и интенсивность дыхания этой ткани, что приводит к возрастанию теплообразования. Локализована эта ткань в районе грудной клетки и шеи – рядом с жизненно важными органами. Предполагается, что теплообразование за счет бурой жировой ткани используется организмом для быстрого разогрева организма, например, при выходе млекопитающих из состояния спячки. Физическая терморегуляция контролирует теплоотдачу и тем самым участвует в поддержании постоянной температуры тела при высокой температуре внешней среды (за счет испарения влаги с поверхности тела – потоотделении – и с верхних дыхательных путей – учащение поверхностного дыхания). Ныряющие животные. Интересно изменяется газообмен у наземных ныряющих животных. Они вторично перешли к водному образу жизни и выработали в процессе эволюции удивительно эффективные приспособления к жизни в воде. Среди млекопитающих – это китообразные (киты, кашалоты, дельфины) и ластоногие (моржи, тюлени). Среди птиц – это утиные, буревестники, пингвины. К ним относятся также и некоторые рептилии (водные черепахи, морские змеи). Все эти животные в обычных условиях дышат легкими. Однако при нырянии прекращается легочное дыхание, а значит и поступление кислорода в организм. Тем не менее, эти животные довольно долго могут находиться под водой без ущерба для своего состояния: дельфины – до 15 мин., кашалоты – до 1,5 ч., а киты – до 1 ч., ныряя при этом на очень большие глубины. Во время ныряния прежде всего уменьшается концентрация кислорода и увеличивается концентрация СО2 в крови. Каким образом организм выходит из этой кризисной ситуации? Перед нырянием в его крови и в мышцах запасается кислород. Кроме того, в процессе ныряния экономно расходуется кислород, что достигается перестройкой деятельности некоторых систем организма. У ныряющих животных выше эффективность извлечения кислорода из вдыхаемого воздуха. Однако запасание кислорода в легких этих животных при нырянии играет второстепенную роль. Более важно в данном случае запасание кислорода в крови и в мышцах. Так, у ныряющих животных общее количество крови, а также ее кислородная емкость (способность гемоглобина крови связываться с кислородом) выше, чем у неныряющих. Важную роль в запасании кислорода при нырянии играет мускулатура. В мышцах ныряющих животных содержится большое количество белка миоглобина, который, как и гемоглобин, способен связываться с кислородом (красный цвет части мышц у животных определяется содержанием в них этого белка). У миоглобина сродство к кислороду в 10 раз выше, чем у гемоглобина. Поэтому содержащийся в мышцах миоглобин способен запасать большое количество кислорода, а при нырянии в условиях кислордного дефицита отдавать его для нужд организма. Приспособления к большим высотам. Значительная перестройка энергетического обмена наблюдается при адаптации к большим высотам над уровнем моря. На больших высотах процентный состав воздуха не меняется, но снижается барометрическое давление, в результате чего возникает гипоксия – кислородная недостаточность. Для поддержания нормального функционирования организм в таких условиях должен уметь компенсировать нехватку кислорода. Достигается это несколькими путями. Прежде всего, это увеличение вентиляции легких, усиление деятельности сердца, возрастание кислордной емкости крови – способность связывать кислород и переносить его к различным тканям и органам. Так, у человека на высоте 4000 м над уровнем моря число эритроцитов в 1мм3 крови возрастает с 5 до 8 млн. При высокой гипоксии расширяются кровеносные сосуды мозга и сердца. Возрастает способность тканей и клеток извлекать кислород из крови. В условиях дефицита кислорода увеличивается доля образования АТФ за счет гликолиза – анаэробного пути энергетического обмена. В этих условиях возрастает дыхательная поверхность легких, увеличивается емкость грудной клетки и мощность дыхательной мускулатуры. Увеличивается и масса сердца, емкость его кровеносных сосудов, повышается масса митохондрий в миокарде. Таким образом, биоэнергетике отводится важная роль в эволюционном процессе. В 1934 году А. Н. Северцовым была сформулирована идея морфофизиологического прогресса (ароморфозов) – комплекса морфологических и функциональных признаков организмов, усложняющихся в процессе эволюции. Северцов полагал, что основной результат морфофизиологического прогресса – это "увеличение энергии жизнедеятельности", что дает организму важные преимущества в борьбе за существование. Я уже отмечала, что в процессе эволюции наблюдается тенденция к увеличению уровня биоэнергетики животных различных систематических групп. У простейших энергетический обмен ниже, чем у многоклеточных беспозвоночных животных; у позвоночных холоднокровных выше, чем у беспозвоночных, и наконец, у теплокровных позвоночных намного выше, чем у холоднокровных позвоночных. Однако внутри каждой из этих групп выраженной тенденции к увеличению уровня биоэнергетики по мере усложнения организации не наблюдается, и связано это главным образом с особенностями образа жизни. У организмов внутри каждой из перечисленных выше групп энергетический обмен определяется в основном двигательной активностью. Таким образом, особенности экологии накладывают серьезный отпечаток на эволюционные закономерности биоэнергетики животных.