

Излучения
1. Видимый свет
Электромагнитные волны, имеющие длину 400—740 нм, соответствуют видимому свету. Видимый свет в качестве источника энергии используют фототрофные бактерии. Цианобактерии осуществляют оксигенный фотосинтез, в процессе которого выделяется молекулярный кислород. Адаптация к разнообразным физико-химическим условиям среды, способность к азотфиксации, термофильность или галофильность некоторых форм определяют весьма широкое распространие цианобактерий на Земле. Oscillatoria Umnetica иногда способна и к аноксигенному фотосинтезу с использованием в качестве донора электронов H2S и устойчива к очень высоким Концентрациям сероводорода.
Пурпурные и зеленые бактерии осуществляют аноксигенныи фотосинтез. Он приурочен к анаэробным условиям, и большинство этих бактерий строгие анаэробы, многие нуждаются в восстановленных соединениях азота и серы. Некоторые формы способны к аэробному дыханию, но в присутствии молекулярного кислорода не могут осуществлять фотосинтез, так как уже в--низких концентрациях молекулярный кислород репрессирует синтез бактериохлорофиллов.
Развитие бактерий, осуществляющих аноксигенныи фотосинтез происходит главным образом в сильно загрязненных водоемах и гиполимнионе озер. Здесь содержатся соединения, специфически необходимые для аноксигенных фототрофов,—сероводород, молекулярный водород, простые органические вещества, образующиеся в результате брожений, которые осуществляются анаэробными бактериями, обитающими в осадках.
Пурпурные и зеленые бактерии часто развиваются в водоемах на глубине под слоем водорослей и цианобактерий, в котором происходит рост высших растений. Перечисленные фото-трофы поглощают значительную часть света в областях спектра, соответствующих адсорбционной способности их светособи-рающих антенн. Светособирающие антенны- аноксигедных фототрофов имеют другие спектральные характеристики, это позволяет им осуществлять фотосинтез за счет света, профильтрованного через клетки эукариот и цианобактерий (табл. 1).

Бактериохлорофиллы имеют максимумы поглощения в красной и инфракрасной частях спектра. Этот свет не используется для оксигенного фотосинтеза. Бактериохлорофилл б поглощает световые волны, длина которых близка к максимальной, еще допекающей их использование в процессе фотосинтеза. Кванты более длинноволнового света несут слишком мало энергии и я могут участвовать в фотохимических реакциях. Существенная особенностью бактериохлорофиллов является также то, что их включение в мембранный светособирающий комплекс приводит к смещению максимумов поглощения в красную область спектра в гораздо большей степени, чем это характерно хлорофилла а оксигенных фототрофов.
Некоторые виды пурпурных бактерий живут на мелководье под слоем влажного песка толщиной в несколько сантиметров который играет роль красного фильтра. Однако на более значительную глубину в водоем красный свет не проникает. Представители зеленых серобактерий имеют каротиноиды, улавливающие световые волны длиной около 460 нм, и способны развиваться в водоемах на глубинах, более 20 м, куда проникает свет только этой длины волны.
В конце 70-х годов были обнаружены аэробные гетеротрофные бактерии, содержащие бактериохлорофилл а. Они отнесены к нескольким родам. Представители рода Erythrobacter—мелкие грамотрицательные палочки с субполярным жгутиком. Кроме бактериохлорофилла они синтезируют каротиноидные пигменты. Ряд штаммов Erythrobacter выделен с поверхности морских макрофитов. Для Erythrobacter свет не может быть единственным источником энергии ни в аэробных, ни в -анаэробных условиях, однако используется этой бактерией в качестве дополнительного источника энергии. При освещении усиливается синтез АТФ, включение СО2 в органическое вещество клетки, наблюдается фотоингибирование дыхания, что свидетельствует о существовании у этих бактерий регуляторных механизмов энергетического обмена, позволяющих при освещении более экономно расходовать органический энергетический субстрат. При исчерпывании органического субстрата в условиях освещения жизнеспособность клеток сохраняется значительно дольше, чем в темноте, В противоположность тому, что наблюдается у пурпурных и зеленых бактерий, синтез бактериохлорофилла а у Erythrobacter стимулируется молекулярным кислородом.
Бактериохлорофилл а содержится и в мембранах гетерот рофной аэробной бактерии Protaminobacter ruber, также синтезирующей АТФ за счет энергии света. Подобная способности обааружена и у некоторых штаммов бактерий из рода Pseiidomonas.
Энергия видимого света может быть использована экстремально галофильными архебактериями, которые, однако, не являются настоящими фототрофами, поскольку не могут расти, используя свет как единственный источник энергии. Экстремально галофильная архебактерия рода Halobacterium — аэробный ге-теротроф, основное количество АТФ у которой образуется в результате процесса аэробного дыхания. Halobacterium обитает поверхностных слоях соленых озер, заливов, лиманов, где растворимость кислорода значительно ниже, чем в пресной или Юрской воде. При истощении в среде кислорода происходит репрессия генов, ответственных за синтез хромопротеина бактеоиородопсина, который включается в состав клеточной мембраны, образуя участки, обозначаемые как пурпурные мембраны. Под действием света происходит выброс протонов во внешнюю среду, создающийся при этом трансмембранный градиент концентрации протонов используется для синтеза АТФ. В тех же мембранах содержится второй пигмент—галородопсин, который при поглощении фотонов осуществляет транспорт ионов хлора в клетку, что также приводит к увеличению мембранного потенциала. Это позволяет бактериям поддерживать жизнеспособность в анаэробных условиях. Экстремально галофильные архебактерии обитают в соленых озерах, расположенных в районах с постоянной высокой инсоляцией.
Видимый свет влияет на поведение фототрофных бактерий. У (Ьототрофных эубактерий и архебактерии Halobacterium salina-rium наблюдается явление фототаксиса: бактерии способны реагировать на изменение спектрального состава света или освещенности. У эубактерий фоторецепторами служат бактерио-хлорофиллы и каротиноиды, т. е. те же пигменты, которые поглощают свет в процессе фотосинтеза. У архебактерии обнаружены специальные сенсорные пигменты. Бактериальные клетки, способные к фототаксису, резко изменяют направление движения, если попадают из более освещенного участка в менее освещенный или в совсем неосвещенный. В результате бактерии двигаются к свету—это положительный фототаксис. В некоторых условиях наблюдается отрицательный фототаксис—движение в сторону уменьшения освещенности.- Подобные реакции иногда определяют как фотофобные. При резком снижении освещенности также происходит изменение направления движения клеток, т. е. они реагируют da изменение освещенности не в пространстве, а во времени, как и в случае реакции на изменение концентрации хемоэффекторов при хемотаксисе (см. с. 126). Изменение силы света может привести к изменению скорости движения бактерий, это явление определяют как фотокинез. В реакциях фототаксиса эубактерий участвуют реакционные центры фотосинтеза. Мутанты с нарушенными реакционными центрами теряют способность к фототаксису. В этом отношении фототрофные бактерии отличаются от подвижных фототрофных эукариот, фототаксис которых не связан с фотосинтезом.
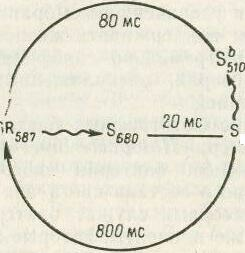
Клетки архебактерии Halobacterium salinarium проявляют фототаксис, причем красный свет действует как аттрактант, а синий—как репеллент. Оба эффекта определяются адсорбцией света одним и тем же пигментом — сенсорным родопсином (S на рис. 1). В отсутствие света сенсорный родопсин имеет максимум поглощения при 587 нм; поглощая фотон красного света, молекула переходит в состояние, при котором максимум смещается в область 680 нм и затем 373 нм. При этом генерируется: сигнал, приводящий к снижению частоты кувырканий клетки,. т. е. к аттракции. Форма 8373 может вернуться в исходное состояние за счет чисто термальной релаксации, и так фотоцикл заканчивается. Но если молекула 8373 успеет поглотить фотов синего света, она переходит в промежуточное состояние S510 а из него—в исходное. При этом генерируется сигнал, приводящий к увеличению частоты кувырканий бактерий, т. е. репеллентному эффекту. Очевидно, что в отсутствие красного света синий не вызывает этого эффекта. При освещении эффекты суммируются, и конечный результат будет зависеть от относительной интенсивности света той или иной длины волны.
Таким образом, энергия света может быть использована бактериями. Пути ее использования / прокариот значительно разнообразнее, чем у эукариот, где известен только один тип оксигенного фотосинтеза. Для некоторых бактерий, не способных использовать энергию света, он служит в качестве регулятора определенных процессов обмена. Так, у водной бактерия Pseudomonas putida наблюдали активацию светом некоторых ферментов метаболизма, что можно рассматривать как адаптацию, поскольку именно при освещении начинается фотосинтез фитопланктона, продукты которого используются гетеротрофными водными бактериями. Под фотохромностыо понимают зависимость образования-пигментов некоторыми микроорганизмами от освещения. Фотохромность свойственна миксобактерии Myxococcus xanthus особенно характерна для многих актиномицетов и близких к ним организмов. В частности, синтез каротиноидов некоторыми микобактериями стимулируется синим светом, причем процессы;
синтеза пигментов имеют светозависимые и темновые этапы.! Фотохромность может контролироваться как хромосомными, j так и плазмидными генами. Кроме того,, пигменты способны; защищать микроорганизмы от действия видимого света.
Солнечный свет может оказывать сильный антимикробный эффект. Так, более 99,9% клеток штамма Escherichia coli с нарушенными репарационными механизмами погибают после облучения солнечным светом в течение трех минут. Правда, при этом более 80% летальных повреждений связано с действием; света длиной волны менее 312 нм, действие видимого света ответственно менее чем за 1 % летальных повреждений. Видимый свет длиной волны 450 нм индуцирует замены пар оснований д мутации сдвига рамки у Е. coli. Световые волны длиной 550 им и особенно 410 нм вызывают фотолизис клеток Myxococcus xanthus. Эффект определяется поглощением света желе-зопорфиринами.
Существуют вещества фотосенсибилизаторы, в- молекуле ко-тооых имеется хромофор, поглощающий свет и передающий его энергию другим молекулам, не способным поглощать свет. Через бесцветные клетки свет проходит без последствий, но если в такую клетку введен фотосенсибилизатор, она повреждается. Известно более 400 веществ, обладающих свойствами фотосенсибилизаторов. Их хромофорами обычно оказываются циклические ядра. Фотосенсибилизаторы содержатся в промышленных стоках, смоге, многие лекарства представляют собой фотосенсибилизаторы, в том числе сульфаниламиды, ряд антибиотиков, хинин, рибофлавин, метиленовый синий и другие красители. Среди .природных веществ фотосенсибилизаторами являются псоралены (фурокумарины), хлорофилл, фикобилины, порфири-ны и др. Многие растения содержат псоралены, свет в их присутствии повреждает ДНК. Порфирины находятся во всех аэробных и некоторых анаэробных клетках. При нарушениях в путях метаболизма могут накапливаться промежуточные продукты синтеза порфиринов, также действующие как фотосенсибилизаторы.
Некоторые фотосенсибилизаторы действуют только в присутствии кислорода—это так называемый фотодинамический эффект (см. с. 92).