

Гормони гіпоталамусу, їх структура. Роль ліберинів і статинів в регуляції функцій аденогіпофізу.
К настоящему времени в гипоталамусе открыто 7 стимуляторов и 3 ингибитора секреции гормонов гипофиза, а именно: кортиколиберин, тиролиберин, люлиберин, фоллилиберин, соматолиберин,пролактолиберин, меланолиберин, соматостатин, пролактостатин и меланостатин. В чистом виде выделено 5 гормонов, для которых установлена первичная структура, подтвержденная химическим синтезом.
Большие трудности при получении гормонов гипоталамуса в чистом виде объясняются чрезвычайно низким содержанием их в исходной ткани.
По химическому строению все гормоны гипоталамуса являются низкомолекулярными пептидами, так называемыми олигопептидами необычного строения, хотя точный аминокислотный состав и первичная структура выяснены не для всех. Приводим полученные к настоящему времени данные о химической природе шести из известных 10 гормонов гипоталамуса.
1. Тиролиберин (Пиро-Глу–Гис–Про–NH2):
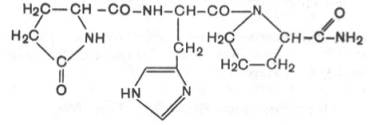
Тиролиберин представлен трипептидом, состоящим из пироглутаминовой (циклической) кислоты, гистидина и пролинамида, соединенных пептидными связями. В отличие от классических пептидов он не содержит свободных NH2- и СООН-групп у N- и С-концевых аминокислот.
Обеспечивает высвобождение тиротропного гормона (ТТГ) из передней доли гипофиза.
2. Гонадолиберин является декапептидом, состоящим из 10 аминокислот последовательности:
Пиро-Глу–Гис–Трп–Сер–Тир–Гли–Лей–Арг–Про–Гли-NН2
Концевая С-аминокислота представлена глицинамидом.
Обеспечивает высвобождение ФСГ, ЛГ и пролактина
3. Соматостатин является циклическим тетрадекапептидом (состоит из 14 аминокислотных остатков) :

Отличается этот гормон от двух предыдущих, помимо циклической структуры, тем, что не содержит на N-конце пироглутаминовой кислоты: дисульфидная связь образуется между двумя остатками цистеина в 3-м и 14-м положениях. Следует отметить, что синтетический линейный аналог соматостатина также наделен аналогичной биологической активностью, что свидетельствует о несущественности дисульфидного мостика природного гормона. Помимо гипоталамуса, соматостатин продуцируется нейронами центральной и периферической нервных систем, а также синтезируется в S-клетках панкреатических островков (островков Лангерганса) в поджелудочной железе и клетках кишечника. Он оказывает широкий спектр биологического действия; в частности, показано ингибирующее действие на синтез гормона роста в аденогипофизе, а также прямое тормозящее действие его на биосинтез инсулина и глюкагона в β- и α-клетках островков Лангерганса.
4. Соматолиберин. Он представлен 44 аминокислотными остатками с полностью раскрытой последовательностью. Биологической активностью соматолиберина наделен, кроме того, химически синтезированный декапептид:
Н-Вал–Гис–Лей–Сер–Ала–Глу–Глн–Лиз–Глу–Ала-ОН.
Этот декапептид стимулирует синтез и секрецию гормона роста гипофиза соматотропина.
5. Меланолиберин, химическая структура которого аналогична структуре открытого кольца гормона окситоцина (без трипептидной боковой цепи), имеет следующее строение:
Н-Цис–Тир–Иле–Глн–Асн–Цис-ОН.
6. Меланостатин (меланотропинингибирующий фактор) представленили трипептидом: Пиро-Глу–Лей–Гли-NН 2 , или пентапептидом со следующей последовательностью:
Пиро-Глу–Гис–Фен–Aрг–Гли–NН2.
Н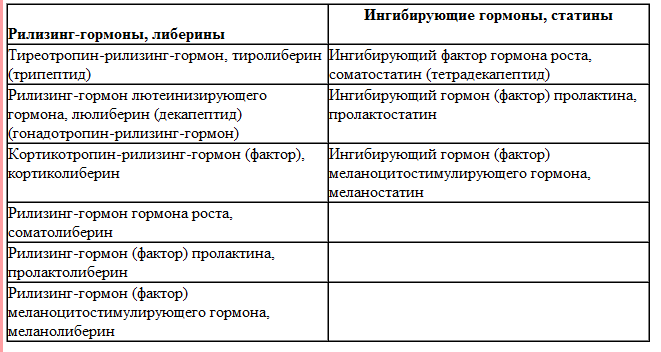 еобходимо
отметить, что меланолиберин оказывает
стимулирующее действие, а меланостатин,
напротив, ингибирующее действие на
синтез и секрецию меланотропина в
передней доле гипофиза.
еобходимо
отметить, что меланолиберин оказывает
стимулирующее действие, а меланостатин,
напротив, ингибирующее действие на
синтез и секрецию меланотропина в
передней доле гипофиза.
Либерины (рилизинг—гормоны) стимулируют, а статины подавляют секрецию гормонов аденогипофиза
Роль гормонів в інтеграції внутрішньоклітинного метаболізму і регуляції фізіологічних функцій організму. Клітинні механізми дії цих гормонів.
Гормони гіпофіза. Гормони передньої, середньої та задньої долі гіпофіза. Класифікація гормонів аденогіпофізу.
СТРОМА капсула, прослойки соединительной ткани в передней доле образованы рыхлой влолкнистой соединительной тканью
ПАРЕНХИМА состоит из передней, промежуточной (средней) и задней долей
передняя и средняя доли образуют аденогипофиз, а задняя доля называется нейрогипофизом
ПЕРЕДНЯЯ ДОЛЯ состоит из клеток - аденоцитов, они подразделяются на:
хромофобные клетки - их цитоплазма плохо окрашивается, являются малодифференцированными клетками или представляют собой старые, закончившие свою работу клетки, которые не синтезируют гормонов
хромофильные клетки - их цитоплазма хорошо окрашивается; эти клетки синтезируют гормоны; зависимости от способности окрашиваться ацидофильно или базофильно, а также в зависимости от синтезируемых гормонов хромофильные клетки подразделяются на:
базофильные клетки - цитоплазма окрашивается базофильно (в синий или фиолетовый цвет); среди базофильных клеток различают:
тиреотрофоциты - продуцируют тиреотропный гормон
адренокортикотрофоциты - продуцируют адренокортикотропный гормон
гонадотрофоциты - продуцируют гонадотропные гормоны -лютеинизирующий гормон и фолликулостимулирующий гормон; эти два гормона синтезируются отдельными подтипами гонадотрофоцитов
ацидофильные клетки - цитоплазма окрашивается ацидофильно (в розовый или красный цвет); среди ацидофильных клеток различают:
соматотрофоциты - продуцируют соматотропный гормон или гормон роста
пролактотрофоциты - продуцируют гормон пролактин
В передней доле гипофиза имеются фолликулярно-звездчатые клетки и образованные ими фолликулярные образования, заполненные жидкостью. Полагают, что фолликулярно- звездчатые клетки могут вырабатывать некоторые медиаторы иммунитета, такие как интерлейкин-1, интерлейкин-6, которые могут модулировать выработку гормонов аденоцитами.
ЗАДНЯЯ ДОЛЯ (НЕЙРОГИПОФИЗ) В ней имеются:
Дистальные части аксонов нервных клеток, перикарионы которых лежат в супраоптическом и паравентрикулярном ядрах гипоталамуса. Аксоны закан- чиваются терминальными расширениями, которые называются тельцами Херинга. Терминальные части аксонов образуют с сосудами аксо-вазальные синапсы.
Видоизмененные клетки нейроглии - питуициты, которые поддерживают аксоны нервных клеток.
В перикарионах нервных клеток супраоптических и паравентрикулярных ядер гипоталамуса синтезируются два гормона - окситоцин и вазопрессин. Они транспор- тируются по аксонам в заднюю долю гипофиза, доходят до самых терминальных частей аксона, где есть расширения - тельца Херинга, в которых окситоцин и вазопрессин могут накапливаться и храниться. Далее, окситоцин и вазопрессин прямо из терминальных частей аксонов сразу попадают в кровь, так как аксоны образуют с сосудами аксо-вазальные синапсы.
ПРОМЕЖУТОЧНАЯ ДОЛЯ
В этой доле имеются клетки, продуцирующие меланоцитстимулирующий гормон
I. Гормоны аденогипофиза
А. Классификация, структура и функции
1. Семейство СТГ. К нему относятся СТГ и пролактин, а также гормон, образующийся в плаценте, — плацентарный лактоген. Все эти гормоны состоят из одной негликозилированной полипептидной цепи и характеризуются значительным сходством первичной структуры.
а. СТГ синтезируется в соматотропных клетках, имеет молекулярную массу 22 000 и содержит 191 аминокислоту. Физиологические эффекты СТГ принято разделять на прямые и непрямые. Прямые эффекты СТГ: стимуляция синтеза и секреции ИФР в печени и других органах и тканях, стимуляция липолиза в жировой ткани и стимуляция продукции глюкозы в печени. Непрямые эффекты СТГ — это его рост-стимулирующее и анаболическое действие. Эти эффекты опосредуются ИФР-I. Основным источником ИФР-I является печень. ИФР-I стимулирует рост кости, хряща и мягких тканей. Непрямые эффекты СТГ подавляются глюкокортикоидами.
б. Пролактин синтезируется в лактотропных клетках, имеет молекулярную массу 22 500 и содержит 198 аминокислот. Главная мишень пролактина — молочные железы. Пролактин стимулирует рост молочных желез во время беременности и лактацию после родов. Во время беременности лактогенный эффект пролактина блокируется эстрогенами и прогестероном. Рецепторы пролактина обнаружены в гипоталамусе, печени, яичках, яичниках, но действие пролактина на эти органы изучено недостаточно. Гиперпролактинемия угнетает гипоталамо-гипофизарно-гонадную систему и является частой причиной бесплодия у женщин. Недавно показали, что рецепторы пролактина присутствуют на T-лимфоцитах и что пролактин влияет на иммунные реакции.
2. Семейство гликопротеидных гормонов включает аденогипофизарные гормоны ЛГ, ФСГ и ТТГ, а также плацентарный гормон ХГ. Эти гормоны состоят из двух сильно гликозилированных полипептидных цепей (субъединиц) — альфа и бета. У всех гормонов субъединицы альфа идентичны: они включают по 92 аминокислоты, расположенных в одинаковой последовательности. Напротив, последовательности аминокислот в субъединицах бета различаются. Именно эти различия определяют специфичность действия гликопротеидных гормонов на ткани-мишени. Молекулярная масса ЛГ, ФСГ, ТТГ и ХГ неодинакова и зависит в первую очередь от количества углеводных остатков.
а. ЛГ и ФСГ синтезируются в гонадотропных клетках. У обоих гормонов субъединица бета включает 115 аминокислот, а молекулярная масса составляет соответственно 29 400 и 32 600. ЛГ и ФСГ регулируют синтез и секрецию половых гормонов и гаметогенез.
1) В яичниках ЛГ стимулирует овуляцию и секрецию прогестерона. Рецепторы ЛГ и ХГ присутствуют на клетках внешней оболочки и гранулярного слоя фолликулов и на интерстициальных клетках. ФСГ стимулирует секрецию эстрогенов, рост и созревание фолликулов. Рецепторы ФСГ имеются только на клетках гранулярного слоя.
2) В яичках ЛГ стимулирует секрецию тестостерона. Рецепторы ЛГ и ХГ присутствуют только на клетках Лейдига. ФСГ не влияет на синтез андрогенов, но необходим для сперматогенеза. Рецепторы ФСГ обнаружены только на клетках Сертоли.
б. ТТГ синтезируется в тиреотропных клетках, имеет молекулярную массу 30 500; субъединица бета включает 112 аминокислот. Основная роль ТТГ — стимуляция синтеза тиреоидных гормонов. ТТГ контролирует почти все этапы синтеза, в том числе — присоединение неорганического йода к тиреоглобулину и образование T3 и T4 из моно- и дийодтирозина.
3. Семейство производных проопиомеланокортина. Кортикотропные клетки аденогипофиза секретируют несколько гормонов: АКТГ, альфа- и бета-МСГ, бета- и гамма-липотропины, а также эндорфины. Все эти гормоны содержат гептапептид Мет-Глу-Гис-Фен-Арг-Трп-Гли и образуются из крупной молекулы-предшественника — проопиомеланокортина (молекулярная масса 31 000).
а. АКТГ имеет молекулярную массу 4500 и содержит 39 аминокислот. АКТГ стимулирует синтез гормонов в коре надпочечников, в первую очередь — синтез глюкокортикоидов в пучковой и сетчатой зонах. Выброс АКТГ из кортикотропных клеток или введение большой дозы АКТГ может вызвать кратковременный подъем уровня альдостерона. Еще один эффект АКТГ — стимуляция синтеза меланина в меланоцитах. По-видимому, это служит причиной гиперпигментации при синдроме Нельсона и первичной надпочечниковой недостаточности.
б. Функции других производных проопиомеланокортина изучены хуже. Установлено, что альфа-МСГ стимулирует синтез меланина в меланоцитах, а гамма-МСГ — синтез альдостерона в коре надпочечников. В опытах на культурах клеток коры надпочечников показали, что бета-липотропин стимулирует синтез кортикостероидов, причем эффект бета-липотропина опосредуется рецепторами АКТГ.
Біосинтез та секреція гормонів аденогіпофізу. Вплив гормонів аденогіпофізу на метаболізм та фізіологічні функції.
I. Гормоны аденогипофиза
А. Классификация, структура и функции
1. Семейство СТГ. К нему относятся СТГ и пролактин, а также гормон, образующийся в плаценте, — плацентарный лактоген. Все эти гормоны состоят из одной негликозилированной полипептидной цепи и характеризуются значительным сходством первичной структуры.
а. СТГ синтезируется в соматотропных клетках, имеет молекулярную массу 22 000 и содержит 191 аминокислоту. Физиологические эффекты СТГ принято разделять на прямые и непрямые. Прямые эффекты СТГ: стимуляция синтеза и секреции ИФР в печени и других органах и тканях, стимуляция липолиза в жировой ткани и стимуляция продукции глюкозы в печени. Непрямые эффекты СТГ — это его рост-стимулирующее и анаболическое действие. Эти эффекты опосредуются ИФР-I. Основным источником ИФР-I является печень. ИФР-I стимулирует рост кости, хряща и мягких тканей. Непрямые эффекты СТГ подавляются глюкокортикоидами.
б. Пролактин синтезируется в лактотропных клетках, имеет молекулярную массу 22 500 и содержит 198 аминокислот. Главная мишень пролактина — молочные железы. Пролактин стимулирует рост молочных желез во время беременности и лактацию после родов. Во время беременности лактогенный эффект пролактина блокируется эстрогенами и прогестероном. Рецепторы пролактина обнаружены в гипоталамусе, печени, яичках, яичниках, но действие пролактина на эти органы изучено недостаточно. Гиперпролактинемия угнетает гипоталамо-гипофизарно-гонадную систему и является частой причиной бесплодия у женщин. Недавно показали, что рецепторы пролактина присутствуют на T-лимфоцитах и что пролактин влияет на иммунные реакции.
2. Семейство гликопротеидных гормонов включает аденогипофизарные гормоны ЛГ, ФСГ и ТТГ, а также плацентарный гормон ХГ. Эти гормоны состоят из двух сильно гликозилированных полипептидных цепей (субъединиц) — альфа и бета. У всех гормонов субъединицы альфа идентичны: они включают по 92 аминокислоты, расположенных в одинаковой последовательности. Напротив, последовательности аминокислот в субъединицах бета различаются. Именно эти различия определяют специфичность действия гликопротеидных гормонов на ткани-мишени. Молекулярная масса ЛГ, ФСГ, ТТГ и ХГ неодинакова и зависит в первую очередь от количества углеводных остатков.
а. ЛГ и ФСГ синтезируются в гонадотропных клетках. У обоих гормонов субъединица бета включает 115 аминокислот, а молекулярная масса составляет соответственно 29 400 и 32 600. ЛГ и ФСГ регулируют синтез и секрецию половых гормонов и гаметогенез.
1) В яичниках ЛГ стимулирует овуляцию и секрецию прогестерона. Рецепторы ЛГ и ХГ присутствуют на клетках внешней оболочки и гранулярного слоя фолликулов и на интерстициальных клетках. ФСГ стимулирует секрецию эстрогенов, рост и созревание фолликулов. Рецепторы ФСГ имеются только на клетках гранулярного слоя.
2) В яичках ЛГ стимулирует секрецию тестостерона. Рецепторы ЛГ и ХГ присутствуют только на клетках Лейдига. ФСГ не влияет на синтез андрогенов, но необходим для сперматогенеза. Рецепторы ФСГ обнаружены только на клетках Сертоли.
б. ТТГ синтезируется в тиреотропных клетках, имеет молекулярную массу 30 500; субъединица бета включает 112 аминокислот. Основная роль ТТГ — стимуляция синтеза тиреоидных гормонов. ТТГ контролирует почти все этапы синтеза, в том числе — присоединение неорганического йода к тиреоглобулину и образование T3 и T4 из моно- и дийодтирозина.
3. Семейство производных проопиомеланокортина. Кортикотропные клетки аденогипофиза секретируют несколько гормонов: АКТГ, альфа- и бета-МСГ, бета- и гамма-липотропины, а также эндорфины. Все эти гормоны содержат гептапептид Мет-Глу-Гис-Фен-Арг-Трп-Гли и образуются из крупной молекулы-предшественника — проопиомеланокортина (молекулярная масса 31 000).
а. АКТГ имеет молекулярную массу 4500 и содержит 39 аминокислот. АКТГ стимулирует синтез гормонов в коре надпочечников, в первую очередь — синтез глюкокортикоидов в пучковой и сетчатой зонах. Выброс АКТГ из кортикотропных клеток или введение большой дозы АКТГ может вызвать кратковременный подъем уровня альдостерона. Еще один эффект АКТГ — стимуляция синтеза меланина в меланоцитах. По-видимому, это служит причиной гиперпигментации при синдроме Нельсона и первичной надпочечниковой недостаточности.
б. Функции других производных проопиомеланокортина изучены хуже. Установлено, что альфа-МСГ стимулирует синтез меланина в меланоцитах, а гамма-МСГ — синтез альдостерона в коре надпочечников. В опытах на культурах клеток коры надпочечников показали, что бета-липотропин стимулирует синтез кортикостероидов, причем эффект бета-липотропина опосредуется рецепторами АКТГ.
Гормоны аденогипофиза: секреция, регуляция
Система обратной связи. ТТГ , АКТГ и оба гонадотропных гормона - ЛГ и ФСГ - стимулируют секрецию гормонов в эндокринных железах-мишенях. В свою очередь, гормоны желез-мишеней подавляют секрецию соответствующих аденогипофизарных гормонов. Например, повышение уровня кортизола в крови тормозит секрецию АКТГ. Такие же связи существуют между тиреоидными гормонами и ТТГ , между половыми гормонами и гонадотропными гормонами . Гормоны, подавляющие секрецию СТГ и пролактина , пока не обнаружены, хотя недавно было установлено, что ИФР-I тормозит секрецию СТГ.
Либерины и статины . Секреторная активность клеток аденогипофиза зависит не только от уровня гормонов эндокринных желез-мишеней. Важнейшую роль в регуляции секреции аденогипофизарных гормонов играет гипоталамус . В ядрах гипоталамуса образуются пептидные гормоны - либерины и статины , поступающие в воротную систему гипофиза:
- Тиролиберин стимулирует секрецию ТТГ и пролактина .
- Гонадолиберин стимулирует секрецию ЛГ и ФСГ .
- Кортиколиберин стимулирует секрецию АКТГ , МСГ и бета-липотропина .
- Соматолиберин стимулирует секрецию СТГ .
- Соматостатин подавляет секрецию СТГ и, в меньшей степени, ТТГ.
- Дофамин подавляет секрецию пролактина и, по-видимому, является главным физиологическим регулятором секреции этого гормона.
Гормоны периферических эндокринных желез могут подавлять не только секрецию соответствующих тропных гормонов аденогипофиза, но и секрецию либеринов гипоталамуса . Например, эстрогены подавляют как секрецию ЛГ и ФСГ , так и секрецию гонадолиберина .
Гормони нейрогіпофізу. Окситоцин і вазопресин. Клітинні та молекулярні механізми дії гормонів нейрогіпофізу.
Гормоны нейрогипофиза. Антидиуретический гормон (АДГ). В общем виде действие АДГ сводится к двум основным эффектам:
1) стимулируется реабсорбция воды в дистальных канальцах почек. В результате увеличивается объем циркулирующей крови, повышается АД, снижается диурез и возрастает относительная плотность мочи. В результате усиленного обратного всасывания воды снижается осмотическое давление межклеточной жидкости. Под действием АДГ происходит активация фермента аденилатциклазы, локализующегося на поверхности базолатеральной (обращенной к интерстицию) мембраны клеток эпителия почечных канальцев. Активация аденилатциклазы приводит к накоплению в цитоплазме этих клеток цАМФ. Последний диффундирует в область апикальной (обращенной в просвет почечного канальца) мембраны и стимулирует образование в цитоплазме белковых везикул, которые затем включаются в структуру апикальной мембраны и образуют в ней каналы, высокопроницаемые для воды. В результате вода из просвета почечных канальцев поступает в цитоплазму клеток эпителия канальцев, перемещается к базолатеральной мембране и, проникая через нее, попадает в интерстициальную ткань. После разрушения АДГ белковые везикулы элиминируются из структуры апикальной мембраны. В результате этого последняя становится непроницаемой для
воды;
2) в больших дозах АДГ вызывает сужение артериол, что приводит к увеличению АД. Развитию гипертензии способствует также наблюдающееся под влиянием АДГ повышение чувствительности сосудистой стенки к констрикторному действию катехоламинов. В связи с тем, что введение АДГ приводит к повышению АД, этот гормон получил также название «вазопрессин». Однако поскольку эффект вазоконстрикции возникает только при действии больших доз АДГ, то считают, что в физиологических условиях значимость его вазоконстрикторного влияния невелика. С другой стороны, развитие вазоконстрикции может иметь существенное адаптивное значение при некоторых патологических состояниях, например при острой кровопотере, сильных болевых воздействиях, поскольку в этих условиях в крови может присутствовать большое количество АДГ.
Основная часть АДГ синтезируется в супраоптическом ядре гипоталамуса (примерно 5/6 от общего количества), меньшая часть — в паравентрикулярном ядре. Секреция этого гормона усиливается при повышении осмотического давления крови. Последнее можно продемонстрировать путем введения гипертонического раствора в сосуды, питающие гипоталамус. В этом случае происходит раздражение осморецепторов, что приводит к увеличению выработки гормона в супраоптическом и паравентрикулярном ядрах и повышенной его секреции из задней доли гипофиза в кровь. Важным стимулом для регуляции секреции АДГ является также изменение объема циркулирующей крови. Показано, что при снижении последнего на 15—20% количество образующегося АДГ может увеличиваться в несколько десятков раз. В этом случае интенсивность секреции гормона меняется в зависимости от характера информации, поступающей в гипоталамус от волюморецепторов, реагирующих на растяжение кровью и локализующихся в правом предсердии, и барорецепторов, расположенных в аортальной и синокаротидной зонах, а также в легочной артерии.
Недостаточная секреция АДГ приводит к развитию несахарного мочеизнурения (diabetes insipidus), основными проявлениями которого являются сильная жажда (полидипсия) и потеря большого количества жидкости с выделяемой мочой (полиурия). Наблюдается учащенное мочеиспускание (поллакиурия), в результате которого больной за сутки выделяет до 10—20 л мочи низкой относительной плотности. Симптомы этого заболевания проходят при введении синтетического вазопрессина или препаратов, приготовленных из задней доли гипофиза животных.
Окситоцин. Эффекты этого гормона реализуются главным образом в двух направлениях:
1) окситоцин вызывает сокращение гладкой мускулатуры матки. Установлено, что при удалении гипофиза у животных родовые схватки становятся длительными и малоэффективными. Таким образом, окситоцин является гормоном, обеспечивающим нормальное протекание родового акта (отсюда произошло и его название — от лат. oxy — сильный, tokos — роды). Адекватное проявление этого эффекта возможно при условии достаточной концентрации в крови эстрогенов, которые усиливают чувствительность матки к окситоцину;
2) окситоцин принимает участие в регуляции процессов лактации. Он усиливает сокращение миоэпителиальных клеток в молочных железах и тем самым способствует выделению молока.
Содержание окситоцина в крови возрастает в конце беременности, в послеродовом периоде. Кроме того, его продукция стимулируется рефлекторно при раздражении соска в процессе грудного вскармливания.
Механизм действия окситоцина неизвестен. Он вызывает сокращение гладких мышц матки и поэтому используется в фармакологических дозах для стимуляции родовой деятельности у женщин ( рис. 10-4сер ).
Интересно, что у беременных животных с поврежденной гипоталамо-гипофизарной системой вовсе не обязательно возникают нарушения родовой деятельности.
Наиболее вероятная физиологическая функция окситоцина заключается в стимуляции сокращений миоэпителиальных клеток, окружающих альвеолы молочной железы. Это вызывает перемещение молока в систему альвеолярных протоков и приводит к его выбросу ( рис. 10-5сер ).
Мембранные рецепторы окситоцина найдены в тканях матки и молочной железы . Их количество возрастает под действием эстрогенов и снижается под влиянием прогестерона . Наступление лактации до родов можно, очевидно, объяснить одновременным повышением количества эстрогенов и падением уровня прогестерона непосредственно перед родами. Производные прогестерона часто используются для подавления послеродовой лактации у женщин. Окситоцин и нейрофизин I , по-видимому, образуются и в яичниках , где окситоцин может ингибировать стероидогенез .
Химические группы, существенные для действия окситоцина, включают первичную аминогруппу N-концевого цистеина, фенольную группу тирозина, 3 карбоксамидные группы аспарагина, глутамина и глицинамида, дисульфидную (S-S) связь. Путем удаления или замещения этих групп получены многочисленные аналоги окситоцина. Например, удаление свободной первичной аминогруппы концевого остатка полуцистеина (положение 1) приводит к образованию дезаминоокситоцина , антидиуретическая активность которого в 4-5 раз превышает активность природного окситоцина.