

Тема 4. Лимитированный рост популяции. Хемостат. Экономический коэффициент и траты на поддержание
Теория: Данный раздел теории разработан в основном для популяций микроорганизмов, однако при соблюдении определенных условий и постоянстве внешних параметров он приложим также для описания динамики популяций многоклеточных организмов.
Экономический коэффициент. Траты на поддержание: Для описания и сравнения эффективности утилизации субстрата S при росте организмов используется так называемый экономический коэффициент Y, который определяется как
![]() ,
(4.1)
,
(4.1)
где X и S соответственно концентрации биомассы и изучаемого субстрата. Данный параметр отражает прирост биомассы на единицу потребляемого субстрата и является важнейшей характеристикой ростовых процессов. По причинам, изложенным ниже, он также называется суммарным экономическим коэффициентом. Размерность экономического коэффициента зависит от применяемых размерностей концентрации биомассы и субстрата.
Если лимитирующий субстрат является источником углерода и энергии для процессов биосинтеза, то его значение, как правило, меньше единицы. Данный субстрат при потреблении затрачивается на два типа процессов. Первый тип процессов связан с собственно ростом и размножением. В этом случае эффективность потребления энергетического субстрата характеризуют так называемым ростовым экономическим коэффициентом Yрост. Размерность данного параметра такая же, что и у определенного выше суммарного экономического коэффициента. Второй тип процессов связан с затратами на поддержание уже имеющейся биомассы – сохранением всех трансмембранных градиентов, восстановлением постоянно разрушающихся биополимеров, биомембран и т.д. В физиологии это называют основным обменом. Количественно траты энергетического субстрата на эти процессы характеризуют параметром m, называемым траты на поддержание. Размерность этого параметра – [время-1]. Суммарный экономический коэффициент Y, ростовой экономический коэффициент Yрост и траты на поддержание m согласно теории связаны следующим соотношением (Перт, 1978):
![]() ,
(4.2)
,
(4.2)
где r - удельная скорость роста биомассы популяции. Соответственно, кинетика потребления субстрата на рост и поддержание биомассы описывается уравнениями:
![]() .
(4.3)
.
(4.3)
Динамика лимитированного роста популяции: Выделяют два типа процессов лимитированного роста популяций. В первом из них субстрат вносится в систему однократно, и далее он в процессе роста только потребляется. В микробиологии и биотехнологии такой процесс называют периодическим культивированием. Во втором процессе, который называется непрерывным культивированием, в ходе роста популяции субстрат постоянно добавляется в систему, при этом изымается избыток биомассы. Система, в которой скорости подачи субстрата и изъятия биомассы постоянны, называется хемостат (Перт, 1978; Smith, Waltman, 1995). Поскольку динамические уравнения, описывающие данные системы, несколько отличаются, рассмотрим их отдельно.
Периодическое культивирование: Динамика популяции в условиях периодического культивирования описывается следующей системой уравнений, известных как модель Моно (Перт, 1978; Варфоломеев, Гуревич, 1999):
 (4.4)
(4.4)
Здесь X – концентрация биомассы, S - концентрация лимитирующего субстрата, t – время, rmax - максимальная удельная скорость роста, KS - константа насыщения, Y - экономический коэффициент. Система (4.4) имеет аналитическое решение вида (Перт, 1978; Варфоломеев, Гуревич, 1999):
![]() .
(4.5)
.
(4.5)
Здесь S0 и X0 - начальные концентрации субстрата и биомассы соответственно. На практике часто применяется также первый интеграл системы (4.4):
![]() (4.6)
(4.6)
Непрерывное культивирование. Хемостат: Для описания лимитированного роста популяции в хемостате применяют следующую систему уравнений (Перт, 1978; Варфоломеев, Гуревич, 1999):
 (4.7)
(4.7)
Обозначения здесь те же, что и в системе (4.4), параметр D называется протоком, он численно равен скорости подачи питательной среды (субстрата) в культиватор (ферментер), нормированной на эффективный объем культиватора. Размерность протока – [время-1] . На практике часто используют соотношения, получаемые из анализа стационарного состояния хемостата (Перт,1978):
![]() . (4.8)
. (4.8)
Здесь
![]() -
стационарная удельная скорость роста
популяции,
-
стационарная удельная скорость роста
популяции,
![]() и
и
![]() -
стационарные концентрации субстрата
и биомассы.
-
стационарные концентрации субстрата
и биомассы.
Задача 4.1
Условия: Культуру клеток кишечной палочки E.coli выращивали в аэробных условиях на бедной минеральной среде с глюкозой, содержащей в качестве единственного источника азота хлорид аммония в лимитирующих рост концентрациях. В ходе опыта измеряли концентрацию биомассы клеток и концентрацию лимитирующего субстрата, хлорида аммония (Керридж, Типтон, 1974). Данные представлены в таблице 4.1.
Таблица 4.1.
Концентрация хлорида аммония, S, г/л |
Концентрация биомассы клеток, X, г/л |
S0 – S г/л |
X – X0 г/л |
0,10 |
0,08 |
0,00 |
0,00 |
0,25 |
0,22 |
0,15 |
0,14 |
0,40 |
0,32 |
0,30 |
0,25 |
0,50 |
0,43 |
0,40 |
0,35 |
0,60 |
0,49 |
0,50 |
0,41 |
0,70 |
0,59 |
0,60 |
0,51 |
Вопросы: Определить по представленным данным экономический коэффициент Y для хлорида аммония у исследуемой популяции.
Решение: Напомним, что экономический коэффициент Y выражает количественные потребности организмов (клеток) в пище и равен по определению Y = dX/dS, где X и S соответственно концентрации биомассы и изучаемого субстрата. В условиях лимитированного роста популяции, когда субстрат вводится в систему однократно в начале опыта, экономический коэффициент связывает концентрацию субстрата и концентрацию биомассы клеток (организмов) соотношением (4.6):
Используем данное уравнение для решения поставленной задачи. Для этого рассчитаем вспомогательные переменные X – X0 и S0 – S, как это указано в таблице 4.1 (3 и 4 столбцы). Далее построим график в этих вспомогательных координатах, используя данные из 3 и 4 столбцов таблицы 4.1 (рис.4.1).

Рис.4.1. Определение экономического коэффициента Y
по данным из таблицы 4.1.
Экспериментальные данные, как видно, группируются около прямой линии, тангенс угла которой равен искомому экономическому коэффициенту. Таким образом, экономический коэффициент для хлорида аммония у исследуемой популяции кишечной палочки равен : Y = 0.834 г биомассы на 1 г хлорида аммония.
Задача 4.2
Условия: С целью определения кинетических параметров, KS и rmax, и экономического коэффициента Y, характеризующих рост популяции кишечной палочки E.coli на глюкозе, проводили периодическое культивирование клеток E.coli на бедной минеральной среде с лимитированием по данному субстрату до выхода популяции в стационарную фазу. В ходе эксперимента измеряли концентрацию биомассы клеток и концентрацию глюкозы.
Результаты (в интервале от начала экспоненциальной фазы роста до фиксирования стационарного состояния) представлены в табл. 4.2 и на рис.4.2.
Таблица 4.2.
Время,t час |
Концентрация биомассы, X мг/л |
Концентрация глюкозы, S мг/л |
S0 - S |
X– X0 |
|
|
0 |
20 |
2000 |
0 |
0 |
|
|
0,5 |
30,52 |
1976 |
23,4 |
10,5 |
0,845299 |
-0,023515 |
1 |
46,6 |
1940 |
59,1 |
26,6 |
0,845868 |
-0,030001 |
1,5 |
71,1 |
1886 |
113,5 |
51,1 |
0,845570 |
-0,038968 |
2 |
108,5 |
1803 |
196,6 |
88 |
0,845508 |
-0,051755 |
2,5 |
165,5 |
1676 |
323,3 |
145 |
0,845295 |
-0,070535 |
3 |
252,5 |
1483 |
516,6 |
232 |
0,845226 |
-0,099618 |
3,5 |
385 |
1188 |
811,1 |
365 |
0,845003 |
-0,148608 |
4 |
587 |
740 |
1260 |
567 |
0,844823 |
-0,248563 |
4,33 |
773 |
326 |
1673 |
753 |
0,844006 |
-0,418467 |
4,5 |
885 |
77 |
1922 |
865 |
0,842190 |
-0,721565 |
4,56 |
918 |
4,4 |
1995 |
898 |
0,839137 |
-1,339747 |
4.564 |
919 |
2,2 |
1997 |
899 |
0,838608 |
-1,490388 |
5 |
920 |
0 |
2000 |
900 |
|
|
5,5 |
920 |
0 |
2000 |
900 |
|
|
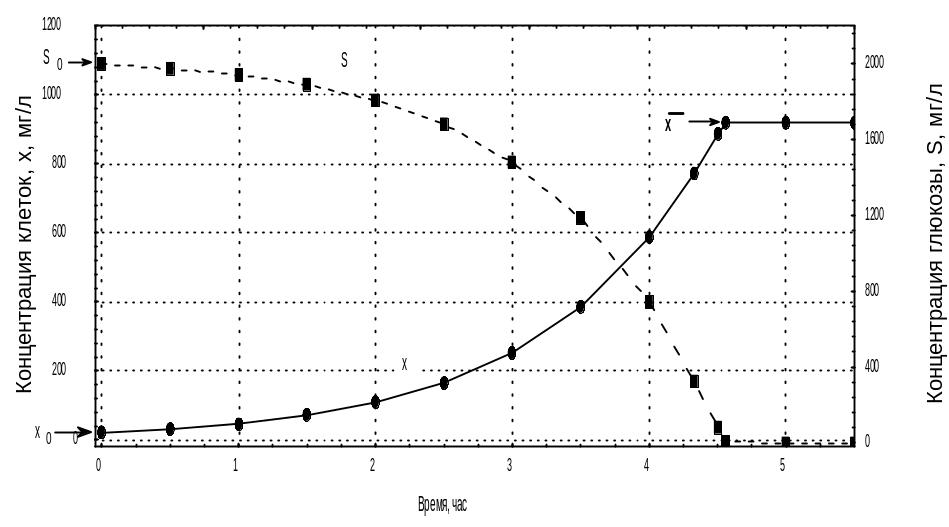
Рис.4.2. Изменение концентрации биомассы, X, и концентрации глюкозы, S, в ходе периодического культивирования популяции E.coli.
Вопросы: Определить по представленным данным значения кинетических параметров, KS и rmax, и экономического коэффициента, Y, характеризующих рост популяции кишечной палочки E.coli на глюкозе.
Решение: Определим на первом этапе значение экономического коэффициента, как это указано в задаче 4.1.

Рис.4.3. Определение экономического коэффициента Y по данным из таблицы 4.2.
Для
этого рассчитаем вспомогательные
переменные S0
–
S
и X
– X0
(таб.4.2, 4 и 5 столбцы) и построим график
в этих координатах, используя данные
из 4 и 5 столбцов таблицы 4.2 (рис.4.3). Тангенс
угла наклона регрессионной прямой,
проведенной через экспериментальные
точки, дает значение экономического
коэффициента: Y
= 0.45. Проверим соответствие данного
значения экономического коэффициента
экспериментальным данным. Согласно
теории стационарное значение концентрации
биомассы
![]() должно удовлетворять соотношению
должно удовлетворять соотношению
![]() .
Используя данные из таб.4.2 получаем:
.
Используя данные из таб.4.2 получаем:
![]() =
920 мг/л,
=
920 мг/л,
![]() =20мг/л,
=20мг/л,
![]() =2000мг/л
=2000мг/л
![]() 920 = 0.45*2000 + 20 = 920. Соответствие налицо.
920 = 0.45*2000 + 20 = 920. Соответствие налицо.
Для определения кинетических параметров, KS и rmax, используем линейную анаморфозу интегральной формы уравнения клеточного роста (4.5)(Варфоломеев, Гуревич, 1999):
![]() .
(4.9)
.
(4.9)
С
этой целью на основе данных из столбцов
1 и 2 таб.4.2 рассчитаем вспомогательные
переменные
![]() и
и
![]() ,
как это указано в
таб.4.2, столбцы 6 и 7, и построим график в
этих координатах (рис.4.4).
,
как это указано в
таб.4.2, столбцы 6 и 7, и построим график в
этих координатах (рис.4.4).

Рис.4.4. Определение кинетических параметров роста клеток KS и rmax по данным из таб.4.2. и рис.4.2
Как
видно, экспериментальные данные
группируются около прямой линии, точка
пересечения которой с осью ординат дает
значение
![]() =
0.8458 час-1,
а тангенс угла
наклона – значение
=
0.8458 час-1,
а тангенс угла
наклона – значение
![]() =
0.00485. Отсюда
нетрудно рассчитать значения параметров
KS
и rmax:
KS
= 9.75 мг/л,
rmax
= 0.85 час-1.
=
0.00485. Отсюда
нетрудно рассчитать значения параметров
KS
и rmax:
KS
= 9.75 мг/л,
rmax
= 0.85 час-1.
Задача 4.3
Условия: Исследовали кинетику роста популяции бактерий Pseudomonas fluorescens в хемостате при различных значениях протока D и концентрации лимитирующего субстрата, глюкозы, в подаваемой питательной среде S0. В каждом случае при достижении стационарного состояния хемостата измерялась концентрация биомассы клеток X и концентрация глюкозы S. Данные измерений представлены в таблице 4.3.
Таблица 4.3.
Концентрация
субстрата в подаваемой питательной
среде,
|
Проток, час-1 |
Стационар-ная концен-трация биомассы,
|
Стационар-ная концен-трация субстрата, , мМ |
|
|
|
4 |
0,1 |
1,903 |
0,025 |
3,97 |
10 |
40 |
4 |
0,2 |
1,822 |
0,056 |
3,94 |
5 |
17,85 |
4 |
0,4 |
1,812 |
0,168 |
3,83 |
2,5 |
5,95 |
4 |
0,6 |
1,67 |
0,49 |
3,51 |
1,67 |
2,04 |
4 |
0,75 |
1,041 |
1,858 |
2,14 |
1,33 |
0,54 |
|
|
|
|
|
|
|
8 |
0,1 |
1,921 |
0,025 |
7,97 |
10 |
40 |
8 |
0,2 |
1,892 |
0,056 |
7,94 |
5 |
17,86 |
8 |
0,7 |
1,67 |
1,04 |
6,96 |
1,43 |
0,96 |
8 |
0,75 |
1,417 |
1,875 |
6,12 |
1,33 |
0,53 |
8 |
0,8 |
0,27 |
7,198 |
0,80 |
1,25 |
0,14 |
|
|
|
|
|
|
|
16 |
0,2 |
1,882 |
0,057 |
15,94 |
5 |
17,54 |
16 |
0,4 |
1,89 |
0,169 |
15,83 |
2,5 |
5,92 |
16 |
0,7 |
1,728 |
1,038 |
14,96 |
1,43 |
0,96 |
16 |
0,75 |
1,685 |
1,883 |
14,12 |
1,33 |
0,53 |
16 |
0,8 |
1,093 |
7,095 |
8,90 |
1,25 |
0,14 |
Вопросы: Определить по представленным данным значения кинетических параметров, KS и rmax, и экономического коэффициента, характеризующих рост в хемостате популяции бактерий Pseudomonas fluorescens в указанных условиях.
Решение: Используем для решения поставленной задачи известные из теории хемостата соотношения (4.8), описывающие стационарное состояние культуры клеток (Перт, 1978):
![]() .
.
Для определения экономического коэффициента применим второе соотношение, предварительно произведя расчет вспомогательной переменной (таб.4.3, 5-ый столбец) и представив данные из таблицы (3 и 5 столбцы) в координатах { , }. Графики будем строить отдельно для каждого значения . На рис.4.5 представлены данные и расчет экономического коэффициента для = 4 мМ. Как видно, для данной концентрации Y1 = 0.45 г/мМ.

Рис.4.5. Определение экономического коэффициента Y1 по данным из таб.4.5 для = 4 мМ
Аналогично находим значения экономического коэффициента для двух других начальных концентраций лимитирующего субстрата (рис.4.6.): Y2 = 0.23 г/мМ и Y3 = 0.11 г/мМ.

Рис.4.6. Определение значений экономического коэффициента по данным из таб.4.5 для = 8 мМ (Y2) и = 16 мМ (Y3)
Для определения кинетических параметров роста клеток, KS и rmax, используем линейную анаморфозу первого из вышеуказанных соотношений (Перт, 1978):
![]() .
(4.10)
.
(4.10)
С
этой целью рассчитаем вспомогательные
переменные,
![]() и
и
![]() ,
как это указано в таб.4.4, столбцы 6-ой и
7-ой, и, далее, для каждого значения
построим график
в этих координатах. Экспериментальные
данные должны сгруппироваться около
прямой линии, которая отсекает на оси
абсцисс значение
,
как это указано в таб.4.4, столбцы 6-ой и
7-ой, и, далее, для каждого значения
построим график
в этих координатах. Экспериментальные
данные должны сгруппироваться около
прямой линии, которая отсекает на оси
абсцисс значение
![]() , а на оси ординат – значение
, а на оси ординат – значение
![]() .
На рис.4.8 (вверху) представлены данные
для
=
4 мМ. Произведенный
расчет дает следующие значения для
искомых кинетических параметров: rmax
= 0.82 час-1,
KS=
0.17 мМ.
.
На рис.4.8 (вверху) представлены данные
для
=
4 мМ. Произведенный
расчет дает следующие значения для
искомых кинетических параметров: rmax
= 0.82 час-1,
KS=
0.17 мМ.


Рис.4.8. Определение кинетических параметров роста клеток KS и rmax по данным хемостатного культивирования для = 4 мМ (вверху), а также для = 8 мМ и = 16 мМ (внизу) на основе данных из таб.4.3
Из нижнего графика на рис.4.8 следует, что значения KS и rmax для для = 8 мМ и = 16 мМ идентичны и равны таковым для = 4 мМ.
Таким образом, из полученных результатов следует, что при хемостатном культивировании исследуемых бактерий на глюкозе как лимитирующем субстрате увеличение ее начальной концентрации с 4 мМ до 16 мМ существенно снижает экономический коэффициент с 0.45 до 0.11 г/мМ и не влияет на значения кинетических параметров роста KS и rmax, которые соответственно равны rmax = 0.82 час-1, KS = 0.17 мМ. По-видимому, это свидетельствует о переключении типа энергетического метаболизма с полностью затратного на запасающий и неизменности транспортных систем для глюкозы.
Задача 4.4
Условия: Исследовали динамику популяции кишечной палочки E.coli при культивировании в хемостате на бедной минеральной среде при лимитировании глюкозой, начальная концентрация которой в подаваемой среде (S0) составляла 0.666 г/л. Изучали поведение популяции во время переходного процесса, обусловленного изменением протока через ферментер с величины Dисх = 0.25 час-1 на величину D = 0.765 час-1. В ходе эксперимента определяли концентрацию биомассы клеток и концентрацию глюкозы. За нулевую отметку времени был взят момент смены протока. Данные представлены в таблице 4.4 и на рис.4.9 (Евдокимов и др., 2000).
Таблица 4.4
Время, t, час
|
Концентрация биомассы, X, г/л |
Концентрация субстрата, S, г/л |
|
|
|
0 |
0,166 |
0,198 |
0,468 |
|
|
1 |
0,142 |
0,297 |
0,369 |
-0,15644 |
-0,18693 |
2 |
0,116 |
0,326 |
0,34 |
-0,17799 |
-0,21741 |
3 |
0,099 |
0,396 |
0,27 |
-0,17175 |
-0,21418 |
4 |
0,095 |
0,446 |
0,220 |
-0,13984 |
-0,1755 |
5 |
0,078 |
0,446 |
0,220 |
-0,15167 |
-0,1966 |
10 |
0,064 |
0,506 |
0,16 |
-0,09531 |
-0,12852 |
14 |
0,058 |
0,509 |
0,157 |
-0,07511 |
-0,10373 |
24 |
0,049 |
0,531 |
0,135 |
-0,05084 |
-0,07378 |
30 |
0,039 |
0,586 |
0,08 |
-0,04828 |
-0,07699 |
48 |
0,025 |
0,594 |
0,072 |
|
|

Рис.4.9. Изменение концентрации биомассы клеток кишечной палочки при культивировании в хемостате при изменении протока: 0.25 час-1 0.765 час-1.
Стрелкой указан момент изменения протока.
Вопросы: Определить по представленным данным значения кинетических параметров, KS и rmax, и экономического коэффициента, характеризующих рост в хемостате популяции бактерий E.coli в указанных условиях переходного процесса.
Решение:
На
первом этапе определим среднее значение
экономического коэффициента
![]() в ходе процесса, исходя из предположения,
что он изменяется несущественно при
смене протока. Для этого воспользуемся
подходом, рассмотренным в предыдущей
задаче. Вспомогательная переменная
представлена в
4-ом столбце таб.4.4, а решение – на
рис.4.10.
в ходе процесса, исходя из предположения,
что он изменяется несущественно при
смене протока. Для этого воспользуемся
подходом, рассмотренным в предыдущей
задаче. Вспомогательная переменная
представлена в
4-ом столбце таб.4.4, а решение – на
рис.4.10.
Как видно,
экспериментальные данные достаточно
компактно группируются около прямой
линии. Это свидетельствует о том, что в
течение переходного процесса изменения
концентрации биомассы и субстрата
связаны таким же соотношением, как и в
стационарном состоянии:![]() ,
и среднее значение экономического
коэффициента
=
0.35 достаточно стабильно.
,
и среднее значение экономического
коэффициента
=
0.35 достаточно стабильно.

Рис.4.10. Определение среднего значения экономического коэффициента по данным из таб.4.4
Для определения кинетических параметров, KS и rmax, используем следующую теорию, развитую в работе (Евдокимов, Шаповалов, 2000). Согласно это теории, если хемостатная культура клеток находится в стационарном состоянии при протоке Dисх, и далее проток изменяется Dисх D, то начиная с момента смены протока динамика системы описывается следующим уравнением:
 ,
(4.11)
,
(4.11)
где
![]() и
и
![]() -
стационарная концентрация биомассы
при протоке D.
Уравнение (4.11) будем использовать в
виде следующей линейной анаморфозы:
-
стационарная концентрация биомассы
при протоке D.
Уравнение (4.11) будем использовать в
виде следующей линейной анаморфозы:
 .
(4.12)
.
(4.12)
При
расчете вспомогательных переменных,
и
![]() ,
(таб.4.4, столбцы 5-ый и 6-ой), применялись
следующие значения начальной и
стационарной концентраций биомассы
клеток:
,
(таб.4.4, столбцы 5-ый и 6-ой), применялись
следующие значения начальной и
стационарной концентраций биомассы
клеток:
![]() =0.166
мг/л и
=0.025
мг/л. Экспериментальные данные,
представленные в указанных координатах,
как видно из рис.4.11, группируются около
прямой линии, которая согласно выражению
(4.6), отсекает на оси ординат значение
=0.166
мг/л и
=0.025
мг/л. Экспериментальные данные,
представленные в указанных координатах,
как видно из рис.4.11, группируются около
прямой линии, которая согласно выражению
(4.6), отсекает на оси ординат значение
 ,
а тангенс угла ее наклона дает значение
,
а тангенс угла ее наклона дает значение
 .
.

Рис.4.11. Определение кинетических параметров роста клеток KS и rmax по данным хемостатного культивирования на основе данных из таб.4.4
Далее, решая найденные соотношения как систему уравнений с двумя неизвестными, легко найти искомые значения кинетических параметров: rmax = 0.8 час-1, KS = 13.6 мг/л.
Задача 4.5
Условия: Исследовали кинетику потребления энергетического субстрата, глюкозы, при росте популяции кишечной палочки E.coli в хемостате в стационарных условиях. Опыты проводили при различных начальных концентрациях глюкозы и значениях протока. В ходе эксперимента измеряли стационарные концентрации биомассы и глюкозы. Данные представлены в таблице 4.5.
Таблица 4.5
Начальная концентрация глюкозы, S0, мМ |
Проток, D, час-1
|
Стационарная концентрация биомассы,
|
Стационарная концентрация глюкозы,
|
Y
|
|
|
4 |
0,1 |
1,74 |
0,03 |
0,438 |
10 |
2,28 |
4 |
0,2 |
1,93 |
0,06 |
0,49 |
5 |
2,04 |
4 |
0,4 |
2 |
0,18 |
0,524 |
2,5 |
1,91 |
4 |
0,6 |
1,88 |
0,49 |
0,536 |
1,67 |
1,87 |
4 |
0,75 |
1,13 |
1,91 |
0,541 |
1,33 |
1,85 |
|
|
|
|
|
|
|
8 |
0,1 |
1,96 |
0,03 |
0,246 |
10 |
4,07 |
8 |
0,2 |
2,08 |
0,06 |
0,262 |
5 |
3,82 |
8 |
0,7 |
1,92 |
1,02 |
0,275 |
1,43 |
3,64 |
8 |
0,75 |
1,68 |
1,88 |
0,275 |
1,33 |
3,64 |
8 |
0,8 |
0,28 |
6,98 |
0,274 |
1,25 |
3,64 |
|
|
|
|
|
|
|
16 |
0,2 |
2,16 |
0,06 |
0,136 |
5 |
7,38 |
16 |
0,4 |
2,18 |
0,17 |
0,138 |
2,5 |
7,26 |
16 |
0,7 |
2,08 |
1,01 |
0,1388 |
1,43 |
7,21 |
16 |
0,75 |
1,96 |
1,86 |
0,1386 |
1,33 |
7,21 |
16 |
0,8 |
1,26 |
6,94 |
0,1390 |
1,25 |
7,19 |
Вопросы: Определить по представленным данным значения ростового экономического коэффициента Yрост и трат на поддержание m, характеризующих кинетику потребления энергетического субстрата, глюкозы, при росте популяции бактерий E.coli в хемостате в указанных условиях.
Решение:
На
первом этапе решения определим для
каждого варианта условий значение
суммарного экономического коэффициента
Y.
Для расчетов Y
(5-ый столбец таб.4.5.) будем использовать
соотношение (4.8) в виде
![]() и
данные из 1-го, 3-го и 4-го столбцов таб.4.5.
и
данные из 1-го, 3-го и 4-го столбцов таб.4.5.
В соответствии с теорией суммарный экономический коэффициент Y, ростовой экономический коэффициент Yрост и траты на поддержание m связаны соотношением (4.2):
.
Учитывая, что в стационарном состоянии в хемостате удельная скорость роста популяции r равна протоку D , будем использовать
соотношение (4.2) в виде:
![]() .
(4.13)
.
(4.13)


Рис.4.12 - 4.13. Определение ростового экономического коэффициента Yрост и трат на поддержание m по данным из таб.4.5 для S0 = 4 мМ (вверху) и S0 = 8мМ (внизу)
С
этой целью рассчитаем вспомогательные
переменные
и
![]() (столбцы 6-ой и 7-ой таб.4.5) и построим
графики в этих координатах для каждого
значения начальной концентрации глюкозы
S0
(рис.4.12-4.14).
(столбцы 6-ой и 7-ой таб.4.5) и построим
графики в этих координатах для каждого
значения начальной концентрации глюкозы
S0
(рис.4.12-4.14).
Экспериментальные данные на этих графиках группируются около прямых линий, которые согласно выражению (4.13) отсекают на оси ординат значение 1/Yрост, а тангенсы угла наклона дают значения m.

Рис.4.14 . Определение ростового экономического коэффициента Yрост и трат на поддержание m по данным из таб.4.5 для S0 = 16 мМ
Полученные в результате значения ростового экономического коэффициента и трат на поддержание представлены в таб.4.6.
Таблица 4.6
Начальная концентрация глюкозы, S0, мМ |
Ростовой экономический коэффициент, Yрост , г/мМ |
Траты на поддержание, m, час-1 |
4 |
0,56 |
0,05 |
8 |
0,28 |
0,049 |
16 |
0,14 |
0,048 |